Selection of Wine Saccharomyces cerevisiae Strains and Their Screening for the Adsorption Activity of Pigments, Phenolics and Ochratoxin A

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Yeast Isolation
2.2. Preliminary Screening
2.3. Fermentation Trials and Adsorption Activity of Pigments and Phenolics
2.4. OTA Adsorption Activity
2.5. Statistical Analysis
3. Results
3.1. Preliminary Trials on the S. cerevisiae Strains
3.2. Assessment of the Fermentation Vigour and Adsorption Activity
3.3. Assessment of the OTA Adsorption Activity and Final Strain Selection
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mateo, R.; Medina, Á.; Mateo, E.M.; Mateo, F.; Jiménez, M. An overview of ochratoxin A in beer and wine. Int. J. Food Microbiol. 2007, 119, 79–83. [Google Scholar] [CrossRef]
- Piotrowska, M.; Nowak, A.; Czyzowska, A. Removal of ochratoxin A by wine Saccharomyces cerevisiae strains. Eur. Food Res. Technol. 2013, 236, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Cecchini, F.; Morassut, M.; Saiz, J.C.; Garcia-Moruno, E. Anthocyanins enhance yeast’s adsorption of Ochratoxin A during the alcoholic fermentation. Eur. Food Res. Technol. 2019, 245, 309–314. [Google Scholar] [CrossRef]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar] [CrossRef]
- Dachery, B.; Manfroi, V.; Berleze, K.J.; Welke, J.E. Occurrence of ochratoxin A in grapes, juices and wines and risk assessment related to this mycotoxin exposure. Ciência Rural 2015, 46, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Rigó, K.; Téren, J.; Mesterhàzy, A. Recent advances in ochratoxin research I. Production, detection and occurrence of ochratoxins. Cereal Res. Commun. 2001, 29, 85–92. [Google Scholar] [CrossRef]
- Angioni, A.; Caboni, P.; Garau, A.; Farris, A.; Orro, D.; Budroni, M.; Cabras, P. In vitro interaction between ochratoxin A and different strains of Saccharomyces cerevisiae and Kloeckera apiculata. J. Agric. Food Chem. 2007, 55, 2043–2048. [Google Scholar] [CrossRef]
- Delage, N.; d’Harlingue, A.; Colonna Ceccaldi, B.; Bompeix, G. Occurrence of mycotoxins in fruit juices and wine. Food Control 2003, 14, 225–227. [Google Scholar] [CrossRef]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Magan, N.; Logrieco, A. European research on ochratoxin A in grapes and wine. Int. J. Food Microbiol. 2006, 111, S2. