
The Androgen Regulated lncRNA NAALADL2-AS2 Promotes Tumor Cell Survival in Prostate Cancer
 , and
, and
Abstract
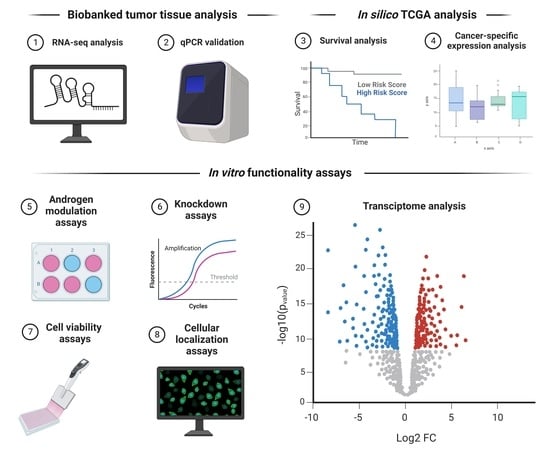
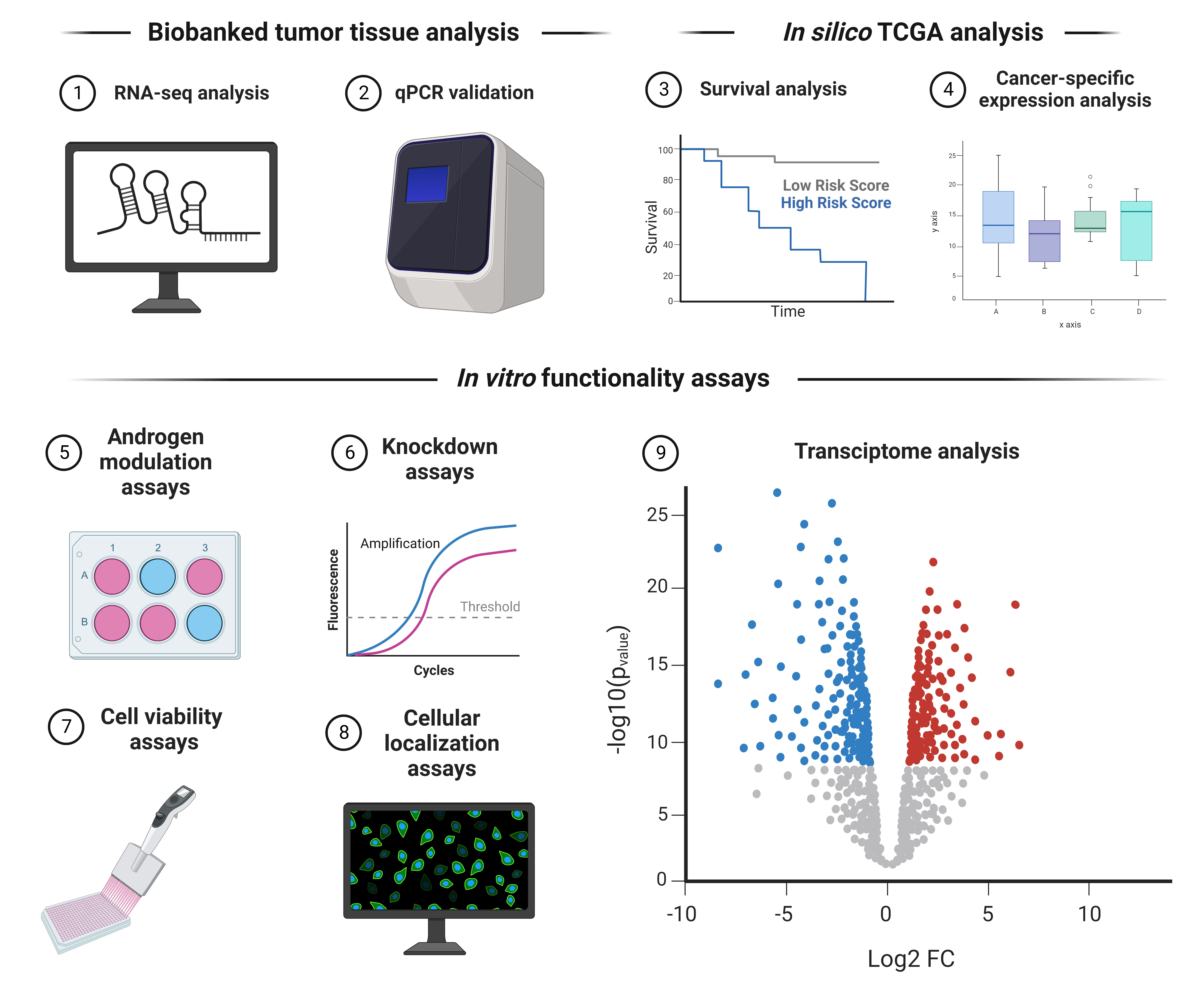
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
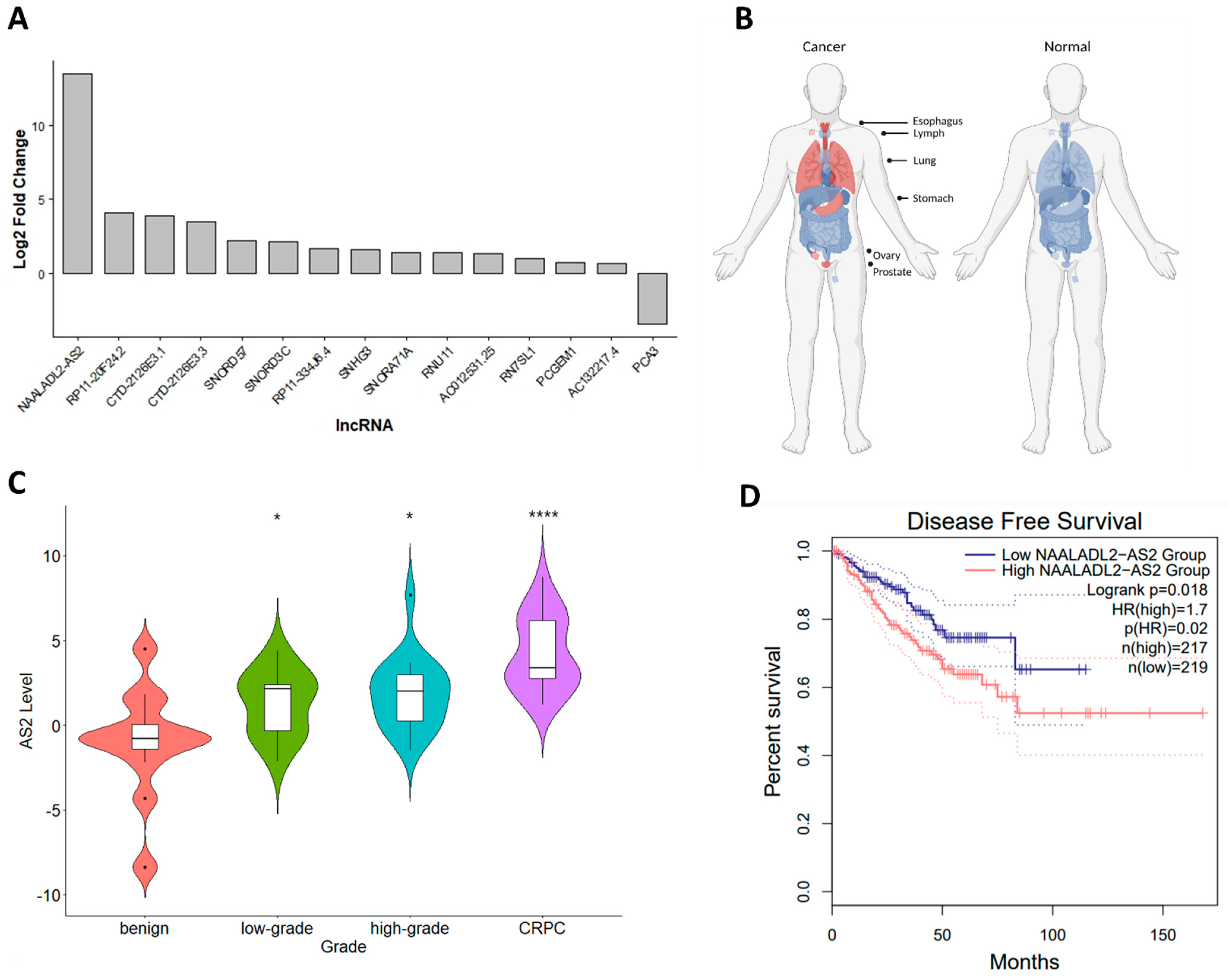
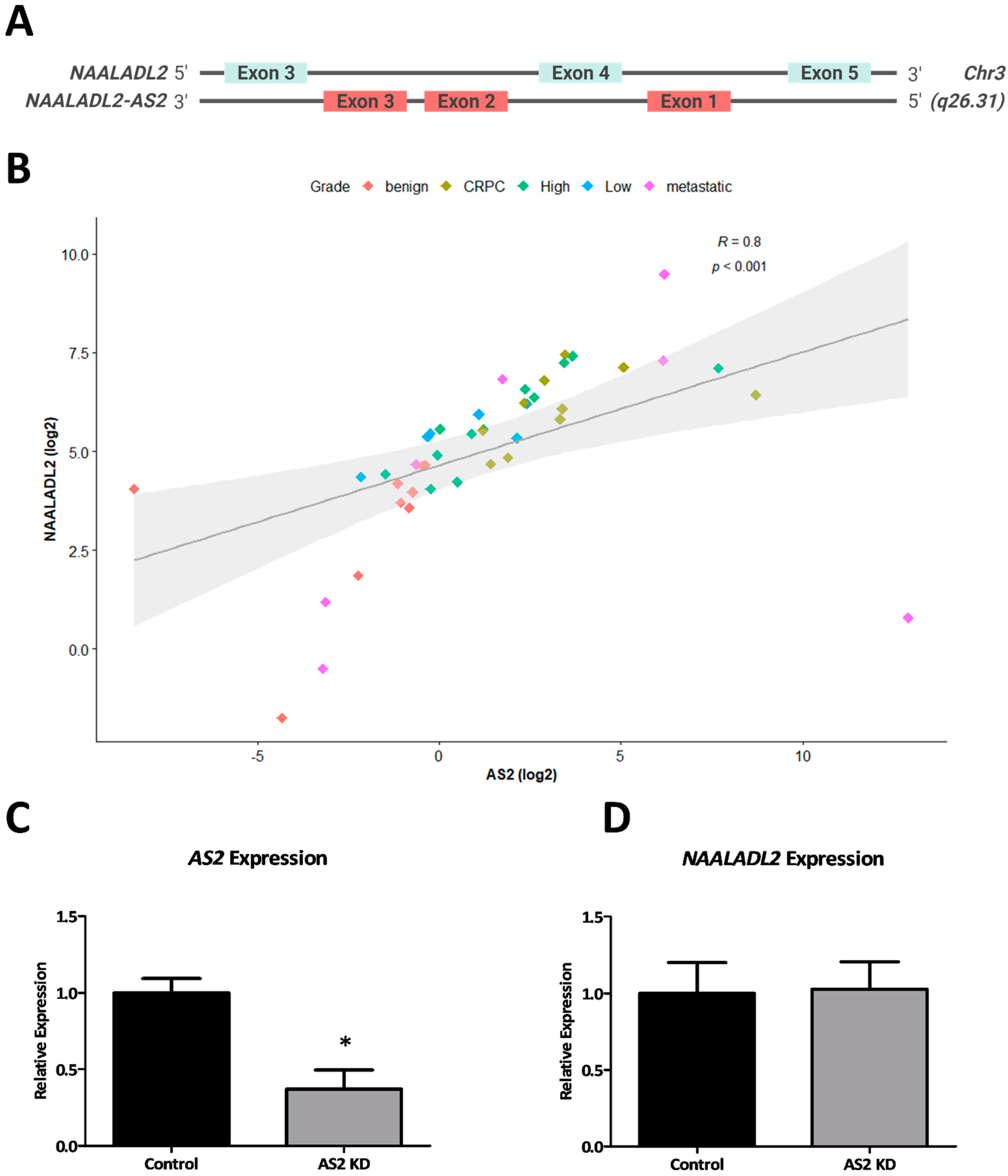
2.1. AS2 Is Overexpressed in CRPC
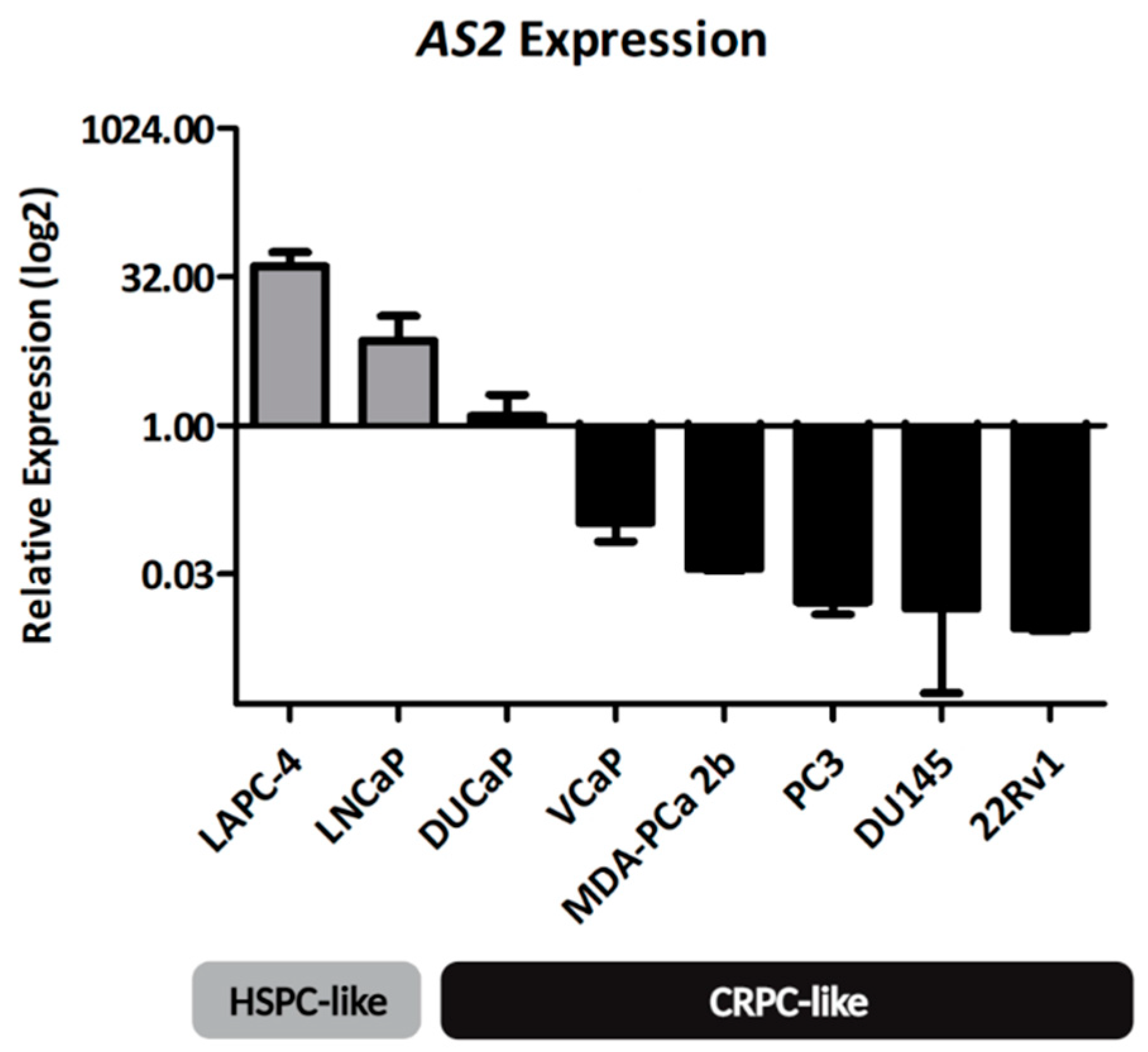
2.1.1. AS2 Expression in PCa Cell Lines
2.1.2. AS2 Is Co-Expressed with NAALADL2
2.1.3. Copy Number Gains around the AS2 Locus Are Highly Prevalent in PCa
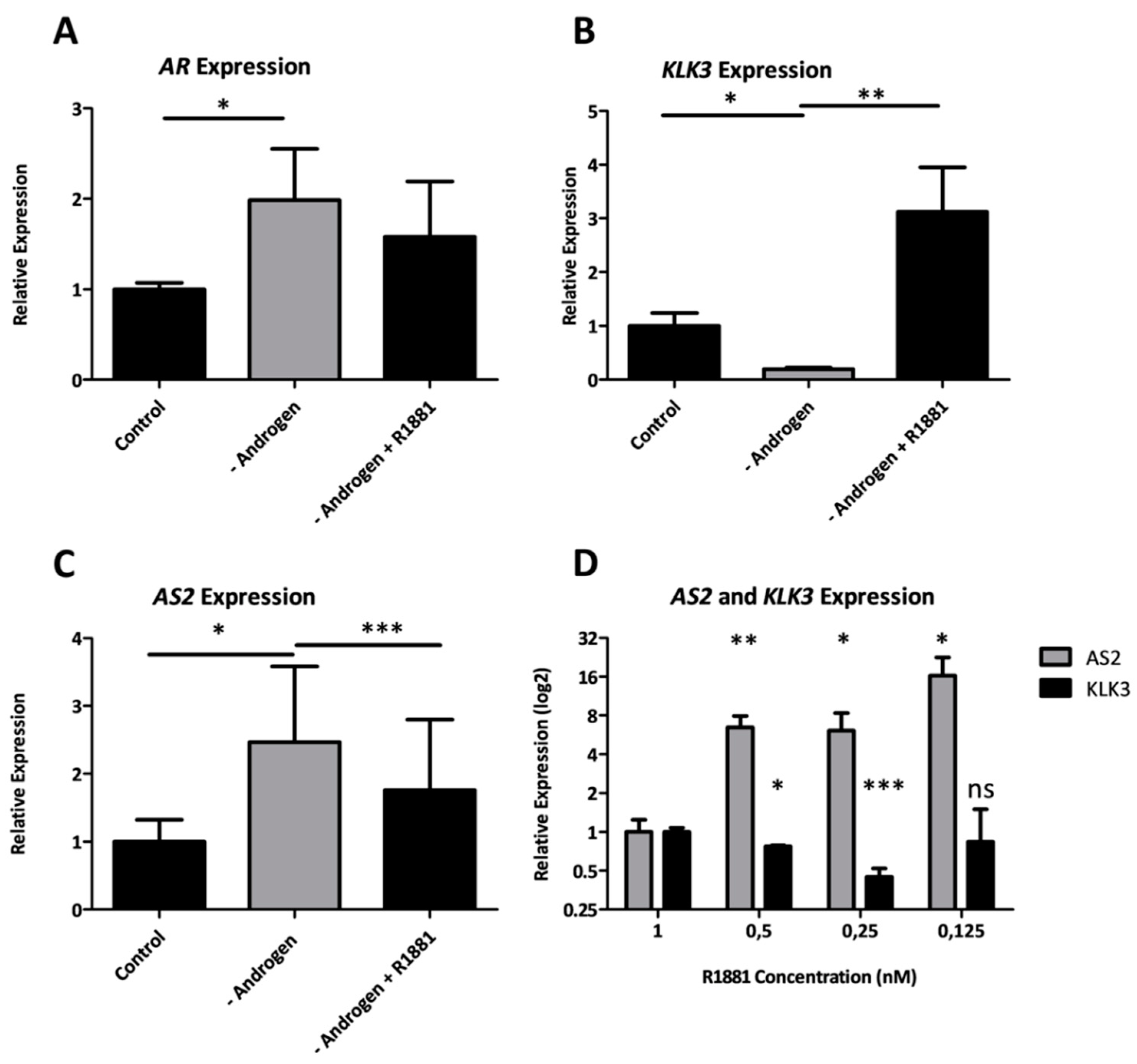
2.1.4. Androgens Regulate the Expression of AS2
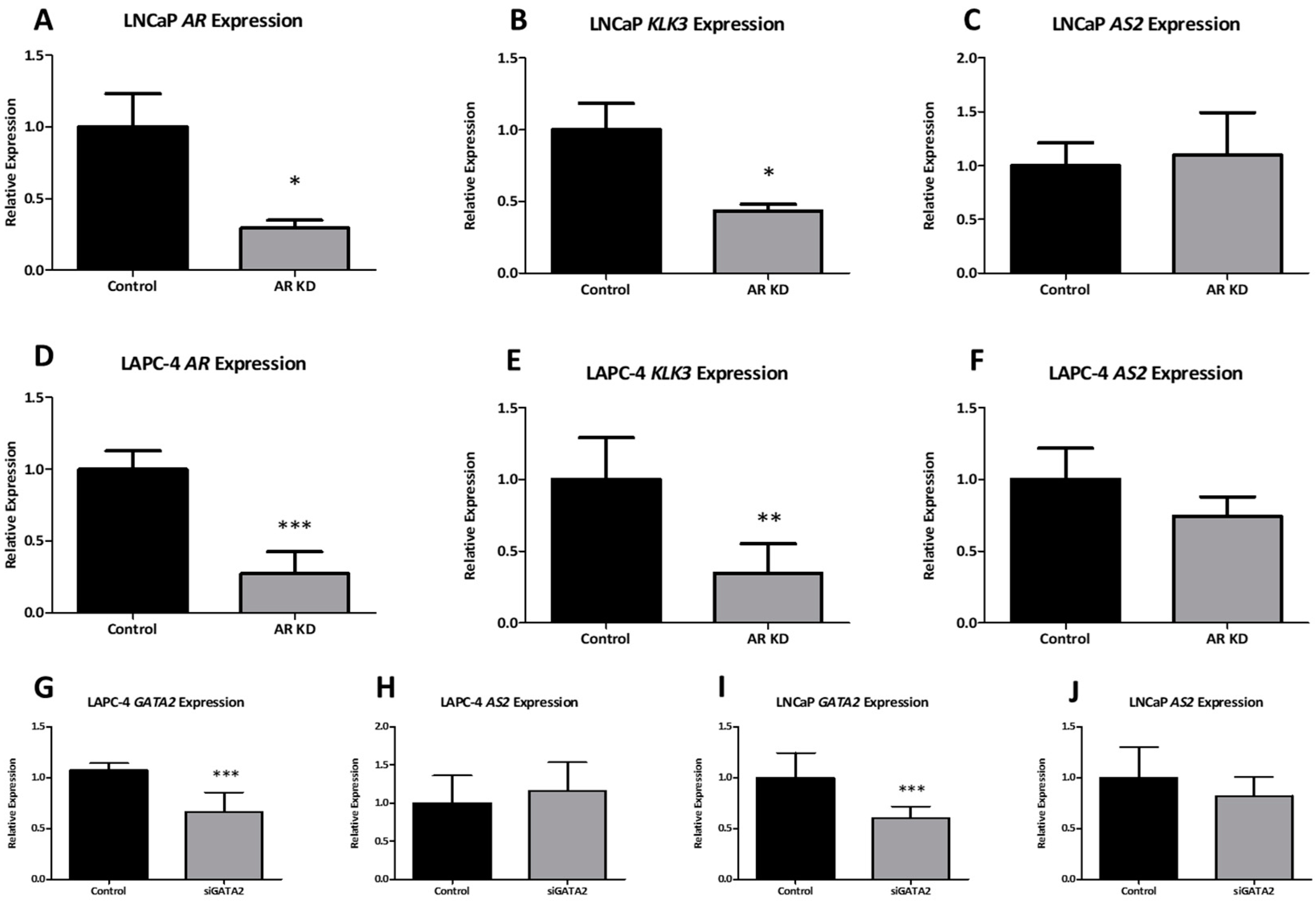
2.1.5. AR Does Not Directly Regulate AS2 Expression
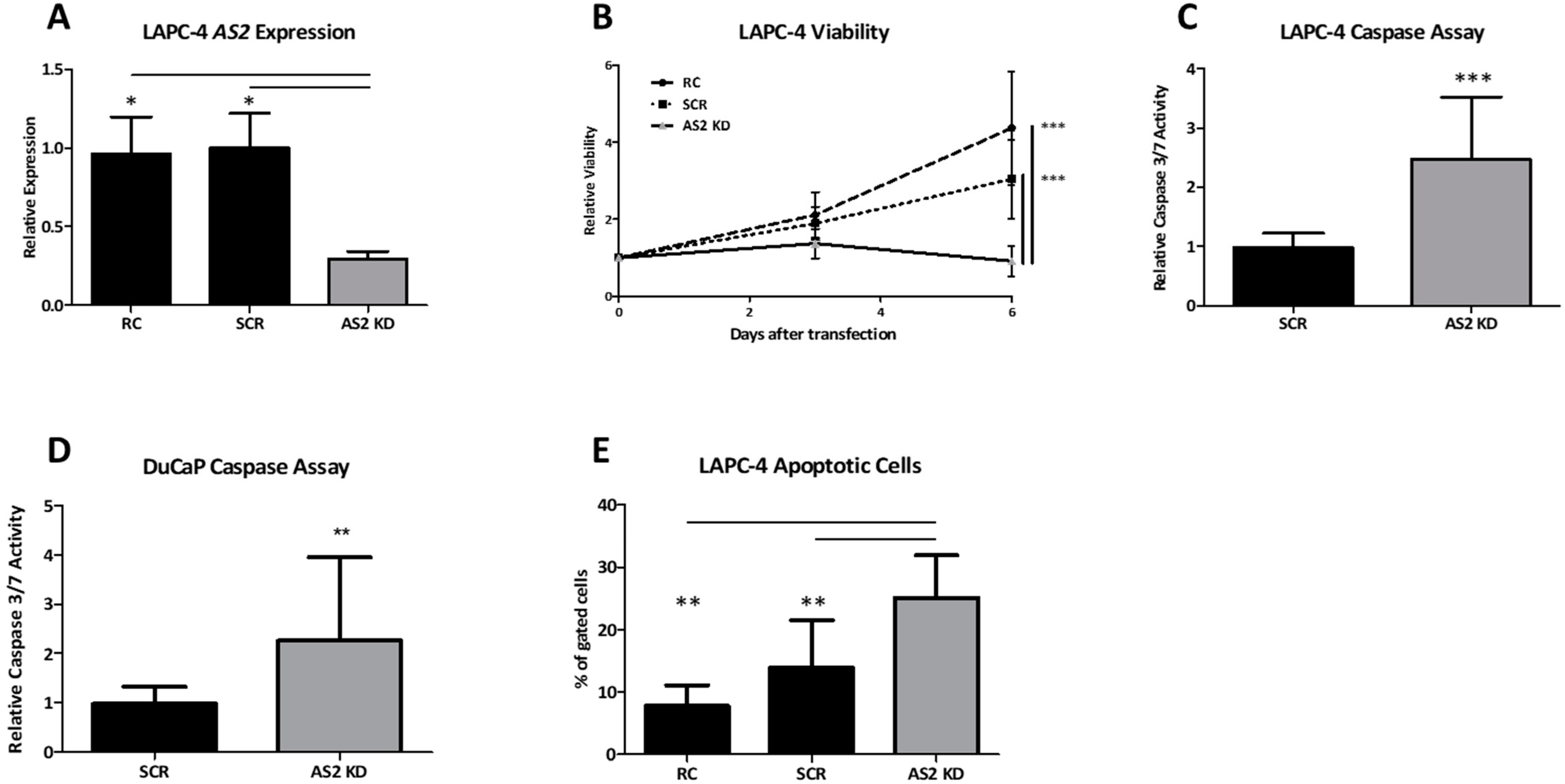
2.1.6. AS2 Promotes Survival by Inhibition of Apoptosis
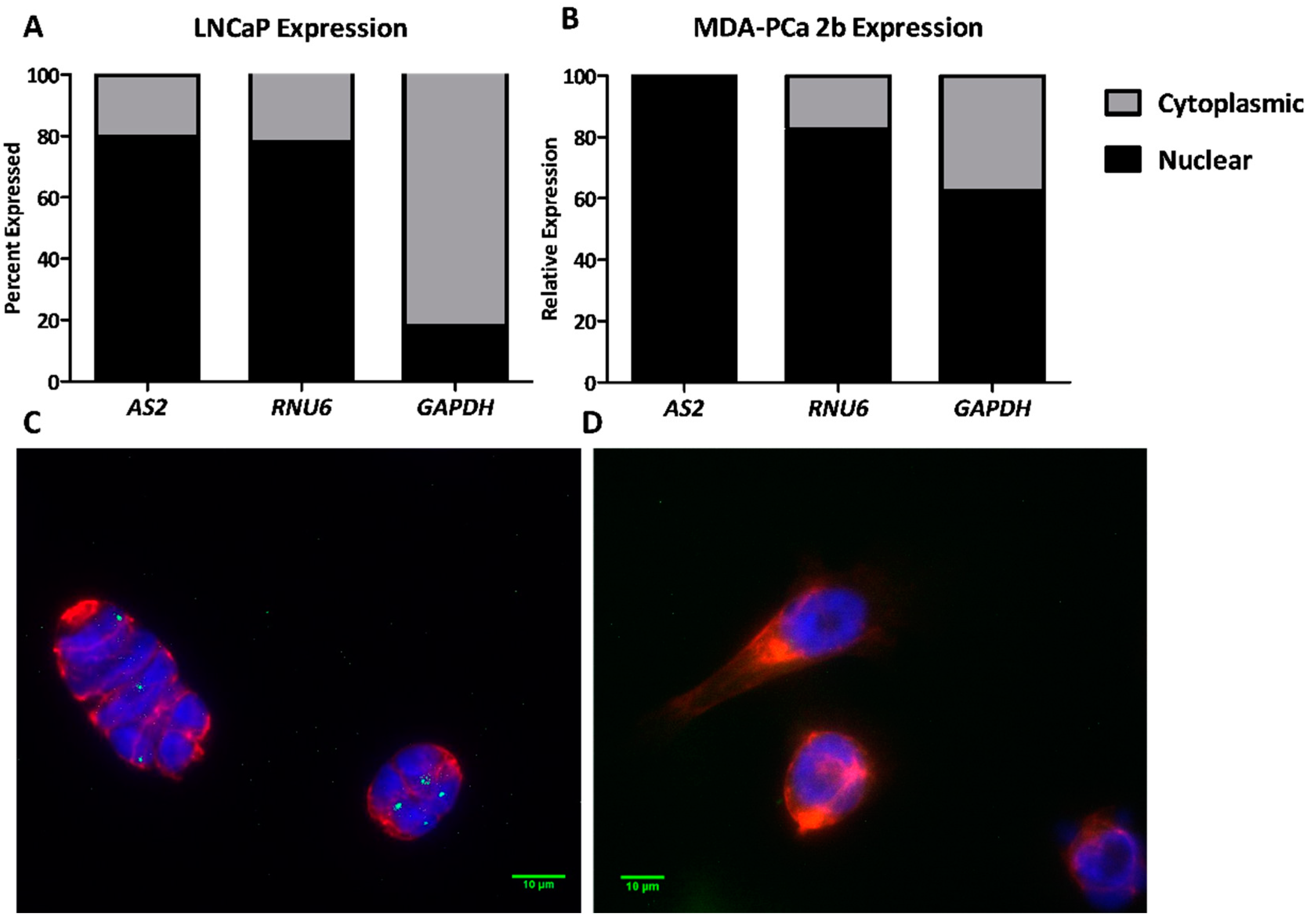
2.1.7. AS2 Is a Nuclear RNA
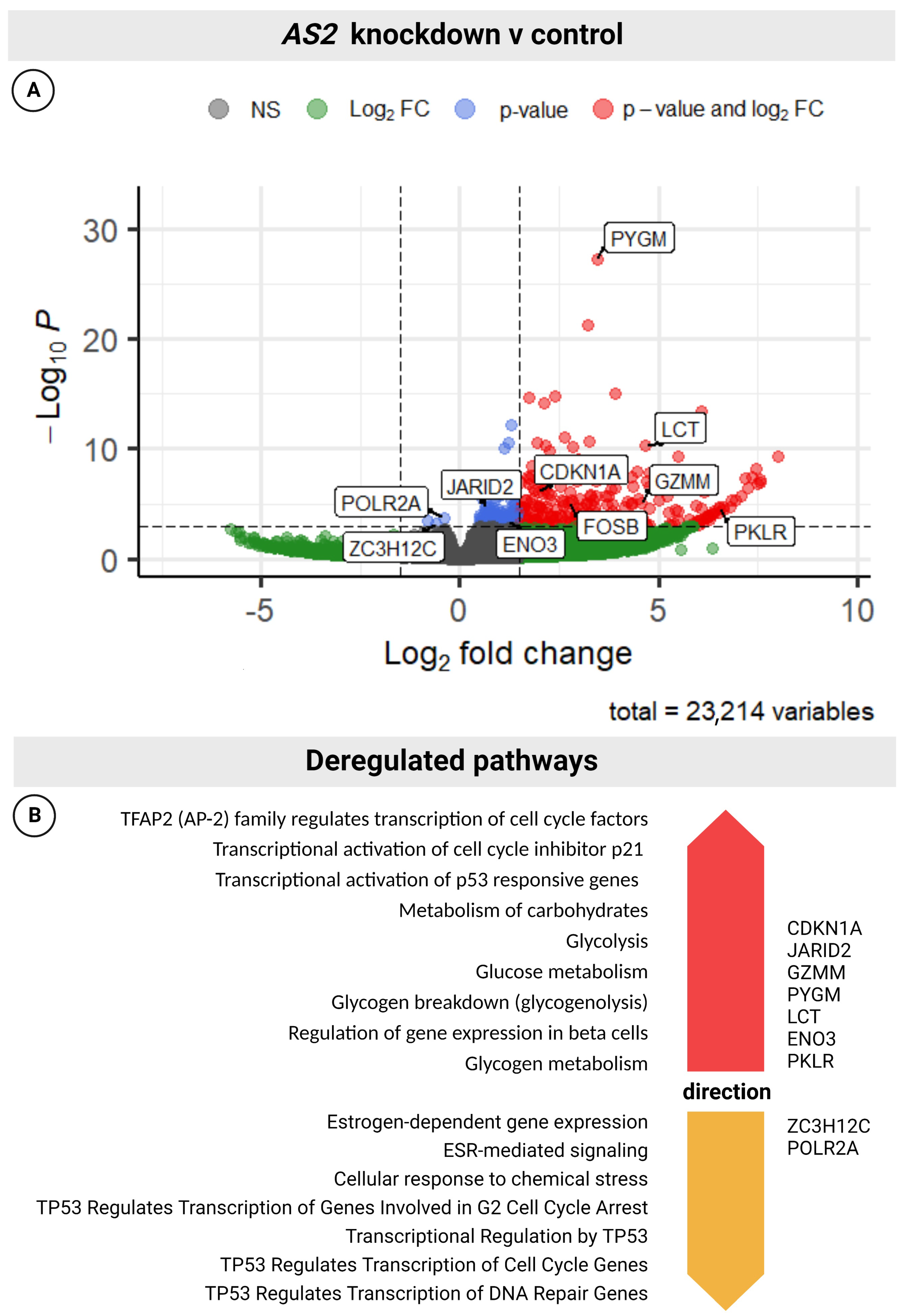
2.1.8. AS2 Is a Transcriptional Modulator
3. Discussion
4. Materials and Methods
4.1. Tumor lncRNA Sequencing Analysis
4.1.1. Clinical Samples
4.1.2. RNA Sequencing
4.1.3. RNA Sequencing Data Analysis
4.1.4. Cell Culture
4.1.5. Androgen Modulation Assays
4.1.6. Androgen Taper Assays
4.1.7. RNA Extraction
4.1.8. Reverse Transcription (RT) Reactions
4.1.9. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
4.1.10. Cytoplasmic and Nuclear RNA Isolation
4.1.11. RNA Fluorescent In Situ Hybridization (RNA-FISH) and Immunofluorescence
4.2. Transfection of GapmeR Antisense Oligonucleotides
4.2.1. Transfection of siRNAs
4.2.2. Cell Viability Assay
