3.1. Taxonomy
3.1.1. Dothideomycetes, Patellariales, Patellariaceae
MycoBank: MB842041.
Diagnosis: Ascomata discoidal to ovoid, black, 1–1.5 × 600–800 µm; asci hyaline, pedicellate, bitunicate, 40–45 × 12–16 µm; ascospores ellipsoid to fusoid, golden brown, transverse septae, uniseriate to irregular biseriate, 32–36 × 10–12 µm.
Type: MEXICO: Hidalgo, Zacualtipán de Ángeles municipality, Bosque El Hayal, sobre la desviación a Tlahuelompa, 20°37′34′’ N, 98°37′07′’ W, 2250 m, 2 July 2013, T. Raymundo 4608 (ENCB).
GenBank: ITS OQ877252, nrLSU OQ880481, SSU OQ878242.
Etymology: The epithet indicates that the species grows in Hidalgo.
Ascomata 1–1.5 mm diameter, 600–800 µm thick, solitary to gregarious, erumpent to superficial, rounded to angular, discoidal to powdery, 1 to 1.5 mm diameter, sessile, rough, exposing the asci, black color; basal peridium green to black, 160-200 µm thick; paraphysoids 3–3.5 µm in diameter, filiform, branched, anastomosed, deep green; paraphyses protruding from asci; asci 40–45 × 12–16 µm, bitunicate, cylindrical to clavate, sessile, rounded at the apex, octosporate; ascospores uniseriate to irregularly biseriate in the apical part (28.8–) 32–36 (40) × (9.6–) 10–12 (–12.8) µm broadly ellipsoid to fusiform, bicellular, with one septum and constricted in the middle part, the distal portion slightly larger than the proximal, pale yellowish to dark golden brown when ripe.
Habitat: Gregarious on decaying branches of angiosperms.
Additional specimens examined: MEXICO: Hidalgo, Zacualtipán de Ángeles municipality, Bosque El Hayal, sobre la desviación a Tlahuelompa, 20°37′34″ N, 98°37′07″ W, 2250 m, 2 July 2013, R. Valenzuela 14997 (ENCB, Paratype).