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.; Patiño, B. Wine Contamination with Ochratoxins: A Review. Beverages 2018, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Otteneder, H.; Majerus, P. Occurrence of ochratoxin A (OTA) in wines: Influence of the type of wine and its geographical origin. Food Addit. Contam. 2000, 17, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Paterson, R.R.M.; Venâncio, A. Biodegradation of ochratoxin a for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caridi, A.; Galvano, F.; Tafuri, A.; Ritieni, A. Ochratoxin A removal during winemaking. Enzyme Microb. Technol. 2006, 40, 122–126. [Google Scholar] [CrossRef]
- Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Beneduce, L.; Bevilacqua, A. Ochratoxin A removal by Saccharomyces cerevisiae strains: Effect of wine-related physicochemical factors. J. Sci. Food Agric. 2013, 93, 2110–2115. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G. Selection of an autochthonous Saccharomyces cerevisiae strain for the vinification of “Moscato di Saracena”, a southern Italy (Calabria Region) passito wine. Food Microbiol. 2016, 54, 30–39. [Google Scholar] [CrossRef]
- Petruzzi, L.; Bevilacqua, A.; Baiano, A.; Beneduce, L.; Corbo, M.R.; Sinigaglia, M. Study of Saccharomyces cerevisiae W13 as a functional starter for the removal of ochratoxin A. Food Control 2014, 35, 373–377. [Google Scholar] [CrossRef]
- Petruzzi, L.; Bevilacqua, A.; Corbo, M.R.; Garofalo, C.; Baiano, A.; Sinigaglia, M. Selection of autochthonous Saccharomyces cerevisiae strains as wine starters using a polyphasic approach and ochratoxin a removal. J. Food Prot. 2014, 77, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Baiano, A.; De Gianni, A.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Differential adsorption of ochratoxin a and anthocyanins by inactivated yeasts and yeast cell walls during simulation of wine aging. Toxins 2015, 7, 4350–4365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caridi, A. Improved screening method for the selection of wine yeasts based on their pigment adsorption activity. Food Technol. Biotechnol. 2013, 51, 137–144. [Google Scholar]