4.2.3. Apoptosis Assay
4.2.4. Cell Cycle Analysis
4.2.5. In Silico Analysis
4.2.6. Transcriptome Analysis
4.2.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Mansinho, A.; Macedo, D.; Fernandes, I.; Costa, L. Castration-Resistant Prostate Cancer: Mechanisms, Targets and Treatment. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2018; Volume 1126, pp. 117–133. [Google Scholar]
- Toren, P.J.; Gleave, M.E. Evolving Landscape and Novel Treatments in Metastatic Castrate-Resistant Prostate Cancer. Asian J. Androl. 2013, 15, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho, I.; Day, T.K.; Tilley, W.D.; Selth, L.A. Androgen Receptor Signaling in Castration-Resistant Prostate Cancer: A Lesson in Persistence. Endocr. Relat. Cancer 2016, 23, T179–T197. [Google Scholar] [CrossRef]
- Wen, S.; Niu, Y.; Lee, S.O.; Chang, C. Androgen Receptor (AR) Positive vs Negative Roles in Prostate Cancer Cell Deaths Including Apoptosis, Anoikis, Entosis, Necrosis and Autophagic Cell Death. Cancer Treat Rev. 2014, 40, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crona, D.J.; Whang, Y.E. Androgen Receptor-Dependent and-Independent Mechanisms Involved in Prostate Cancer Therapy Resistance. Cancers 2017, 9, 67. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Y.; Li, L.; Niu, Y.; Lin, W.; Lin, C.; Antonarakis, E.S.; Luo, J.; Yeh, S.; Chang, C. Preclinical Study Using Malat1 Small Interfering RNA or Androgen Receptor Splicing Variant 7 Degradation Enhancer ASC-J9® to Suppress Enzalutamide-Resistant Prostate Cancer Progression. Eur. Urol. 2017, 72, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, K.Y.; Liu, Q.; Cao, Q. Androgen Receptor-Related Non-Coding RNAs in Prostate Cancer. Front. Cell Dev. Biol. 2021, 9, 750. [Google Scholar] [CrossRef]