Taxonomical notes: This species is characterized by pulvinated ascomata, black and erumpent, ascospores 32–36 × 10–12 µm, bicellular, with a golden-brown color and a germinative pore. Morphologically and phylogenetically, it is close to
Holmiella sabina (De Not.) Petrini, Samuels & E. Müll. However, the former presents ascomata with toothed margins, ascospores of 25–40 × 13–20 µm, bicellular, reddish-brown with two germinative pores.
Holmiella juniperi-semiglobosae Pem, Gafforov, Jeewon & K.D. Hyde and
H. junipericola Pem, Gafforov, Jeewon & K.D. Hyde are species that are phylogenetically related; however, they grow on
Juniperus semiglobosa and
J. zerawschanica, respectively, from Uzbekistan [
17].
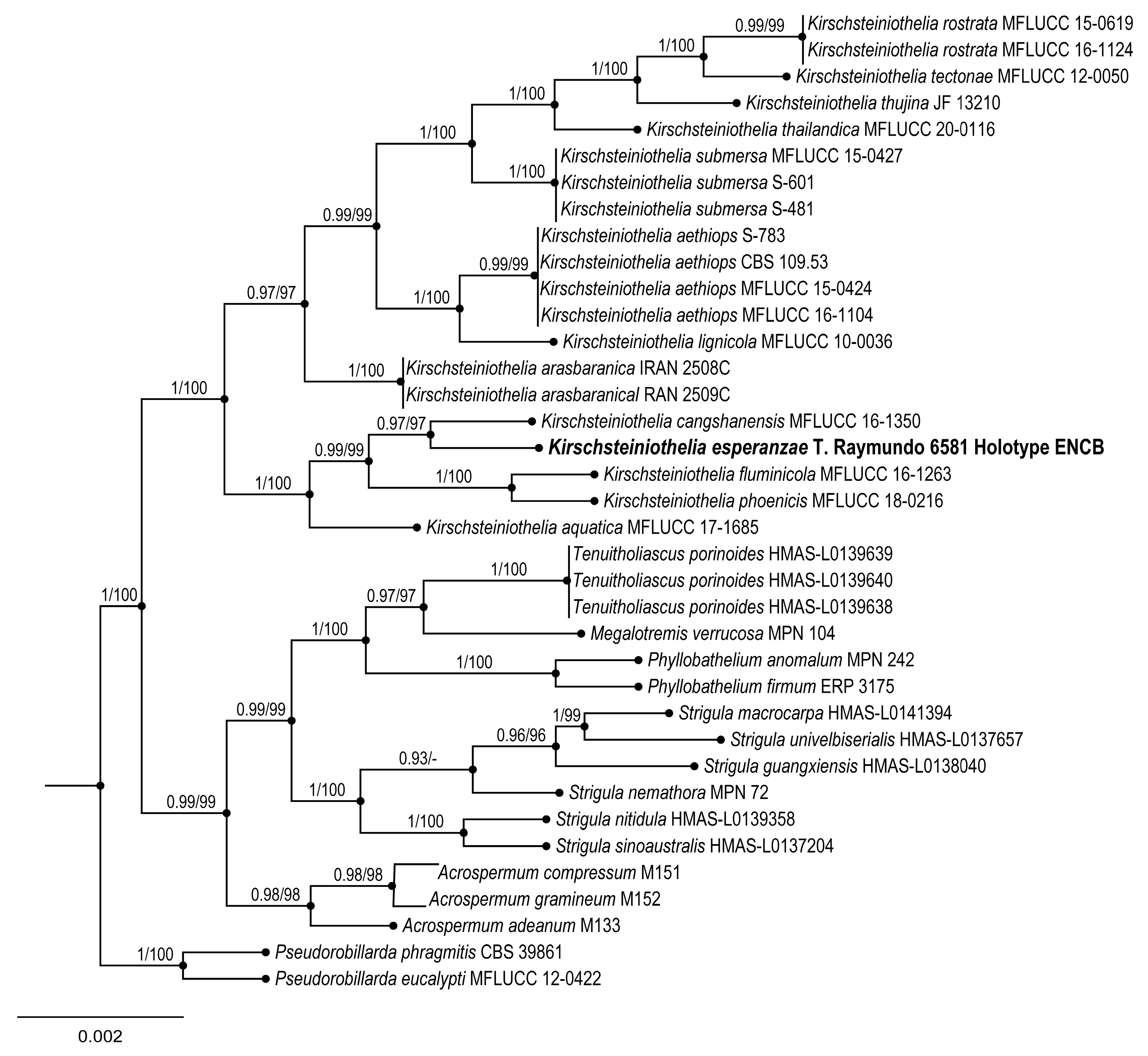
3.1.2. Pleosporales, Kirschteiniotheliaceae
- 2.
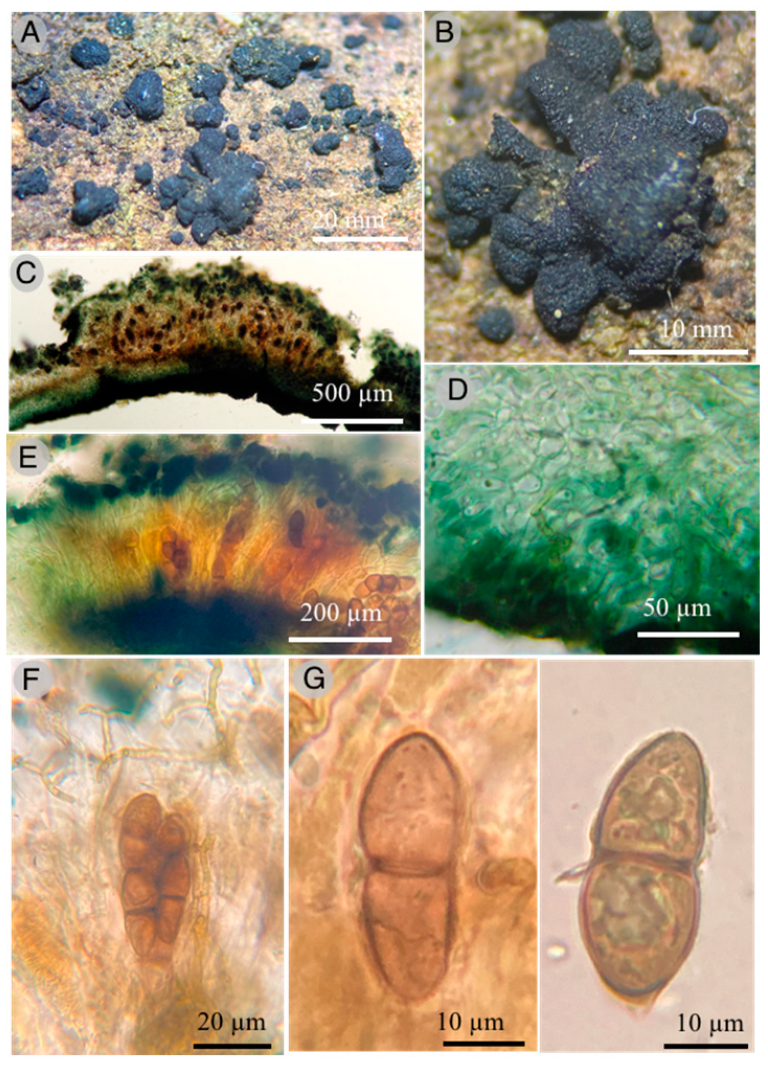
Kirschsteiniothelia esperanzae Raymundo, Cobos-Villagrán & R. Valenz. sp. nov.
MycoBank: MB822042.
Diagnosis: Ascomata 300–700 µm diameter × 600–700 µm high, peridium 55 to 100 μm thick, pseudoperiphyses 5 μm wide, asci (168–)178–203 × 32–35 μm and ascospores 40–50(–53) × 14–17 μm.
Type: MEXICO: Oaxaca, Sierra de Juárez, Ixtlán district, Santiago Comaltepec, La Esperanza, Carretera Oaxaca-Tuxtepec km 51, 17°37′55″ N, 96°22′01″ W, 1600 m, 21 May 2017; T. Raymundo 6581 (ENCB, Holotype).
GenBank: ITS: OQ877253, LSU: OQ880482.
Etymology: The epithet refers to La Esperanza’s locality, where the species was collected.