- De Vero, L.; Boniotti, M.B.; Budroni, M.; Buzzini, P.; Cassanelli, S.; Comunian, R.; Gullo, M.; Logrieco, A.F.; Mannazzu, I.; Musumeci, R.; et al. Preservation, characterization and exploitation of microbial biodiversity: The perspective of the italian network of culture collections. Microorganisms 2019, 7, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulvirenti, A.; Rainieri, S.; Boveri, S.; Giudici, P. Optimizing the selection process of yeast starter cultures by preselecting strains dominating spontaneous fermentations. Can. J. Microbiol. 2009, 55, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Lemaresquier, H.; Gainvors, A.; Lequart, C.; Charlemagne, B.; Frezier, V.; Belarbi, A. Sélection de levures œnologiques à activité clarifiante: Les différentes techniques utilisées pour caractériser des levures, intérêt de la sélection d’ une levure productrice d’ enzymes pectolytiques. Rev. française d’oenologie 1995, 35, 23–29. [Google Scholar]
- Nickersok, W.J. Reduction of inorganic substances by yeasts. J. Infect. Dis. 1953, 93, 43–56. [Google Scholar] [CrossRef]
- Glories, Y. La couleur des vins rouges. lre partie: Les équilibres des anthocyanes et des tanins. OENO One 1984, 18, 195. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Caridi, A.; Cufari, A.; Lovino, R.; Palumbo, R.; Tedesco, I. Influence of Yeast on Polyphenol Composition of Wine. Food Technol. Biotechnol. 2004, 42, 37–40. [Google Scholar]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of Montepulciano d’abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Perpetuini, G.; Tittarelli, F.; Battistelli, N.; Suzzi, G.; Tofalo, R. Contribution of Pichia manshurica strains to aroma profile of organic wines. Eur. Food Res. Technol. 2020, 246, 1405–1417. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Colomo, B.; Suárez, J.A. Cell wall anthocyanin adsorption by different Saccharomyces strains during the fermentation of Vitis vinifera L. cv Graciano grapes. Eur. Food Res. Technol. 2005, 220, 341–346. [Google Scholar] [CrossRef]
- Stockley, C.S.; Høj, P.B. Better wine for better health: Fact or fiction? Aust. J. Grape Wine Res. 2005, 11, 127–138. [Google Scholar] [CrossRef]
- Caridi, A.; Sidari, R.; Giuffrè, A.M.; Pellicanò, T.M.; Sicari, V.; Zappia, C.; Poiana, M. Test of four generations of Saccharomyces cerevisiae concerning their effect on antioxidant phenolic compounds in wine. Eur. Food Res. Technol. 2017, 243, 1287–1294. [Google Scholar] [CrossRef]