- You, Z.; Liu, C.; Wang, C.; Ling, Z.; Wang, Y.; Wang, Y.; Zhang, M.; Chen, S.; Xu, B.; Guan, H.; et al. LncRNA CCAT1 Promotes Prostate Cancer Cell Proliferation by Interacting with DDX5 and MIR-28-5P. Mol. Cancer Ther. 2019, 18, 2469–2479. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Ueda, T.; Yokoyama, A.; Fujihara, A.; Hongo, F.; Ukimura, O. PCA3 Controls Chromatin Organization and P53 Signal Activation by Regulating LAP2α-Lamin A Complexes. Cancer Gene Ther. 2021, 3, 358–368. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Li, D.; Ma, X.; Xu, K.; Ding, B.; Li, H.; Wang, Z.; Ouyang, W.; Long, G.; et al. Androgen Receptor Splice Variant 7 Predicts Shorter Response in Patients with Metastatic Hormone-Sensitive Prostate Cancer Receiving Androgen Deprivation Therapy. Eur. Urol. 2021, 79, 879–886. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bokhoven, A.; Varella-Garcia, M.; Korch, C.; Johannes, W.U.; Smith, E.E.; Miller, H.L.; Nordeen, S.K.; Miller, G.J.; Lucia, M.S. Molecular Characterization of Human Prostate Carcinoma Cell Lines. Prostate 2003, 57, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Guo, X.; Wang, M.; Qin, R. The Patterns of Antisense Long Non-Coding RNAs Regulating Corresponding Sense Genes in Human Cancers. J. Cancer 2021, 12, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.S.; Camacho, N.; Luxton, H.J.; Pye, H.; Finn, R.; Heavey, S.; Pitt, J.; Moore, C.M.; Whitaker, H.C. Genetic Alterations in the 3q26.31-32 Locus Confer an Aggressive Prostate Cancer Phenotype. Commun. Biol. 2020, 3, 440. [Google Scholar] [CrossRef]