Pseudothecia 400–700 µm diameter × 600–700 µm high, hemispherical to globose-subglobose, generally aggregated, seldom dispersed, completely superficial when mature, black, with a well-defined brown ostiole; peridium 55–100 µm thick, tapering at the base, 55 µm, laterally to 60 µm and broader at the base of the ostiole, up to 100 µm wide, pseudoparenchymatous, composed of isoradiated cells of (10–) 16–20 × (11–) 12–15 µm, prismatic texture, slightly thick walls, 1–1.5 µm; pseudoparaphysis very thick up to 5 µm wide, hyaline, branched and anastomosed; asci (168–) 178–203 × 32–35 µm, bitunicate, fisitunicate, fusiform to soleiform, with internal apical beak, eight spores; ascospores 40–50 (–53) × 14–17 µm, ellipsoid or soleiform, 1-septate, slightly constricted at the septum, light brown to olive-brown, smooth.
Habitat: Gregarious on decaying wood.
Taxonomical notes: This species is characterized by having larger asci (168–)178–203 × 32–35 µm and ascospores 40–50(–53) × 14–17 µm. This species is morphologically similar to
Kirschsteiniothelia thujina (Peck) D. Hawksw. due to the long ascomata (300–600 µm) and ascospores. However,
K. thujina has a dark reddish-brown ostiole and an angular texture in the peridium, and the host is
Abies balsamea Mill. and
Thuja occidentalis L.
Kirschsteiniothelia esperanzae has a brown ostiole and peridium cells with prismatic texture, and the host is not identified. This last species was collected in the
Oreomunnea mexicana (Standl.) J.-F.Leroy TMCF of Oaxaca. Phylogenetic data confirm that
K. esperanzae is a new species, close to
K. thujina and
K. rostrata Jing Yang & K.D. Hyde. These two species and
K.
arasbaranica Mehrabi, R. Hemmati & Asgari form a large clade. These three species have the largest ascospores of the group, more than 30 × 15 µm [
35,
36].