- Mazauric, J.P.; Salmon, J.M. Interactions between yeast lees and wine polyphenols during simulation of wine aging: I. Analysis of remnant polyphenolic compounds in the resulting wines. J. Agric. Food Chem. 2005, 53, 5647–5653. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Yeast interactions with anthocyanins during red wine fermentation. Am. J. Enol. Vitic. 2005, 56, 104–109. [Google Scholar]
- Petruzzi, L.; Bevilacqua, A.; Corbo, M.R.; Speranza, B.; Capozzi, V.; Sinigaglia, M. A Focus on quality and safety traits of Saccharomyces cerevisiae isolated from Uva di Troia Grape Variety. J. Food Sci. 2017, 82, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Mozaffary, P.; Milani, J.M.; Heshmati, A. The influence of yeast level and fermentation temperature on Ochratoxin A decrement during bread making. Food Sci. Nutr. 2019, 7, 2144–2150. [Google Scholar] [CrossRef] [PubMed]
- Bonciani, T.; De Vero, L.; Mezzetti, F.; Fay, J.C.; Giudici, P. A multi-phase approach to select new wine yeast strains with enhanced fermentative fitness and glutathione production. Appl. Microbiol. Biotechnol. 2018, 102, 2269–2278. [Google Scholar] [CrossRef]
- Bonciani, T.; De Vero, L.; Giannuzzi, E.; Verspohl, A.; Giudici, P. Qualitative and quantitative screening of the β-glucosidase activity in Saccharomyces cerevisiae and Saccharomyces uvarum strains isolated from refrigerated must. Lett. Appl. Microbiol. 2018, 67, 72–78. [Google Scholar] [CrossRef]
- Giudici, P.; Solieri, L.; Pulvirenti, A.M.; Cassanelli, S. Strategies and perspectives for genetic improvement of wine yeasts. Appl. Microbiol. Biotechnol. 2005, 66, 622–628. [Google Scholar] [CrossRef] [PubMed]
- De Vero, L.; Bonciani, T.; Verspohl, A.; Mezzetti, F.; Giudici, P. High-glutathione producing yeasts obtained by genetic improvement strategies: A focus on adaptive evolution approaches for novel wine strains. AIMS Microbiol. 2017, 3, 155–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzetti, F.; Fay, J.C.; Giudici, P.; De Vero, L. Genetic variation and expression changes associated with molybdate resistance from a glutathione producing wine strain of Saccharomyces cerevisiae. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caridi, A.; Sidari, R.; Pulvirenti, A.; Meca, G.; Ritieni, A. Ochratoxin A adsorption phenotype: An inheritable yeast trait. J. Gen. Appl. Microbiol. 2012, 58, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caridi, A.; Sidari, R.; Solieri, L.; Cufari, A.; Giudici, P. Wine colour adsorption phenotype: An inheritable quantitative trait loci of yeasts. J. Appl. Microbiol. 2007, 103, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Tempère, S.; Marchal, A.; Barbe, J.C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Wine Sample Cultivar. | Region (Country) | Number of Non-Redundant S. cerevisiae Strains (Codes Range) |