- Haffner, M.C.; Bhamidipati, A.; Tsai, H.K.; Esopi, D.M.; Vaghasia, A.M.; Low, J.Y.; Patel, R.A.; Guner, G.; Pham, M.T.; Castagna, N.; et al. Phenotypic Characterization of Two Novel Cell Line Models of Castration-Resistant Prostate Cancer. Prostate 2021, 81, 1159–1171. [Google Scholar] [CrossRef]
- Ahmed Fouad, Y.; Aanei, C. Revisiting the Hallmarks of Cancer. Am. J. Cancer Res. 2017, 7, 1016–1036. [Google Scholar]
- Villegas, V.E.; Zaphiropoulos, P.G. Neighboring Gene Regulation by Antisense Long Non-Coding RNAs. Int. J. Mol. Sci. 2015, 16, 3251–3266. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome Pathway Analysis: A High-Performance in-Memory Approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [Green Version]
- Wadosky, K.M.; Koochekpour, S. Molecular Mechanisms Underlying Resistance to Androgen Deprivation Therapy in Prostate Cancer. Oncotarget 2016, 7, 64447. [Google Scholar] [CrossRef] [Green Version]
- Attard, G.; Murphy, L.; Clarke, N.W.; Cross, W.; Jones, R.J.; Parker, C.C.; Gillessen, S.; Cook, A.; Brawley, C.; Amos, C.L.; et al. Abiraterone Acetate and Prednisolone with or without Enzalutamide for High-Risk Non-Metastatic Prostate Cancer: A Meta-Analysis of Primary Results from Two Randomised Controlled Phase 3 Trials of the STAMPEDE Platform Protocol. Lancet 2022, 399, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Karsh, L.I.; Nissenblatt, M.J.; Canfield, S.E. Androgen Receptor Splice