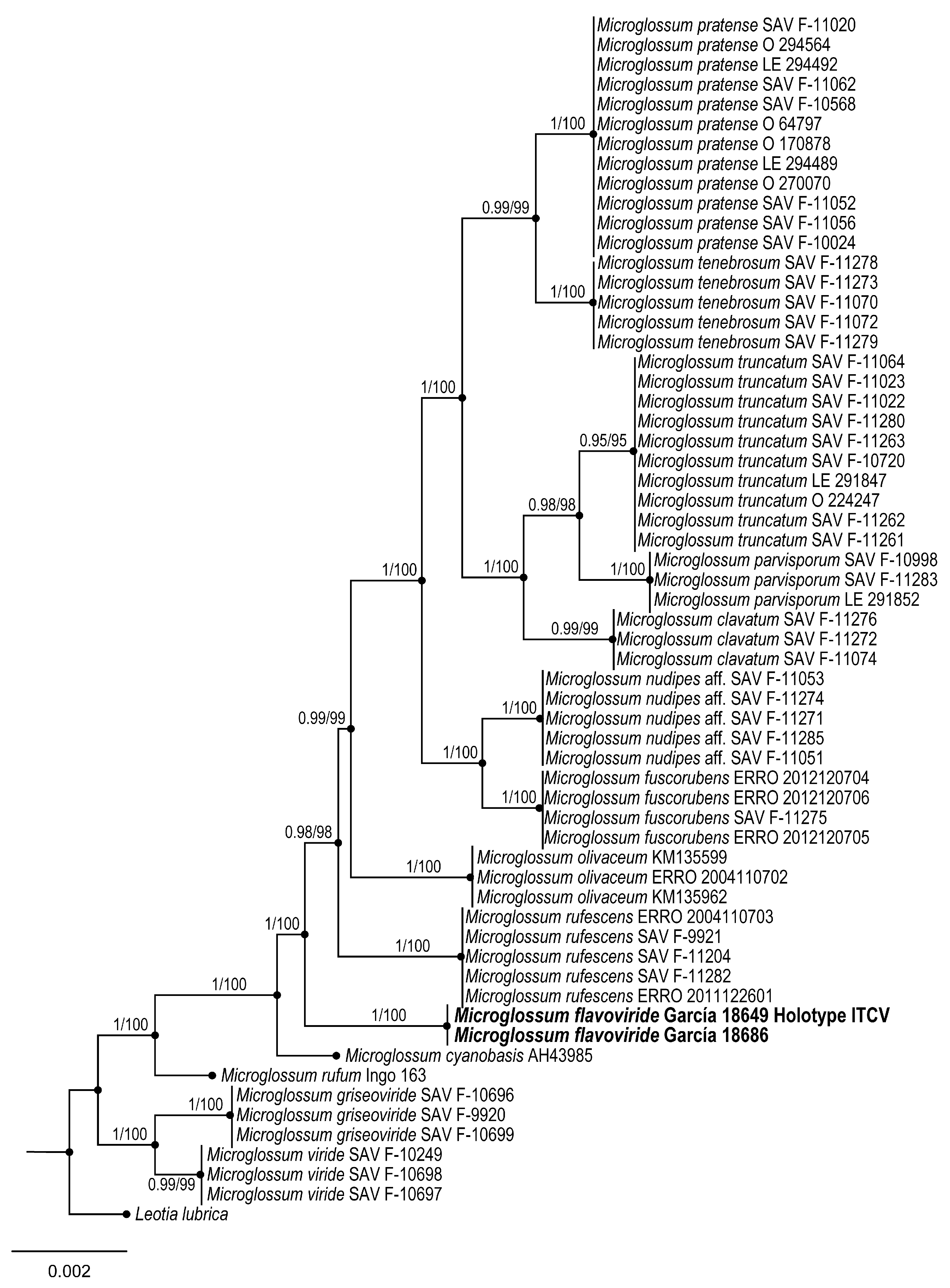
3.1.3. Geoglossomycetes, Geoglossales, Geoglossaceae
- 3.