|---|---|---|
| Nerello mascalese | Sicily (Italy) | 21 (RE001–RE021) |
| Carricante | 3 (RE022–RE024) | |
| Grecanico | 8 (RE025–RE032) | |
| Grecanico | Penedès and La Rioja (Spain) | 2 (RE033–RE034) |
| Tempranillo | 33 (RE035–RE067) | |
| Touriga | 3 (RE068–RE070) | |
| Touriga national | Porto (Portugal) | 7 (RE071–RE077) |
| Touriga franca | 12 (RE078–RE089) | |
| Tinta rorizi | 10 (RE090–RE099) |
| Strain | Yeast Biomass Colour on Grape-Skin Agar | Fermentation Vigour after 3 Days (g CO2/100 mL) | Wine Colour Intensity | Phenolics Content | ||
|---|---|---|---|---|---|---|
| Red Component | Green Component | Blue Component | Absorbance (420 + 520 + 620) nm | Folin–Ciocalteu’s Index (FC) | ||
| RE001 | 81 ± 4 | 40 ± 2 | 45 ± 2 | 8.58 ± 0.11 | 3.908 ± 0.181 | 11.08 ± 5.68 |
| RE002 | 73 ± 3 | 37 ± 1 | 40 ± 2 | 8.45 ± 0.14 | 4.933 ± 0.392 | 11.98 ± 6.48 |
| RE004 | 95 ± 2 | 52 ± 2 | 50 ± 1 | 8.65 ± 0.22 | 4.580 ± 1.052 | 10.98 ± 5.87 |
| RE005 | 78 ± 3 | 39 ± 2 | 39 ± 2 | 9.14 ± 0.08 | 3.145 ± 0.818 | 8.83 ± 3.30 |
| RE006 | 73 ± 3 | 37 ± 4 | 40 ± 4 | 8.56 ± 0.07 | 3.897 ± 0.161 | 11.33 ± 5.68 |
| RE007 | 60 ± 5 | 28 ± 2 | 34 ± 2 | 8.58 ± 0.11 | 4.108 ± 0.590 | 10.22 ± 2.55 |
| RE008 | 82 ± 3 | 38 ± 2 | 42 ± 2 | 8.12 ± 0.14 | 1.855 ± 0.398 | 9.18 ± 2.55 |
| RE010 | 71 ± 2 | 36 ± 2 | 39 ± 2 | 8.14 ± 0.22 | 3.528 ± 0.763 | 12.08 ± 7.07 |
| RE011 | 65 ± 1 | 32 ± 2 | 38 ± 2 | 9.15 ± 0.08 | 5.418 ± 0.460 | 22.85 ± 3.50 |
| RE012 | 66 ± 2 | 31 ± 2 | 37 ± 1 | 8.41 ± 0.07 | 5.557 ± 0.333 | 10.25 ± 8.46 |
| RE013 | 67 ± 4 | 33 ± 1 | 41 ± 1 | 8.25 ± 0.11 | 2.833 ± 0.855 | 9.12 ± 5.74 |
| RE014 | 80 ± 2 | 43 ± 1 | 46 ± 2 | 7.45 ± 0.14 | 4.715 ± 0.654 | 12.82 ± 2.33 |
| RE015 | 86 ± 2 | 48 ± 2 | 49 ± 3 | 7.12 ± 0.18 | 3.532 ± 0.488 | 12.25 ± 0.73 |
| RE017 | 108 ± 5 | 65 ± 4 | 61 ± 4 | 8.51 ± 0.08 | 6.417 ± 0.336 | 21.55 ± 0.41 |
| RE018 | 91 ± 3 | 49 ± 3 | 50 ± 1 | 9.56 ± 0.07 | 1.588 ± 0.172 | 9.65 ± 3.34 |
| RE019 | 69 ± 3 | 34 ± 1 | 39 ± 2 | 7.75 ± 0.11 | 4.498 ± 0.595 | 11.00 ± 2.35 |
