
Direct Comparison of Chol-siRNA Polyplexes and Chol-DsiRNA Polyplexes Targeting STAT3 in a Syngeneic Murine Model of TNBC


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Activities of 5′-Overlapping siSTAT3 and DsiSTAT3 Sequences in Murine Syngeneic 4T1 Breast Cancer Epithelial Cells
2.2. Hydrodynamic Diameters, Surface Charges, and Physical Stabilities of Chol-siRNA Polyplexes and Chol-DsiRNA Polyplexes
2.3. Activities of Chol-siSTAT3 Polyplexes and Chol-DsiSTAT3 Polyplexes in Murine Syngeneic 4T1 Breast Cancer Epithelial Cells
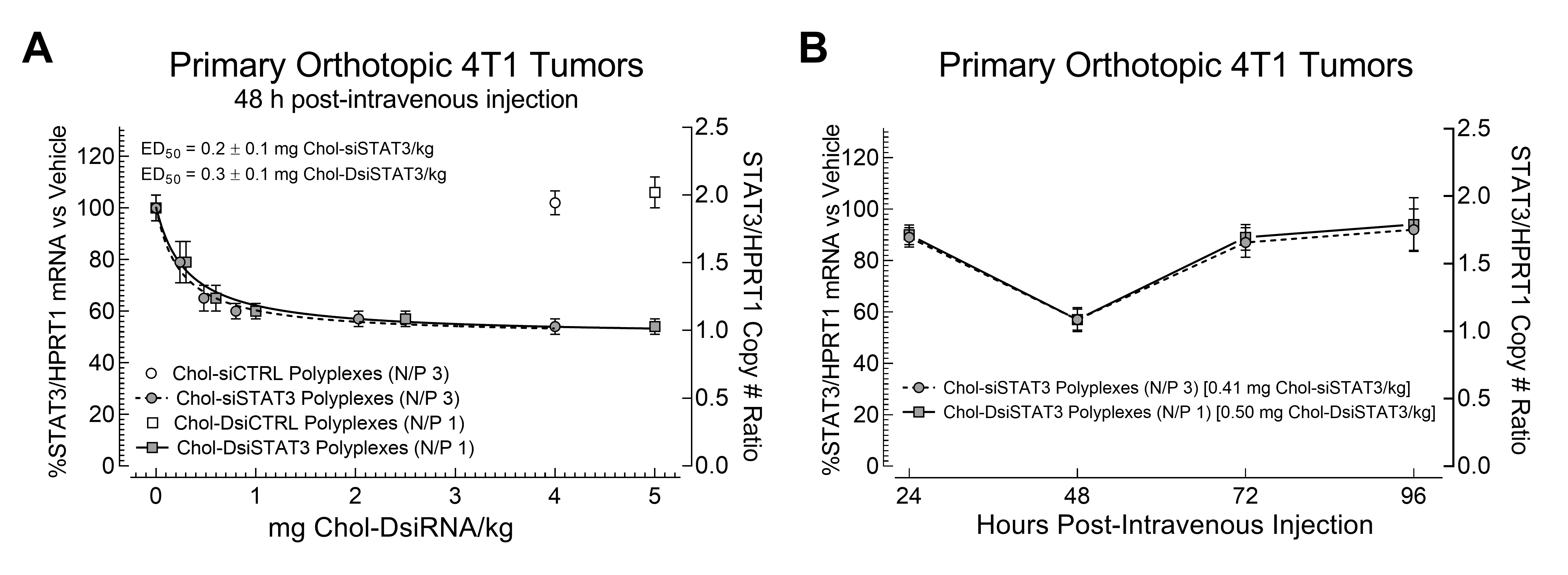
2.4. Potencies of Chol-siSTAT3 Polyplexes and Chol-DsiSTAT3 Polyplexes in Primary Murine Syngeneic Breast Tumors after IV Administration
2.5. Kinetics of mRNA Suppression in Primary Murine Syngeneic Breast Tumors by Chol-siSTAT3 Polyplexes and Chol-DsiSTAT3 Polyplexes after IV Administration
2.6. Chol-siSTAT3 Polyplex and Chol-DsiSTAT3 Polyplex Activities against the Growth of Primary Murine Syngeneic Breast Tumors after IV Administration
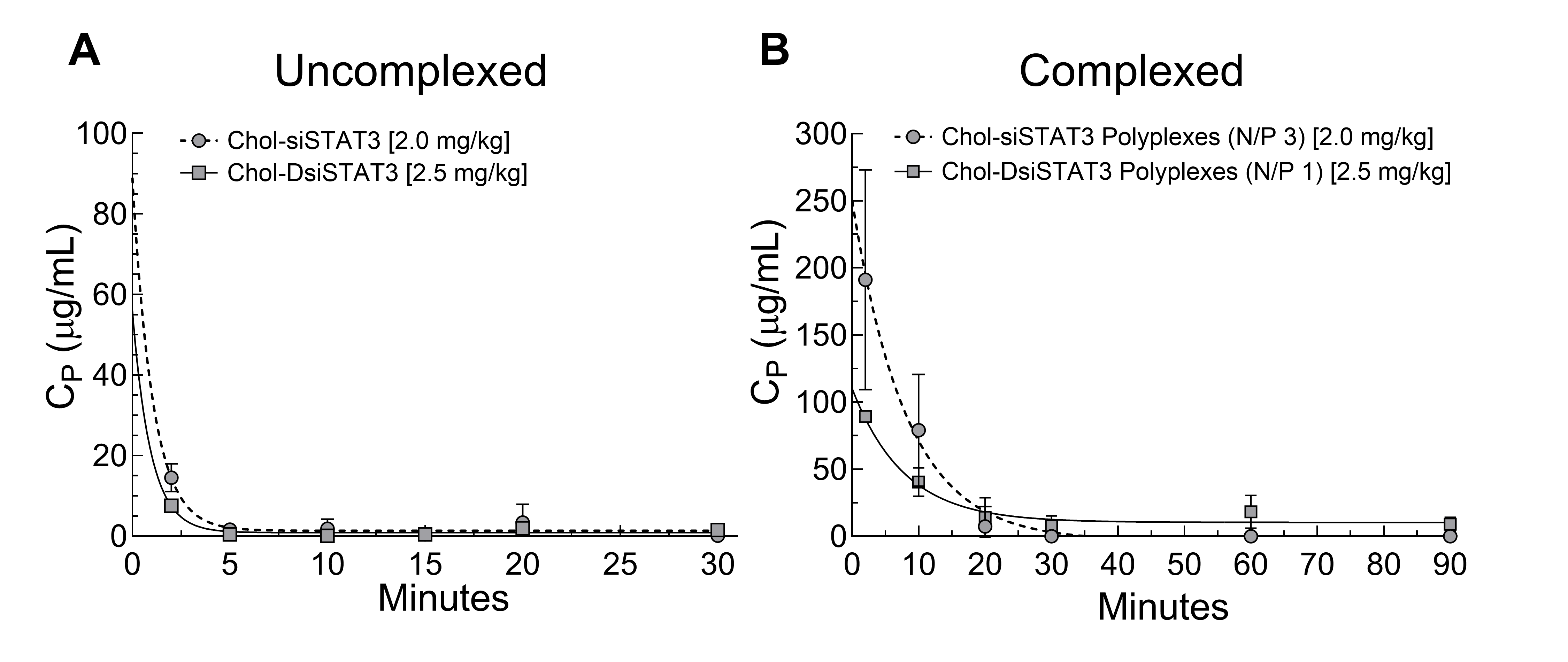
2.7. Pharmacokinetics of Chol-DsiSTAT3 and Chol-DsiSTAT3 Polyplexes in Healthy Female BALB/c Mice after IV Administration
2.8. Distribution of Complexed and Uncomplexed Chol-siSTAT3 and Chol-DsiSTAT3 in 4T1 Breast Tumor-Bearing Female BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Polymer
4.2. RNAi Molecules
4.3. Cell Culture
4.4. Electroporation of Murine 4T1 Cells
4.5. Formation of Chol-RNAi Polyplexes
4.6. Hydrodynamic Diameters and Zeta Potentials of Chol-RNAi Polyplexes
4.7. Endotoxin Levels
4.8. Transfection of Murine 4T1 Cells
4.9. Quantitation of Murine STAT3 mRNA, Chol-siSTAT3, and Chol-DsiSTAT3 by Reverse Transcription-Droplet Digital™ PCR (RT-ddPCR)
4.10. Potencies and Kinetics of STAT3 mRNA Suppression in Primary Murine 4T1 Breast Tumors by Chol-RNAi Polyplexes after IV Administration
4.11. Tumor Growth Delay of Primary Murine 4T1 Breast Tumors
4.12. Quantitation of Stat3 Protein in Primary 4T1 Tumors by Western Blot