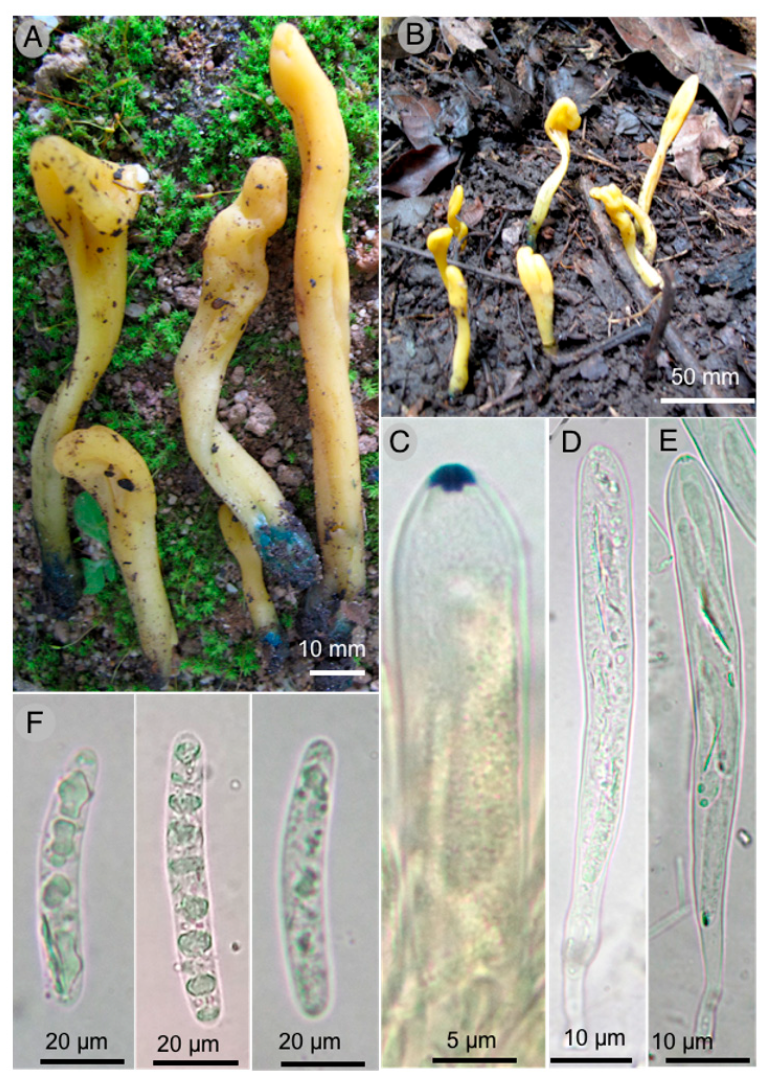
Microglossum flavoviride Sánchez-Flores, García-Jiménez & Raymundo sp. nov.
MycoBank: MB842043.
Diagnosis: Ascomata 16–65 × 4–9 mm, gregarious, lanceolate to spatulate, yellowish-green and deep green at the base, asci 111–160 × 11–14 µm, octosporate, hyaline, ascospores (20–) 22–45 × 4–6 (–6.5) µm, bacilliform, cylindrical, with 6–10 septa.
Type: MEXICO: Chiapas, Ocozocoautla municipality, Laguna Bélgica, 16°52′44.12″ N, 93°27′25.64″ W, 1004 m, 16 August 2011, J. García 18649 (ITCV, Holotype).
GenBank: ITS: OQ877254, LSU: OQ880483.
Etymology: It was named flavoviride for the ascoma color.
Ascomata 16–65 mm long, gregarious, lanceolate to spatulate, yellowish-green (30A7) color, deep green (27E8) at the base, cartilaginous consistency, viscous-moist texture; stipe 14–36 mm long, 2 mm wide toward the apex and 4–9 mm wide toward the base, flattened laterally, hollow, turns green when cut, fertile part 10–30 × 4–9 mm; medullar excipulus with intricate texture, formed by hyphae 3–8 µm in diameter, hyaline, indistinguishable subhymen; hymenium 160–185 µm thick; paraphyses 2–4 (–5) µm diameter, filiform, hyaline, septate, bifurcate toward the base, blunt apex, nodulous, irregular to rounded, hook-shaped to straight; asci 111–160 × 11–14 µm, octosporate, hyaline, clavate, amyloid operculum; ascospores (20–) 22–45 × 4–6 (–6.5) µm, bacilliform, cylindrical, slightly allantoic to spindle-shaped, hyaline, multigutulate, 6–10 septa not very visible.
