
Lactic Acid Bacteria and Formic Acid Improve Fermentation Quality and Beneficial Predicted Functional Characteristics in Mixed Silage Consisting of Alfalfa and Perennial Ryegrass
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Preparation
2.2. Analysis of Microbial Populations, Chemical Compositions, and Fermentation Quality
2.3. Bacterial Community Analysis
2.4. Predicted Metabolic Function Analysis
2.5. Statistical Analysis
3. Results
3.1. Chemical Composition of Mixed Silage with Alfalfa and Perennial Ryegrass during the Ensiling Process
3.2. Fermentation Characteristics of Mixed Alfalfa and Perennial Ryegrass Silage during the Ensiling Process
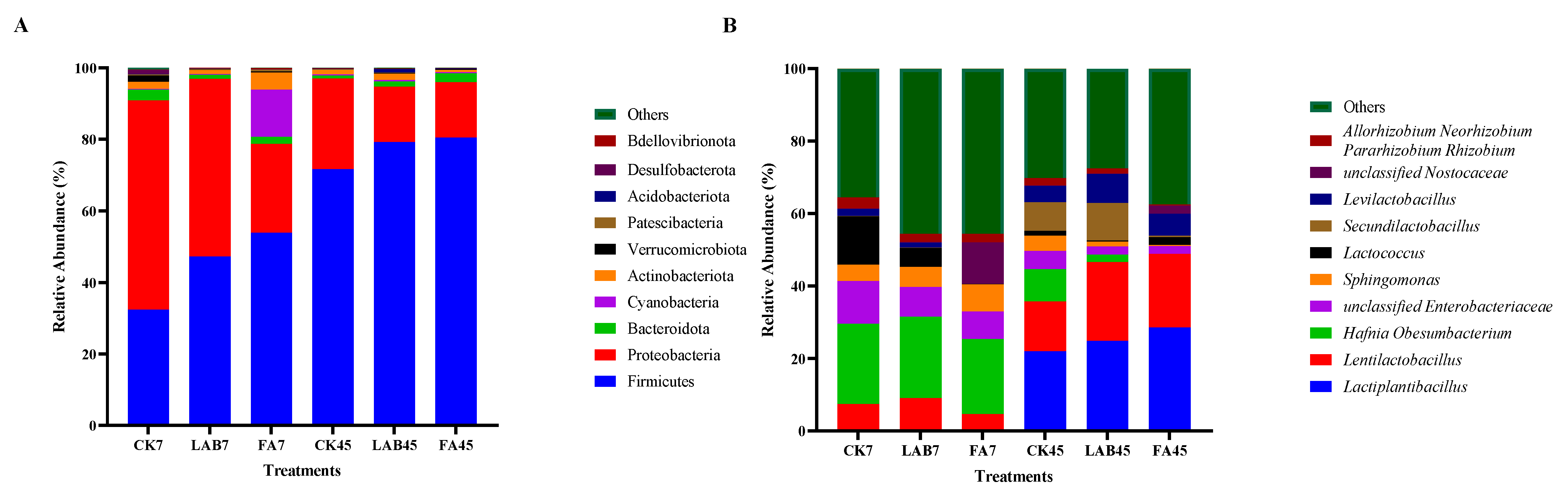
3.3. Dynamic Changes in the Microbial Community of Mixed Alfalfa and Perennial Ryegrass Silage during the Ensiling Process
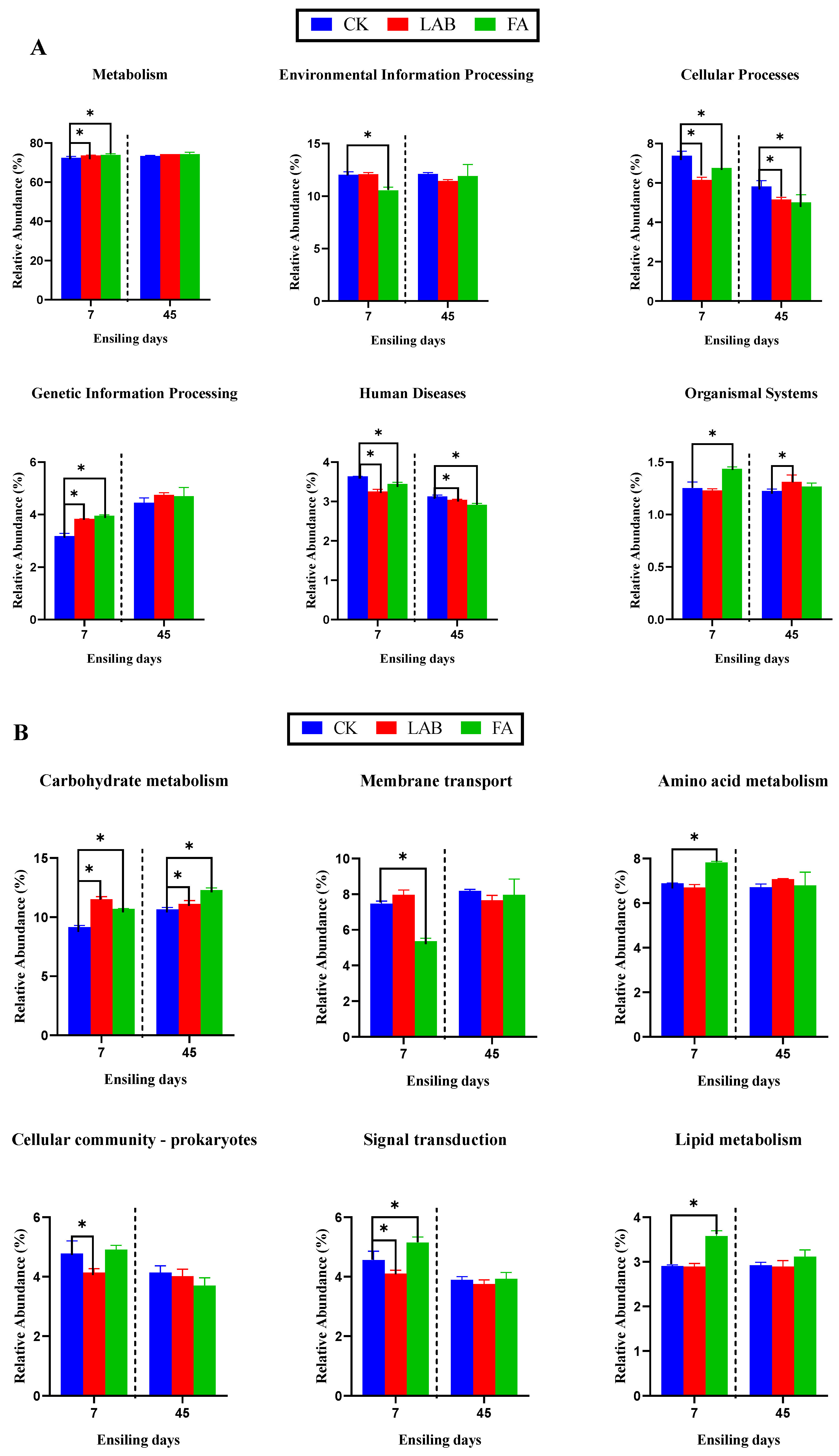
3.4. Predicted Metabolic Functions of the Bacterial Community of Mixed Silage with Alfalfa and Perennial Ryegrass during the Ensiling Process
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lan, J.; Hu, N.; Fu, W. Soil carbon-nitrogen coupled accumulation following the natural vegetation restoration of abandoned farmlands in a karst rocky desertification region. Ecol. Eng. 2020, 158, 106033. [Google Scholar] [CrossRef]
- Peng, X.; Dai, Q.; Ding, G.; Shi, D.; Li, C. Impact of vegetation restoration on soil properties in near-surface fissures located in karst rocky desertification regions. Soil Tillage Res. 2020, 200, 104620. [Google Scholar] [CrossRef]
- Long, Y.; Yue, K.; Gu, Z.; Chen, H.; Chi, Y. Analysis of rainfall factors and soil erosion in different soil and water conservation measures in the karst plateau-mountain. Pol. J. Environ. Stud. 2021, 30, 5343–5349. [Google Scholar]
- Zhang, J.; Guo, G.; Chen, L.; Li, J.; Yuan, X.; Yu, C.; Shimojo, M.; Shao, T. Effect of applying lactic acid bacteria and propionic acid on fermentation quality and aerobic stability of oats-common vetch mixed silage on the tibetan plateau. Anim. Sci. J. 2015, 86, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Wu, X.; Chen, M.; Wang, Y.; Liu, X.; Gong, P.; Xu, Q.; Wang, X.; Gao, H.; Wang, Z. Identification of genetic loci associated with crude protein and mineral concentrations in alfalfa (Medicago sativa) using association mapping. BMC Plant Biol. 2017, 17, 97. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.H.; Dong, Z.H.; Shao, T. Effect of additives on fatty acid profile of high moisture alfalfa silage during ensiling and after exposure to air. Anim. Feed Sci. Technol. 2018, 236, 29–38. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Yu, Z. Fermentation dynamics and bacterial diversity of mixed lucerne and sweet corn stalk silage ensiled at six ratios. Grass Forage Sci. 2019, 74, 264–273. [Google Scholar] [CrossRef]