4.13. Pharmacokinetics of Chol-RNAi after i.v. Administration
4.14. Distribution of Chol-RNAi Molecules in Tumors and Organs after i.v. Administration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caillaud, M.; El Madani, M.; Massaad-Massade, L. Small interfering RNA from the lab discovery to patients’ recovery. J. Control. Release Off. J. Control. Release Soc. 2020, 321, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Abdelmoaty, M.M.; Ambardekar, V.V.; Curran, S.M.; Dyavar, S.R.; Arnold, L.L.; Cohen, S.M.; Kumar, D.; Alnouti, Y.; Coulter, D.W.; et al. Preliminary preclinical study of Chol-DsiRNA polyplexes formed with PLL[30]-PEG[5K] for the RNAi-based therapy of breast cancer. Nanomedicine 2021, 33, 102363. [Google Scholar] [CrossRef] [PubMed]
- Hager, S.; Fittler, F.J.; Wagner, E.; Bros, M. Nucleic Acid-Based Approaches for Tumor Therapy. Cells 2020, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- Ambardekar, V.V.; Wakaskar, R.R.; Sharma, B.; Bowman, J.; Vayaboury, W.; Singh, R.K.; Vetro, J.A. The efficacy of nuclease-resistant Chol-siRNA in primary breast tumors following complexation with PLL-PEG(5K). Biomaterials 2013, 34, 4839–4848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grijalvo, S.; Alagia, A.; Jorge, A.F.; Eritja, R. Covalent Strategies for Targeting Messenger and Non-Coding RNAs: An Updated Review on siRNA, miRNA and antimiR Conjugates. Genes 2018, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Mangala, L.S.; Rodriguez-Aguayo, C.; Kong, X.; Lopez-Berestein, G.; Sood, A.K. RNA interference-based therapy and its delivery systems. Cancer Metastasis Rev. 2018, 37, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Dagnaes-Hansen, F.; Nielsen, E.J.; Wengel, J.; Besenbacher, F.; Howard, K.A.; Kjems, J. The effect of chemical modification and nanoparticle formulation on stability and biodistribution of siRNA in mice. Mol. Ther. 2009, 17, 1225–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambardekar, V.V.; Wakaskar, R.R.; Ye, Z.; Curran, S.M.; McGuire, T.R.; Coulter, D.W.; Singh, R.K.; Vetro, J.A. Complexation of Chol-DsiRNA in place of Chol-siRNA greatly increases the duration of mRNA suppression by polyplexes of PLL(30)-PEG(5K) in primary murine syngeneic breast tumors after i.v. administration. Int. J. Pharm. 