Additional specimens: MEXICO. Chiapas, Ocozocoautla municipality, Laguna Bélgica, 16°52′44.12″ N, 93°27′25.64″ W, 1004 m, 16 August 2011, J. García 18686 (ITCV).
Taxonomical notes: Ascomata 16–65 mm long, yellowish-green and deep green at the base, ascospores (20–) 22–45 × 4–6 (–6.5) µm, bacilliform, cylindrical, slightly allantoic to spindle-shaped, hyaline, 6-10 septa. It can be confused with
M. rufum (Schwein.) Underw. due to the color of the ascomata; however, this species presents granulations along the stipe and lacks the green tones at the base, with ascospores of similar size, although slightly smaller (18–) 20–36 (–40) × 4–6 µm, as well as smaller asci 100–135 × 9–12 µm [
37]. It can be separated from
M. fumosum (Peck) E.J. Durand by the size of the ascospores; the spores of the latter species are broader (16–) 20–40 (–48) × 4–5 µm, and the ascomata are pale yellow, cinnamon brown to reddish ocher [
37]. It is distinguished from
M. longisporum E.J. Durand by its cinnamon brown ascomata and larger ascospores 40–90 (–100) × 4–6 µm. Macroscopically, it resembles
M. cyanobasis P. Iglesias & Arauzo due to the green color at the base of the ascoma; however, ascomata are brown and not yellow as in
M. flavoviride, where the ascospores are smaller, 15.4–22.5 × 4.4–6.1 µm, and paraphyses present different forms [
38]. Likewise, it has similar shades at the base to
M. viride (Schrad. ex J.F. Gmel.) Gillet; however, ascospores of the latter species differ in size and shape, measuring (11–) 18–22 (–25) × (4–) 5–7 µm, and are elliptical to oblong, sometimes curved, and without visible septa [
39].