Variant, AR-V7, as a Biomarker of Resistance to Androgen Axis-Targeted Therapies in Advanced Prostate Cancer. Clin. Genitourin Cancer 2020, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, M.; Hirst, C.; Crawford, E.D. Characterising the Castration-Resistant Prostate Cancer Population: A Systematic Review. Int. J. Clin. Pract. 2011, 65, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Su, Z.; Liang, X.; Huang, Y.; Lan, Z.; Jiang, X. Long Non-Coding Rnas in Prostate Tumorigenesis and Therapy (Review). Mol. Clin. Oncol. 2020, 13, 76. [Google Scholar] [CrossRef]
- Osman, I.; Drobnjak, M.; Fazzari, M.; Ferrara, J.; Scher, H.I.; Cordon-Cardo, C. Inactivation of the P53 Pathway in Prostate Cancer: Impact on Tumor Progression. Clin. Cancer Res. 1999, 5, 2082–2088. [Google Scholar]
- Whitaker, H.C.; Shiong, L.L.; Kay, J.D.; Grönberg, H.; Warren, A.Y.; Seipel, A.; Wiklund, F.; Thomas, B.; Wiklund, P.; Miller, J.L.; et al. N-Acetyl-L-Aspartyl-L-Glutamate Peptidase-like 2 Is Overexpressed in Cancer and Promotes a pro-Migratory and pro-Metastatic Phenotype. Oncogene 2014, 33, 5274–5287. [Google Scholar] [CrossRef] [Green Version]
- Kreis, N.N.; Louwen, F.; Yuan, J. The Multifaceted P21 (Cip1/Waf1/CDKN1A) in Cell Differentiation, Migration and Cancer Therapy. Cancers 2019, 11, 1220. [Google Scholar] [CrossRef] [Green Version]
- Gartel, A.L. The Conflicting Roles of the Cdk Inhibitor P21(CIP1/WAF1) in Apoptosis. Leuk Res. 2005, 29, 1237–1238. [Google Scholar] [CrossRef]