2018, 543, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Manaia, E.B.; Abucafy, M.P.; Chiari-Andreo, B.G.; Silva, B.L.; Oshiro Junior, J.A.; Chiavacci, L.A. Physicochemical characterization of drug nanocarriers. Int. J. Nanomed. 2017, 12, 4991–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, S.; Sousa, J.; Pais, A.; Vitorino, C. Nanomedicine: Principles, Properties, and Regulatory Issues. Front. Chem. 2018, 6, 360. [Google Scholar] [CrossRef] [PubMed]
- Schrors, B.; Boegel, S.; Albrecht, C.; Bukur, T.; Bukur, V.; Holtstrater, C.; Ritzel, C.; Manninen, K.; Tadmor, A.D.; Vormehr, M.; et al. Multi-Omics Characterization of the 4T1 Murine Mammary Gland Tumor Model. Front. Oncol. 2020, 10, 1195. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Fang, M.; Alroy, J.; Sahagian, G.G. Imagable 4T1 model for the study of late stage breast cancer. BMC Cancer 2008, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, O.M.; Takabe, K. Animal models for exploring the pharmacokinetics of breast cancer therapies. Expert Opin. Drug Metab. Toxicol. 2015, 11, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, X.; Arlinghaus, R.B. Knockdown of STAT3 expression by RNA interference inhibits the induction of breast tumors in immunocompetent mice. Cancer Res. 2005, 65, 2532–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.; Chand, A.; Gough, D.; Ernst, M. Therapeutically exploiting STAT3 activity in cancer—using tissue repair as a road map. Nat. Rev. Cancer 2018, 19, 82–96. [Google Scholar] [CrossRef]