3.1.4. Leotiomycetes, Helotiales, Helotiaceae
- 4.
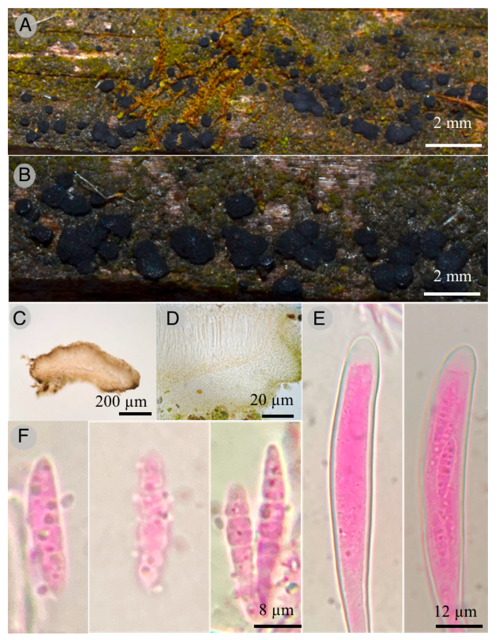
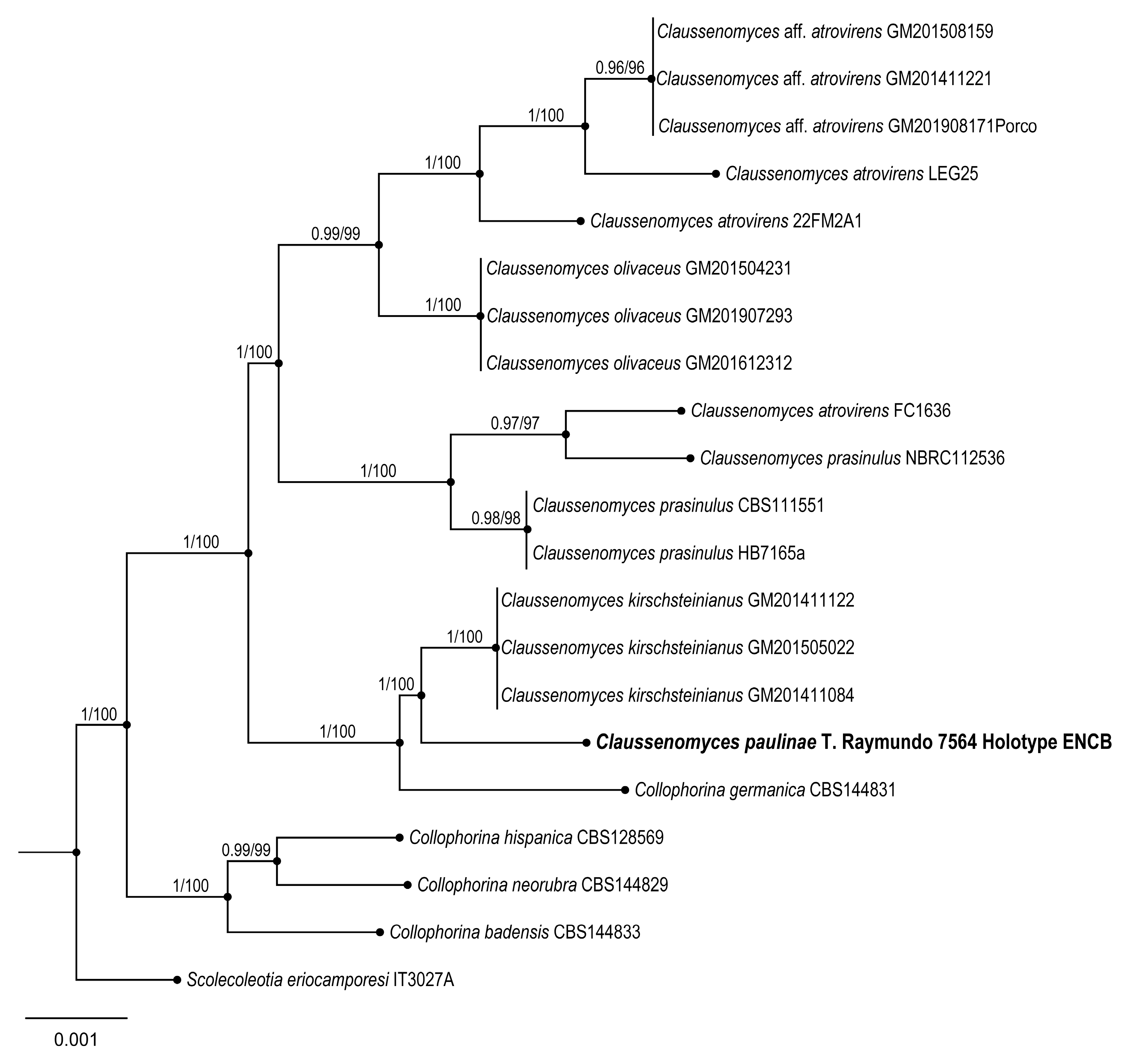
Claussenomyces paulinae Raymundo
MycoBank: MB842044
Diagnosis: Apothecia 600–800 μm diameter, discoid to flat, pulvinate, dark, gelatinous consistency; asci 85–120 × 8–10 μm, claviform, septum simple at the base, second basal cell presents crosier; ascospores 18–22 × 3–3.5 μm, fusoid, with three septa and smooth wall, curved, hyaline, some germinating and forming conidia 4 × 2 μm.
Type: MEXICO. Hidalgo: Zacualtipán de Ángeles municipality, El Hayal forest, 20°37′41.6″ N, 98°36′58.4″ W, 2000 m, 30 May 2018, T. Raymundo 7577 (ENCB).
GenBank: ITS: OQ877256.
Etymology: Dedicated to Rosa Paulina Calvillo Medina for her contributions to Mexican mycology.
Diagnosis: Apothecia 600–800 μm in diameter and 600–800 μm in height, flat pulvinate to discoid, bright black color, substipitate, with gelatinous consistency, slightly verrucose texture; ectal excipulum epidermoid to globular, with cells 14–20 μm in diameter, hyaline to pale yellow and green at the margin, thin and smooth walls. Intricate medullar excipulum with swollen hyphae 2 μm in diameter, tapering toward the margin; hymenium hyaline 110 μm thick, filiform paraphyses with capitate apices; asci 85–120 × 8–10 μm, claviform with blunt apices and simple septum at the base; second basal cell presents crosier, biseriate apically when young and uniseriate when mature, obliquely located, octosporic, hyaline; ascospores 18–22 × 3–3.5 μm, fusoid with seven septa and smooth walls, curved, hyaline, some germinating and forming conidia 4 × 2 μm, ovoid, hyaline.