- Pardo, J.; Bosque, A.; Brehm, R.; Wallich, R.; Naval, J.; Müllbacher, A.; Anel, A.; Simon, M.M. Apoptotic Pathways Are Selectively Activated by Granzyme A and/or Granzyme B in CTL-Mediated Target Cell Lysis. J. Cell Biol. 2004, 167, 457–468. [Google Scholar] [CrossRef]
- Gazon, H.; Barbeau, B.; Mesnard, J.M.; Peloponese, J.M. Hijacking of the AP-1 Signaling Pathway during Development of ATL. Front. Microbiol. 2018, 8, 2686. [Google Scholar] [CrossRef] [Green Version]
- Park, J.A.; Na, H.H.; Jin, H.O.; Kim, K.C. Increased Expression of Fosb through Reactive Oxygen Species Accumulation Functions as Pro-Apoptotic Protein in Piperlongumine Treated MCF7 Breast Cancer Cells. Mol. Cells 2019, 42, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Migocka-Patrzałek, M.; Elias, M. Muscle Glycogen Phosphorylase and Its Functional Partners in Health and Disease. Cells 2021, 10, 883. [Google Scholar] [CrossRef] [PubMed]
- Zois, C.E.; Harris, A.L. Glycogen Metabolism Has a Key Role in the Cancer Microenvironment and Provides New Targets for Cancer Therapy. J. Mol. Med. 2016, 94, 137–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnier, J.B.; Nishi, K.; Gumerlock, P.H.; Gorin, F.A.; Bradbury, E.M. Glycogen Synthesis Correlates with Androgen-Dependent Growth Arrest in Prostate Cancer. BMC Urol. 2005, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Lou, W.; Zhu, Y.; Nadiminty, N.; Schwartz, C.T.; Evans, C.P.; Gao, A.C. Niclosamide Inhibits Androgen Receptor Variants Expression and Overcomes Enzalutamide Resistance in Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2014, 20, 3198–3210. [Google Scholar] [CrossRef] [Green Version]