- Yu, X.; Pijut, P.M.; Byrne, S.; Asp, T.; Bai, G.; Jiang, Y. Candidate gene association mapping for winter survival and spring regrowth in perennial ryegrass. Plant Sci. 2015, 235, 37–45. [Google Scholar] [CrossRef]
- Luo, Y.; Xing, R.; Wan, Z.; Chen, Y. Vertical distribution of nutrients, enzyme activities, microbial properties, and heavy metals in zinc smelting slag site revegetated with two herb species: Implications for direct revegetation. Sci. Total Environ. 2023, 879, 163206. [Google Scholar] [CrossRef]
- Dong, D.; Xu, G.; Dai, T.; Zong, C.; Yin, X.; Bao, Y.; Shao, T. Effect of molasses on fermentation quality of wheat straw ensiled with perennial ryegrass. Anim. Prod. Sci. 2022, 62, 1471–1479. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, H.; Gao, Y.; Diao, Q. Dynamic profiles of fermentation characteristics and bacterial community composition of Broussonetia papyrifera ensiled with perennial ryegrass. Bioresour. Technol. 2020, 310, 123396. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, Y.; Gou, W.; Cheng, Q.; Bai, S.; Cai, Y. Silage fermentation and bacterial community of bur clover, annual ryegrass and their mixtures prepared with microbial inoculant and chemical additive. Anim. Feed Sci. Technol. 2019, 247, 285–293. [Google Scholar] [CrossRef]
- Fan, X.; Xie, Z.; Cheng, Q.; Li, M.; Long, J.; Lei, Y.; Jia, Y.; Chen, Y.; Chen, C.; Wang, Z. Fermentation quality, bacterial community, and predicted functional profiles in silage prepared with alfalfa, perennial ryegrass and their mixture in the karst region. Front. Microbiol. 2022, 13, 1062515. [Google Scholar] [CrossRef] [PubMed]
- Mpanza, T.D.E.; Mani, S. Effects of Vachellia mearnsii Tannin Extract as an Additive on Fermentation Quality, Aerobic Stability, and Microbial Modulation of Maize Silage. Microorganisms 2023, 11, 2767. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, J.; Shi, W.; Sun, J.; Xia, T.; Huang, F.; Liu, Y.; Li, H.; Teng, K.; Zhong, J. Dynamic changes in fermentation quality and structure and function of the microbiome during mixed silage of sesbania cannabina and sweet sorghum grown on saline-alkaline land. Microbiol. Spectr. 2022, 10, e0248322. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.R.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy. Sci. 2018, 1015, 3980–4000. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60ts-2, with or without homofermentative lactobacillus plantarum b90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Wen, A.; Yuan, X.; Wang, J.; Desta, S.T.; Shao, T. Effects of four short-chain fatty acids or salts on dynamics of fermentation and microbial characteristics of alfalfa silage. Anim. Feed Sci. Technol. 2017, 223, 141–148. [Google Scholar] [CrossRef]