- Dai, L.; Cheng, L.; Zhang, X.; Jiang, Q.; Zhang, S.; Wang, S.; Li, Y.; Chen, X.; Du, T.; Yang, Y.; et al. Plasmid-based STAT3-siRNA efficiently inhibits breast tumor growth and metastasis in mice. Neoplasma 2011, 58, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Hather, G.; Liu, R.; Bandi, S.; Mettetal, J.; Manfredi, M.; Shyu, W.C.; Donelan, J.; Chakravarty, A. Growth rate analysis and efficient experimental design for tumor xenograft studies. Cancer Inf. 2014, 13, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyplex | Endotoxin 1 EU/mg (±SD) | Drug Loading Wt% Chol-RNAi | Diameter 2 nm (±SD) | Zeta Potential 3 mV (±SD) |
|---|---|---|---|---|
| Chol-siCTRL Polyplexes (N/P 3) | 0.122 (0.003) | 25 | 25 (2) | 8 (2) |
| Chol-DsiCTRL Polyplexes (N/P 1) | 0.023 (0.002) | 50 | 33 (2) | 5.2 (0.7) |
| PK Parameter | Chol-siSTAT3 (±SD) | Chol-siSTAT3 Polyplexes (±SD) | Chol-DsiSTAT3 (±SD) | Chol-DsiSTAT3 Polyplexes (±SD) |
|---|---|---|---|---|
| C0 (μg/mL) | 89 (65) | 250 (123) | 56 (64) | 110 (7) |
| AUC0–∞ (h·μg/mL) | 2.3 (0.9) | 44 (2) | 3 (1) | 39 (13) |
| MRT0–∞ (h) | 0.07 (0.03) | 0.5 (0.5) | 0.8 (0.9) | 0.9 (0.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Z.; Abdelmoaty, M.M.; Curran, S.M.; Dyavar, S.R.; Kumar, D.; Alnouti, Y.; Coulter, D.W.; Podany, A.T.; Singh, R.K.; Vetro, J.A. Direct Comparison of Chol-siRNA Polyplexes and Chol-DsiRNA Polyplexes Targeting STAT3 in a Syngeneic Murine Model of TNBC. Non-Coding RNA 2022, 8, 8. https://doi.org/10.3390/ncrna8010008
Ye Z, Abdelmoaty MM, Curran SM, Dyavar SR, Kumar D, Alnouti Y, Coulter DW, Podany AT, Singh RK, Vetro JA. Direct Comparison of Chol-siRNA Polyplexes and Chol-DsiRNA Polyplexes Targeting STAT3 in a Syngeneic Murine Model of TNBC. Non-Coding RNA. 2022; 8(1):8. https://doi.org/10.3390/ncrna8010008
Chicago/Turabian StyleYe, Zhen, Mai Mohamed Abdelmoaty, Stephen M. Curran, Shetty Ravi Dyavar, Devendra Kumar, Yazen Alnouti, Don W. Coulter, Anthony T. Podany, Rakesh K. Singh, and Joseph A. Vetro. 2022. "Direct Comparison of Chol-siRNA Polyplexes and Chol-DsiRNA Polyplexes Targeting STAT3 in a Syngeneic Murine Model of TNBC" Non-Coding RNA 8, no. 1: 8. https://doi.org/10.3390/ncrna8010008