| RE020 | 69 ± 2 | 36 ± 3 | 41 ± 3 | 9.12 ± 0.14 | 4.870 ± 0.441 | 12.85 ± 6.29 |
| RE022 | 72 ± 4 | 39 ± 2 | 42 ± 2 | 8.90 ± 0.22 | 4.940 ± 0.440 | 18.42 ± 2.68 |
| RE023 | 84 ± 2 | 45 ± 3 | 45 ± 2 | 7.52 ± 0.08 | 3.128 ± 0.418 | 8.38 ± 6.13 |
| RE024 | 70 ± 5 | 35 ± 1 | 38 ± 2 | 7.75 ± 0.07 | 3.147 ± 0.511 | 12.97 ± 1.88 |
| RE025 | 70 ± 4 | 34 ± 2 | 39 ± 4 | 8.42 ± 0.11 | 3.317 ± 0.587 | 8.62 ± 5.15 |
| RE026 | 68 ± 4 | 34 ± 2 | 38 ± 3 | 8.21 ± 0.14 | 2.358 ± 0.660 | 9.62 ± 5.40 |
| RE027 | 114 ± 3 | 69 ± 2 | 62 ± 3 | 7.21 ± 0.18 | 3.957 ± 0.305 | 12.07 ± 1.83 |
| RE028 | 62 ± 1 | 30 ± 2 | 36 ± 1 | 8.05 ± 0.22 | 4.958 ± 0.398 | 12.77 ± 2.48 |
| RE029 | 57 ± 3 | 33 ± 2 | 37 ± 3 | 8.74 ± 0.08 | 1.530 ± 0.411 | 9.43 ± 5.34 |
| RE030 | 70 ± 2 | 36 ± 2 | 40 ± 3 | 7.78 ± 0.07 | 3.483 ± 0.700 | 10.17 ± 3.60 |
| RE031 | 79 ± 2 | 38 ± 1 | 40 ± 2 | 7.89 ± 0.11 | 2.888 ± 0.935 | 12.17 ± 3.12 |
| RE032 | 91 ± 3 | 50 ± 2 | 51 ± 2 | 7.56 ± 0.14 | 3.522 ± 0.713 | 15.08 ± 0.53 |
| RE033 | 76 ± 4 | 36 ± 2 | 40 ± 3 | 9.78 ± 0.18 | 4.990 ± 0.156 | 15.45 ± 2.99 |
| RE034 | 61 ± 3 | 28 ± 4 | 33 ± 2 | 8.95 ± 0.22 | 3.903 ± 0.582 | 9.72 ± 3.77 |
| RE036 | 83 ± 3 | 44 ± 1 | 47 ± 3 | 8.45 ± 0.07 | 4.817 ± 0.189 | 16.43 ± 1.43 |
| RE037 | 76 ± 2 | 40 ± 2 | 40 ± 2 | 9.67 ± 0.11 | 2.580 ± 0.420 | 7.55 ± 6.37 |
| RE038 | 76 ± 3 | 40 ± 4 | 41 ± 3 | 9.56 ± 0.14 | 4.462 ± 1.220 | 11.37 ± 2.02 |
| RE039 | 74 ± 1 | 37 ± 2 | 42 ± 4 | 9.63 ± 0.18 | 4.235 ± 0.075 | 13.78 ± 0.88 |
| RE040 | 62 ± 2 | 31 ± 3 | 35 ± 3 | 8.67 ± 0.22 | 4.922 ± 0.473 | 12.32 ± 3.07 |
| RE041 | 85 ± 3 | 45 ± 3 | 48 ± 3 | 8.36 ± 0.08 | 2.383 ± 0.626 | 6.15 ± 4.90 |
| RE042 | 76 ± 2 | 36 ± 2 | 41 ± 2 | 9.67 ± 0.07 | 3.532 ± 0.647 | 12.53 ± 1.46 |
| RE043 | 76 ± 3 | 37 ± 2 | 42 ± 1 | 7.89 ± 0.11 | 2.517 ± 0.756 | 7.52 ± 5.95 |
| RE044 | 112 ± 6 | 69 ± 4 | 64 ± 2 | 6.89 ± 0.14 | 4.397 ± 0.305 | 19.90 ± 0.97 |
| RE045 | 81 ± 3 | 43 ± 1 | 43 ± 1 | 9.89 ± 0.18 | 4.912 ± 0.903 | 17.5 2 ± 11.23 |
| RE046 | 94 ± 4 | 54 ± 1 | 54 ± 2 | 7.45 ± 0.22 | 4.435 ± 0.371 | 12.40 ± 2.35 |
| RE048 | 112 ± 4 | 69 ± 2 | 64 ± 3 | 8.92 ± 0.07 | 5.008 ± 0.375 | 23.67 ± 0.13 |
| RE049 | 102 ± 2 | 56 ± 1 | 57 ± 1 | 7.12 ± 0.11 | 5.845 ± 0.153 | 25.18 ± 0.65 |
| RE050 | 93 ± 3 | 52 ± 3 | 53 ± 2 | 8.76 ± 0.14 | 4.653 ± 0.888 | 12.72 ± 2.15 |
| RE051 | 64 ± 5 | 36 ± 3 | 41 ± 3 | 9.82 ± 0.18 | 4.372 ± 0.518 | 12.97 ± 0.95 |