- Bai, J.; Ding, Z.; Su, R.; Wang, M.; Cheng, M.; Xie, D.; Guo, X. Storage temperature is more effective than lactic acid bacteria inoculations in manipulating fermentation and bacterial community diversity, co-occurrence and functionality of the whole-plant corn silage. Microbiol. Spectr. 2022, 10, e101–e122. [Google Scholar] [CrossRef]
- Wang, S.; Shao, T.; Li, J.; Zhao, J.; Dong, Z. Fermentation profiles, bacterial community compositions, and their predicted functional characteristics of grass silage in response to epiphytic microbiota on legume forages. Front. Microbiol. 2020, 13, 830888. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wan, C.; Ma, Y.; Zhang, K.; Wang, F.; Shen, S. Study on the Quality of Mixed Silage of Rapeseed with Alfalfa or Myriophyllum. Int. J. Environ. Res. Public Health 2023, 20, 3884. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; He, L.W.; Xing, Y.Q.; Zhou, W.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics and aerobic deterioration of silage. J. Dairy. Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Owens, V.; Albrecht, K.A.; Muck, R.E.; Duke, S.H. Protein degradation and fermentation characteristics of red clover and alfalfa silage harvested with varying levels of total nonstructural carbohydrates. Crop Sci. 1999, 39, 1873–1880. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; AOAC: Rockville, MD, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy. Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media1. J. Dairy. Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Jia, T.; Sun, Z.; Gao, R.; Yu, Z. Lactic acid bacterial inoculant effects on the vitamin content of alfalfa and chinese leymus silage. Asian Australas. J. Anim. Sci. 2019, 32, 1873–1881. [Google Scholar]
- Li, P.; Lu, Y.; Zhao, M.; Chen, L.; Zhang, C.; Cheng, Q.; Chen, C. Effects of phenyllactic acid, lactic acid bacteria, and their mixture on fermentation characteristics and microbial community composition of timothy silage. Front. Microbiol. 2021, 12, 743433. [Google Scholar] [CrossRef]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4fun: Predicting functional profiles from metagenomic 16s rrna data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Yuan, X.J.; Wen, A.Y.; Wang, J.; Desta, S.T.; Dong, Z.H.; Shao, T. Effects of four short-chain fatty acids or salts on fermentation characteristics and aerobic stability of alfalfa (Medicago sativa L.) Silage. J. Sci. Food. Agric. 2018, 98, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Nazar, M.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Kaka, N.A.; Shao, T. The feasibility and effects of exogenous epiphytic microbiota on the fermentation quality and microbial community dynamics of whole crop corn. Bioresour. Technol. 2020, 306, 123106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Z.; Na, R. Effects of different additives on fermentation quality and aerobic stability of leymus chinensis silage. Grass Forage Sci. 2018, 73, 413–419. [Google Scholar] [CrossRef]