- Rehman, Y.; Rosenberg, J.E. Abiraterone Acetate: Oral Androgen Biosynthesis Inhibitor for Treatment of Castration-Resistant Prostate Cancer. Drug Des. Devel. Ther. 2012, 6, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Beltran, H.; Prandi, D.; Mosquera, J.M.; Benelli, M.; Puca, L.; Cyrta, J.; Marotz, C.; Giannopoulou, E.; Chakravarthi, B.V.S.K.; Varambally, S.; et al. Divergent Clonal Evolution of Castration-Resistant Neuroendocrine Prostate Cancer. Nat. Med. 2016, 22, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Benoist, G.E.; van Oort, I.M.; Boerrigter, E.; Verhaegh, G.W.; van Hooij, O.; Groen, L.; Smit, F.; de Mol, P.; Hamberg, P.; Dezentjé, V.O.; et al. Prognostic Value of Novel Liquid Biomarkers in Patients with Metastatic Castration-Resistant Prostate Cancer Treated with Enzalutamide: A Prospective Observational Study. Clin. Chem. 2020, 66, 842–851. [Google Scholar] [CrossRef]
- Boerrigter, E.; Benoist, G.E.; van Oort, I.M.; Verhaegh, G.W.; de Haan, A.F.J.; van Hooij, O.; Groen, L.; Smit, F.; Oving, I.M.; de Mol, P.; et al. RNA Biomarkers as a Response Measure for Survival in Patients with Metastatic Castration-Resistant Prostate Cancer. Cancers 2021, 13, 6279. [Google Scholar] [CrossRef]
- Wu, T.D.; Reeder, J.; Lawrence, M.; Becker, G.; Brauer, M.J. GMAP and GSNAP for Genomic Sequence Alignment: Enhancements to Speed, Accuracy, and Functionality. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1418, pp. 283–334. [Google Scholar]
- Ramsköld, D.; Wang, E.T.; Burge, C.B.; Sandberg, R. An Abundance of Ubiquitously Expressed Genes Revealed by Tissue Transcriptome Sequence Data. PLoS Comput. Biol. 2009, 5, e1000598. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification Strategies in Real-Time PCR; A-Z of Quantitative PCR: La Jolla, CA, USA, 2004. [Google Scholar]