| RE052 | 86 ± 5 | 45 ± 2 | 49 ± 4 | 9.75 ± 0.22 | 3.150 ± 1.197 | 9.92 ± 3.03 |
| RE053 | 93 ± 4 | 47 ± 3 | 47 ± 1 | 7.56 ± 0.08 | 5.072 ± 0.719 | 12.20 ± 2.68 |
| RE054 | 97 ± 4 | 50 ± 1 | 53 ± 1 | 7.50 ± 0.07 | 2.010 ± 0.752 | 9.45 ± 2.84 |
| RE055 | 106 ± 3 | 62 ± 2 | 59 ± 3 | 8.72 ± 0.11 | 5.284 ± 0.367 | 22.90 ± 0.80 |
| RE056 | 75 ± 4 | 37 ± 1 | 39 ± 2 | 7.69 ± 0.14 | 1.968 ± 0.450 | 14.58 ± 4.82 |
| RE057 | 102 ± 3 | 56 ± 2 | 59 ± 2 | 7.98 ± 0.18 | 2.873 ± 0.475 | 19.82 ± 0.60 |
| RE058 | 67 ± 1 | 31 ± 1 | 34 ± 2 | 8.23 ± 0.22 | 5.375 ± 0.679 | 19.98 ± 5.57 |
| RE059 | 73 ± 2 | 37 ± 3 | 43 ± 2 | 7.54 ± 0.08 | 1.900 ± 0.566 | 9.05 ± 4.93 |
| RE060 | 96 ± 2 | 53 ± 1 | 52 ± 2 | 8.88 ± 0.07 | 3.485 ± 0.805 | 13.03 ± 2.99 |
| RE061 | 110 ± 2 | 68 ± 3 | 63 ± 2 | 6.56 ± 0.11 | 3.343 ± 0.296 | 18.82 ± 0.60 |
| RE062 | 82 ± 2 | 43 ± 2 | 46 ± 1 | 9.55 ± 0.14 | 2.197 ± 0.955 | 10.87 ± 2.62 |
| RE063 | 73 ± 2 | 37 ± 3 | 41 ± 1 | 8.21 ± 0.18 | 6.210 ± 0.874 | 22.83 ± 7.61 |
| RE064 | 107 ± 2 | 64 ± 1 | 60 ± 2 | 8.84 ± 0.22 | 4.056 ± 0.318 | 19.17 ± 0.23 |
| RE065 | 64 ± 1 | 32 ± 2 | 36 ± 2 | 7.71 ± 0.08 | 4.640 ± 1.043 | 14.30 ± 3.36 |
| RE066 | 100 ± 3 | 56 ± 2 | 54 ± 1 | 7.34 ± 0.07 | 5.620 ± 0.231 | 20.48 ± 0.21 |
| RE068 | 107 ± 2 | 65 ± 1 | 57 ± 2 | 7.65 ± 0.14 | 3.220 ± 0.304 | 14.37 ± 0.25 |
| RE069 | 127 ± 2 | 87 ± 1 | 78 ± 2 | 7.87 ± 0.18 | 3.473 ± 0.476 | 16.15 ± 0.52 |
| RE070 | 107 ± 1 | 65 ± 1 | 59 ± 1 | 7.45 ± 0.22 | 2.943 ± 0.927 | 16.42 ± 0.40 |
| RE071 | 111 ± 3 | 68 ± 3 | 66 ± 2 | 7.76 ± 0.08 | 5.093 ± 0.213 | 25.48 ± 1.52 |
| RE072 | 96 ± 3 | 53 ± 2 | 51 ± 2 | 7.68 ± 0.07 | 5.803 ± 0.363 | 12.87 ± 3.91 |
| RE073 | 68 ± 2 | 34 ± 2 | 36 ± 2 | 7.38 ± 0.11 | 1.670 ± 0.527 | 7.70 ± 4.45 |
| RE074 | 89 ± 2 | 48 ± 2 | 48 ± 1 | 7.98 ± 0.14 | 1.452 ± 0.414 | 7.57 ± 1.76 |
| RE075 | 87 ± 3 | 45 ± 2 | 46 ± 2 | 7.89 ± 0.18 | 6.125 ± 0.423 | 10.55 ± 8.71 |
| RE076 | 129 ± 5 | 88 ± 1 | 82 ± 1 | 8.93 ± 0.22 | 4.543 ± 0.508 | 22.77 ± 1.27 |
| RE077 | 112 ± 3 | 71 ± 2 | 67 ± 1 | 9.78 ± 0.08 | 2.628 ± 0.340 | 15.80 ± 0.18 |
| RE079 | 51 ± 2 | 28 ± 1 | 31 ± 3 | 6.98 ± 0.11 | 3.823 ± 0.078 | 13.32 ± 1.70 |
| RE080 | 82 ± 2 | 41 ± 1 | 43 ± 1 | 7.78 ± 0.14 | 1.715 ± 0.373 | 8.48 ± 1.04 |
| RE081 | 68 ± 4 | 33 ± 2 | 36 ± 3 | 7.12 ± 0.18 | 6.253 ± 0.437 | 19.13 ± 1.01 |
| RE083 | 87 ± 4 | 47 ± 4 | 49 ± 3 | 7.54 ± 0.08 | 2.453 ± 1.164 | 8.08 ± 1.81 |