- Wei, S.N.; Li, Y.F.; Jeong, E.C.; Kim, H.J.; Kim, J.G. Effects of formic acid and lactic acid bacteria inoculant on main summer crop silages in korea. J. Anim. Sci. Technol. 2021, 63, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.C.; Bernardes, T.F.; Roth, A.P.D.T.P.; Lodo, B.N.; Berchielli, T.T.; Reis, R.A. Fermentation and aerobic stability of corn silage inoculated with Lactobacillus buchneri. Rev. Bras. Zootecn. 2012, 41, 1789–1794. [Google Scholar] [CrossRef]
- Mcdonald, P.; Henderson, A.R.; Heron, S. The Biochemistry of Silage; John Wiley & Sons: Chichester, UK, 1991. [Google Scholar]
- Dong, M.; Li, Q.; Xu, F.; Wang, S.; Li, W. Effects of microbial inoculants on the fermentation characteristics and microbial communities of sweet sorghum bagasse silage. Sci. Rep. 2020, 10, 837. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Wang, S.; Sun, Y.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Assessment of inoculating various epiphytic microbiota on fermentative profile and microbial community dynamics in sterile italian ryegrass. J. Appl. Microbiol. 2020, 129, 509–520. [Google Scholar] [CrossRef]
- Zhao, S.; Yang, F.; Wang, Y.; Fan, X.; Feng, C.; Wang, Y. Dynamics of fermentation parameters and bacterial community in high-moisture alfalfa silage with or without lactic acid bacteria. Microorganisms 2021, 9, 1225. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in southwest china. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Xian, S.; Liu, X.; Shen, G.; Zhang, Z.; Hou, X.; Chen, A. Metagenomic study on chinese homemade paocai: The effects of raw materials and fermentation periods on the microbial ecology and volatile components. Foods. 2022, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Li, M.; Fan, X.; Cheng, Q.; Chen, Y.; Long, J.; Lei, Y.; Li, P.; Chen, C. Effect of amomum villosum essential oil as an additive on the chemical composition, fermentation quality, and bacterial community of paper mulberry silage. Front. Microbiol. 2022, 13, 951958. [Google Scholar] [CrossRef] [PubMed]
- Tsialtas, I.T.; Baxevanos, D.; Vlachostergios, D.N.; Dordas, C.; Lithourgidis, A. Cultivar complementarity for symbiotic nitrogen fixation and water use efficiency in pea-oat intercrops and its effect on forage yield and quality. Field Crops Res. 2018, 226, 28–37. [Google Scholar] [CrossRef]
- Du, Z.; Lin, Y.; Sun, L.; Yang, F.; Cai, Y. Microbial community structure, co-occurrence network and fermentation characteristics of woody plant silage. J. Sci. Food. Agric. 2022, 102, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Saldinger, S.S. Bacterial dynamics of wheat silage. Front. Microbiol. 2019, 10, 01532. [Google Scholar] [CrossRef]