- Bishop, E.; Engelman, J.M.; Mizuuchi, A.; Craigie, K.; Katzman, R.; Katz, M.; Skalka, R.A.; Leis, A.M.; Craigie, J.; Fujiwara, R.; et al. Evolution of Sex in RNA Viruses. J. Theor. Biol. 1989, 86, 99–112. [Google Scholar]
- Blighe, K.; Rana, S.; Lewis, M. Publication-Ready Volcano Plots with Enhanced Colouring and Labeling; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Sander, C.; Stuart, J.M.; Chang, K.; Creighton, C.J.; et al. The Cancer Genome Atlas Pan-Cancer Analysis Project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groen, L.; Yurevych, V.; Ramu, H.; Chen, J.; Steenge, L.; Boer, S.; Kuiper, R.; Smit, F.P.; Verhaegh, G.W.; Mehra, N.; et al. The Androgen Regulated lncRNA NAALADL2-AS2 Promotes Tumor Cell Survival in Prostate Cancer. Non-Coding RNA 2022, 8, 81. https://doi.org/10.3390/ncrna8060081
Groen L, Yurevych V, Ramu H, Chen J, Steenge L, Boer S, Kuiper R, Smit FP, Verhaegh GW, Mehra N, et al. The Androgen Regulated lncRNA NAALADL2-AS2 Promotes Tumor Cell Survival in Prostate Cancer. Non-Coding RNA. 2022; 8(6):81. https://doi.org/10.3390/ncrna8060081
Chicago/Turabian StyleGroen, Levi, Viktor Yurevych, Harshitha Ramu, Johnny Chen, Lianne Steenge, Sabrina Boer, Renske Kuiper, Frank P. Smit, Gerald W. Verhaegh, Niven Mehra, and et al. 2022. "The Androgen Regulated lncRNA NAALADL2-AS2 Promotes Tumor Cell Survival in Prostate Cancer" Non-Coding RNA 8, no. 6: 81. https://doi.org/10.3390/ncrna8060081