| RE084 | 68 ± 3 | 33 ± 2 | 39 ± 1 | 7.66 ± 0.07 | 2.108 ± 0.798 | 7.47 ± 3.18 |
| RE085 | 69 ± 3 | 35 ± 1 | 39 ± 2 | 8.12 ± 0.11 | 2.013 ± 0.894 | 7.00 ± 3.43 |
| RE086 | 65 ± 2 | 35 ± 1 | 39 ± 2 | 9.31 ± 0.14 | 2.038 ± 0.736 | 7.33 ± 2.38 |
| RE087 | 64 ± 5 | 29 ± 1 | 35 ± 4 | 7.23 ± 0.18 | 2.365 ± 0.573 | 9.17 ± 1.37 |
| RE088 | 96 ± 3 | 54 ± 2 | 54 ± 2 | 6.99 ± 0.22 | 2.198 ± 0.778 | 8.42 ± 1.52 |
| RE089 | 85 ± 3 | 44 ± 4 | 45 ± 1 | 7.23 ± 0.08 | 2.132 ± 0.878 | 7.53 ± 3.96 |
| RE090 | 106 ± 3 | 65 ± 1 | 62 ± 1 | 7.54 ± 0.07 | 5.588 ± 0.316 | 23.57 ± 1.25 |
| RE092 | 80 ± 3 | 40 ± 2 | 43 ± 2 | 7.98 ± 0.14 | 2.295 ± 0.898 | 10,42 ± 2.93 |
| RE093 | 82 ± 4 | 42 ± 3 | 48 ± 3 | 7.54 ± 0.18 | 5.325 ± 0.158 | 15.07 ± 3.66 |
| RE097 | 59 ± 3 | 27 ± 2 | 33 ± 2 | 6.87 ± 0.18 | 4.222 ± 0.762 | 10.50 ± 3.50 |
| RE098 | 82 ± 1 | 41 ± 1 | 42 ± 1 | 7.78 ± 0.22 | 5.570 ± 0.413 | 17.17 ± 3.27 |
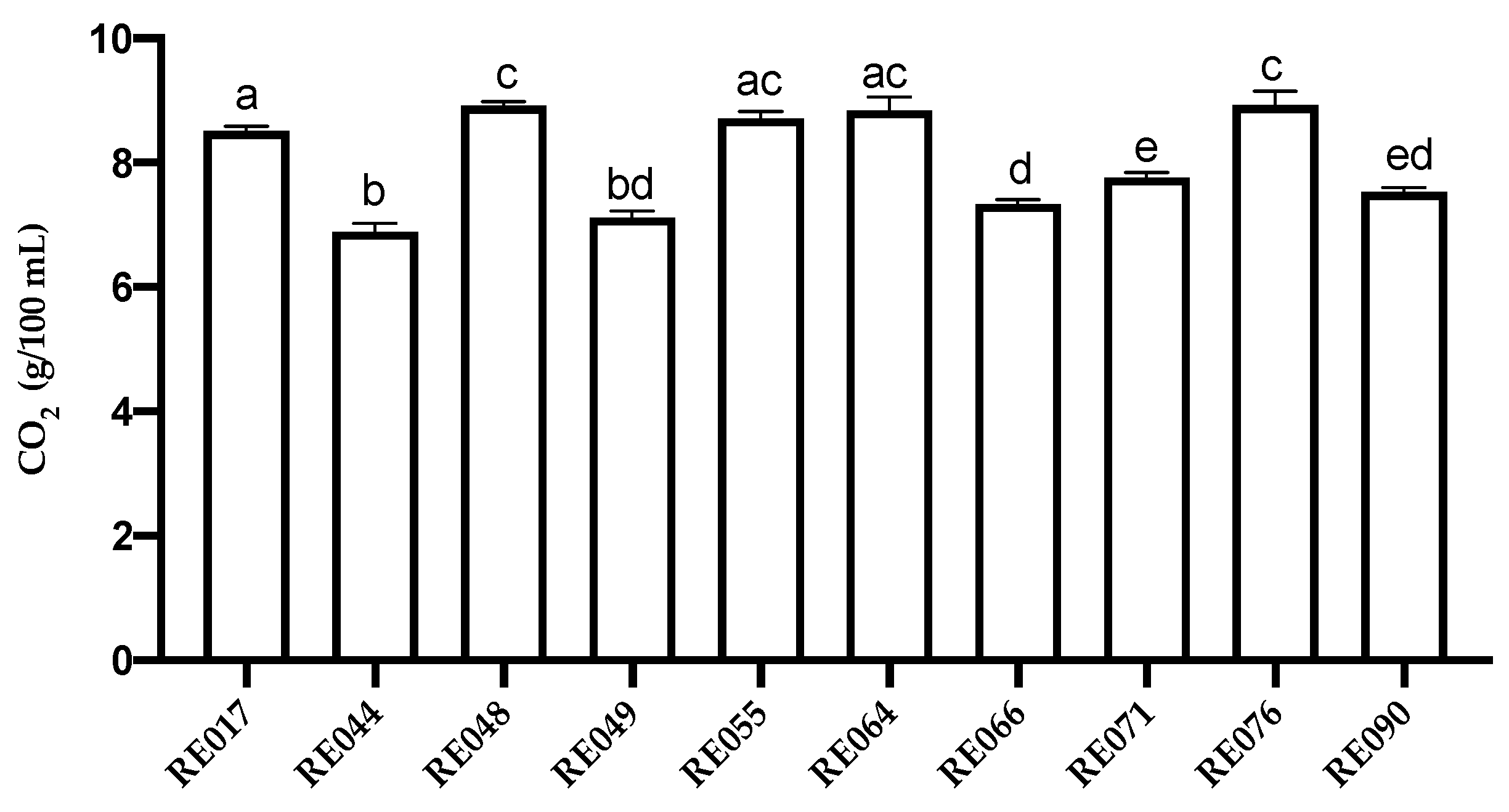
| UMCC Code | Original Code | OTA Absorption Activity (%) |
|---|---|---|
| UMCC 2954 | RE017 | 42.03 ± 7.33 |
| UMCC 2955 | RE044 | 41.54 ± 4.98 |
| UMCC 2956 | RE048 | 40.01 ± 7.08 |
| UMCC 2957 | RE049 | 48.96 ± 4.83 |
| UMCC 2958 | RE055 | 40.90 ± 6.18 |
| UMCC 2959 | RE064 | 34.52 ± 1.80 |
| UMCC 2960 | RE066 | 45.72 ± 1.95 |
| UMCC 2961 | RE071 | 46.42 ± 6.75 |
| UMCC 2962 | RE076 | 45.23 ± 8.28 |
| UMCC 2963 | RE090 | 44.75 ± 2.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulvirenti, A.; De Vero, L.; Blaiotta, G.; Sidari, R.; Iosca, G.; Gullo, M.; Caridi, A. Selection of Wine Saccharomyces cerevisiae Strains and Their Screening for the Adsorption Activity of Pigments, Phenolics and Ochratoxin A. Fermentation 2020, 6, 80. https://doi.org/10.3390/fermentation6030080
Pulvirenti A, De Vero L, Blaiotta G, Sidari R, Iosca G, Gullo M, Caridi A. Selection of Wine Saccharomyces cerevisiae Strains and Their Screening for the Adsorption Activity of Pigments, Phenolics and Ochratoxin A. Fermentation. 2020; 6(3):80. https://doi.org/10.3390/fermentation6030080
Chicago/Turabian StylePulvirenti, Andrea, Luciana De Vero, Giuseppe Blaiotta, Rossana Sidari, Giovanna Iosca, Maria Gullo, and Andrea Caridi. 2020. "Selection of Wine Saccharomyces cerevisiae Strains and Their Screening for the Adsorption Activity of Pigments, Phenolics and Ochratoxin A" Fermentation 6, no. 3: 80. https://doi.org/10.3390/fermentation6030080