- Li, J.W.; Meng, Q.W.; Wang, C.; Song, C.S.; Lyu, Y.; Li, J.P.; Shan, A. The interaction between temperature and citric acid treatment in the anaerobic fermentation of chinese cabbage waste. J. Clean. Prod. 2023, 383, 135502. [Google Scholar] [CrossRef]
- Fu, Z.; Sun, L.; Wang, Z.; Liu, J.; Hou, M.; Lu, Q.; Hao, J.; Jia, Y.; Ge, G. Effects of growth stage on the fermentation quality, microbial community, and metabolomic properties of italian ryegrass (Lolium multiflorum Lam.) Silage. Front. Microbiol. 2022, 13, 1054612. [Google Scholar] [CrossRef]
- Singh, V.K.; Mishra, A.; Jha, B. Anti-quorum sensing and anti-biofilm activity of delftia tsuruhatensis extract by attenuating the quorum sensing-controlled virulence factor production in pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2019, 9, 00308. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, N. Two-Component Systems. In Fundamentals of Bacterial Physiology and Metabolism; Gupta, R., Gupta, N., Eds.; Springer: Singapore, 2021; Volume 2021, pp. 557–573. [Google Scholar]
- Wu, C.; Sun, W.; Huang, Y.; Dai, S.; Peng, C.; Zheng, Y.; Chen, C.; Hao, J. Effects of different additives on the bacterial community and fermentation mode of whole-plant paper mulberry silage. Front. Microbiol. 2022, 13, 904193. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhu, B.; Li, F.; Zhang, D.; Guo, T.; Li, F.; Yang, G. The destruction of the anaerobic environment caused by rumen fistula surgery leads to differences in the rumen microbial diversity and function of sheep. Front. Vet. Sci. 2021, 8, 754195. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Treatment | Ensiling Period (D) | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| (T) | Day 7 | Day 15 | Day 45 | T | D | T × D | ||
| DM | CK | 21.09 Ab | 17.78 Bb | 17.29 Bb | ||||
| %FM | LAB | 20.71 Ab | 17.41 Bb | 17.27 Bb | 0.100 | <0.001 | <0.001 | 0.827 |
| FA | 22.36 Aa | 18.90 Ba | 18.70 Ba | |||||
| CP | CK | 16.62 Ac | 16.48 Ac | 15.70 Ba | ||||
| %DM | LAB | 18.13 Ab | 17.29 Ab | 16.05 Ba | 0.082 | <0.001 | <0.001 | 0.002 |
| FA | 19.39 Aa | 18.75 Aa | 16.12 Ba | |||||
| NDF | CK | 56.76 Aa | 51.87 Ba | 51.62 Ba | ||||
| %DM | LAB | 51.63 Ab | 49.48 Bb | 46.53 Cb | 0.244 | <0.001 | <0.001 | 0.041 |
| FA | 48.84 Ab | 48.57 Ab | 46.63 Bb | |||||
| ADF | CK | 46.79 Aa | 42.84 Ba | 37.96 Ca | ||||
| %DM | LAB | 42.13 Ab | 35.78 Bb | 34.59 Cb | 0.343 | <0.001 | <0.001 | 0.222 |
| FA | 43.40 Ab | 34.40 Bb | 34.13 Cb | |||||
| WSC | CK | 3.42 Ab | 2.70 Ab | 1.69 Bc | ||||
| %DM | LAB | 3.50 Ab | 2.46 Bb | 2.46 Bb | 0.056 | <0.001 | <0.001 | 0.034 |
| FA | 5.04 Aa | 4.07 Ba | 4.04 Ba | |||||
| Items | Treatment | Ensiling Period (D) | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| (T) | Day 7 | Day 15 | Day 45 | T | D | T × D | ||
| pH | CK | 4.96 Aa | 4.83 ABa | 4.67 Ba | ||||
| LAB | 4.59 Ab | 4.58 Abb | 4.47 Bb | 0.022 | <0.001 | 0.048 | 0.232 | |
| FA | 4.14 Ac | 4.07 ABc | 4.06 Bc | |||||
| LA | CK | 0.24 Cc | 3.62 Bc | 4.35 Ac | ||||
| %DM | LAB | 1.81 Cb | 4.53 Bb | 5.71 Ab | 0.103 | <0.001 | <0.001 | 0.288 |
| FA | 2.03 Ca | 5.48 Ba | 6.22 Aa | |||||
| AA | CK | 0.14 Bc | 1.52 Ab | 1.54 Ab | ||||
| %DM | LAB | 2.15 Aa | 2.20 Aab | 2.91 Aa | 0.091 | <0.001 | <0.001 | 0.099 |
| FA | 1.22 Bb | 2.75 Aa | 2.47 Aa | |||||
| PA | CK | 0.24 | 0.21 | - | ||||
| %DM | LAB | 0.18 | - | - | - | - | - | - |
| FA | - | - | - | |||||
| BA | CK | - | - | - | ||||
| %DM | LAB | - | - | - | - | - | - | - |
| FA | - | - | - | |||||
| NH3-N | CK | 2.82 Ca | 3.56 ABa | 4.36 Aa | ||||
| %DM | LAB | 1.86 Cb | 2.12 Bb | 3.17 Ab | 0.109 | <0.001 | 0.001 | 0.646 |
| FA | 1.57 Cb | 2.45 Bb | 2.58 Ab | |||||
| Items | Treatment | Ensiling Period (D) | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| (T) | Day 7 | Day 45 | T | D | T × D | ||
| Observed species | CK | 529 a | 465 a | ||||
| LAB | 470 b | 445 a | 2.790 | 0.003 | <0.001 | <0.001 | |
| FA | 465 b | 446 a | |||||
| ACE | CK | 559.06 a | 476.53 a | ||||
| LAB | 486.48 c | 479.33 a | 2.059 | <0.001 | <0.001 | <0.001 | |
| FA | 517.94 b | 490.50 a | |||||
| Chao1 | CK | 564.95 a | 491.34 a | ||||
| LAB | 514.31 b | 497.33 a | 2.184 | 0.003 | <0.001 | 0.001 | |
| FA | 532.39 b | 488.77 a | |||||
| Simpson | CK | 0.92 ab | 0.90 a | ||||
| LAB | 0.86 b | 0.75 b | 0.008 | <0.001 | <0.001 | <0.001 | |
| FA | 0.96 a | 0.67 c | |||||
| Shannon | CK | 5.12 b | 4.51 a | ||||
| LAB | 4.48 b | 3.44 a | 0.0141 | 0.040 | <0.001 | 0.019 | |
| FA | 6.24 a | 3.44 a | |||||
| Coverage | CK | 0.998 | 0.997 | ||||
| LAB | 0.998 | 0.999 | |||||
| FA | 0.997 | 0.998 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Y.; Li, M.; Liu, Y.; Wang, J.; He, X.; Zhao, Y.; Chen, Y.; Cheng, Q.; Chen, C. Lactic Acid Bacteria and Formic Acid Improve Fermentation Quality and Beneficial Predicted Functional Characteristics in Mixed Silage Consisting of Alfalfa and Perennial Ryegrass. Fermentation 2024, 10, 43. https://doi.org/10.3390/fermentation10010043
Lei Y, Li M, Liu Y, Wang J, He X, Zhao Y, Chen Y, Cheng Q, Chen C. Lactic Acid Bacteria and Formic Acid Improve Fermentation Quality and Beneficial Predicted Functional Characteristics in Mixed Silage Consisting of Alfalfa and Perennial Ryegrass. Fermentation. 2024; 10(1):43. https://doi.org/10.3390/fermentation10010043
Chicago/Turabian StyleLei, Yao, Maoya Li, Yinghao Liu, Jiachuhan Wang, Xiangjiang He, Yuanyuan Zhao, Yulian Chen, Qiming Cheng, and Chao Chen. 2024. "Lactic Acid Bacteria and Formic Acid Improve Fermentation Quality and Beneficial Predicted Functional Characteristics in Mixed Silage Consisting of Alfalfa and Perennial Ryegrass" Fermentation 10, no. 1: 43. https://doi.org/10.3390/fermentation10010043