Habitat: Saprotrophic species found on decaying wood of Pinus patula Schiede ex Schltdl. & Cham.
Additional specimens: MEXICO: Hidalgo, Zacualtipán de Ángeles municipality, El Hayal forest, 20°37′41.6″ N, 98°36′58.4″ W, 2000 m, 30 May 2018, R. Valenzuela 18282 (ENCB).
Taxonomical notes: This species has dark gregarious apothecia with jelly consistency, inamyloid asci, and ascospore fragments form secondary spores. Morphologically, it is similar to
C. atrovirens (Pers.) Korf & Abawi, which differs by forming dark green apothecia and ascospores with 4–7 septate [
40,
41]. Another similar species is
C. prassinulus (P. Karst.) Korf & Abawi, which has emerald green apothecia with ascospores 13–14 × 3–3.5 μm [
42,
43]. Phylogenetically,
C. paulinae is confirmed as an independent lineage forming an independent branch.
3.1.5. Pezizomycetes, Pezizales, Chorioactidaceae
- 5.
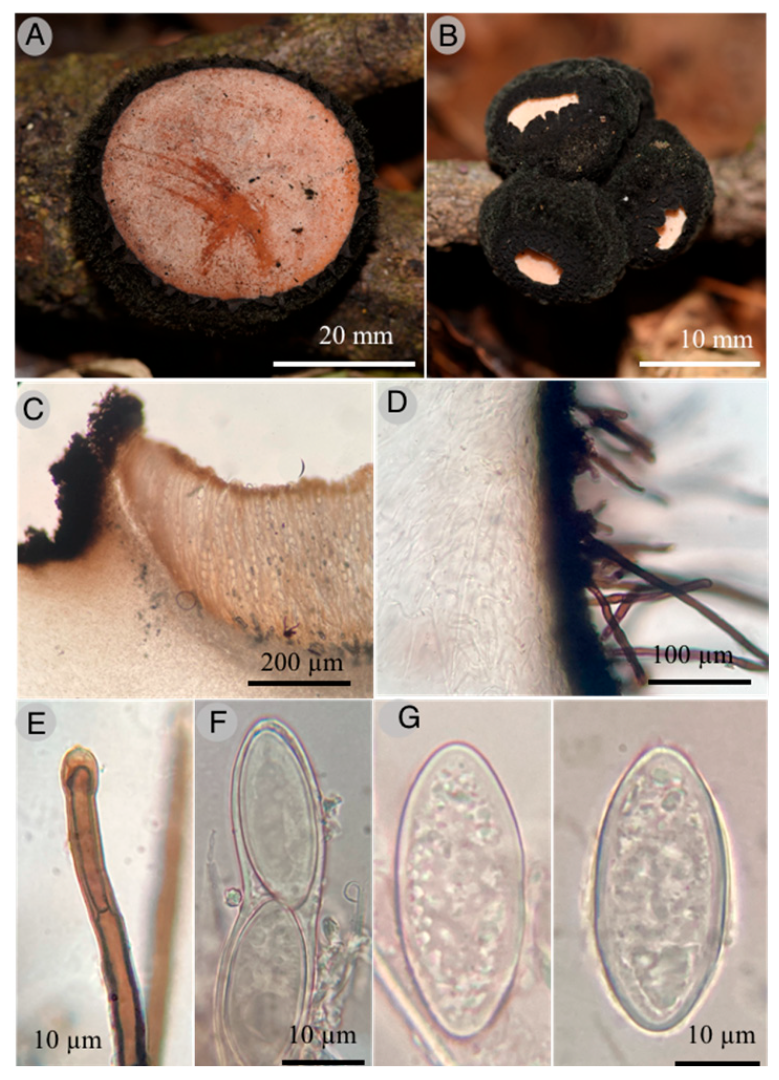
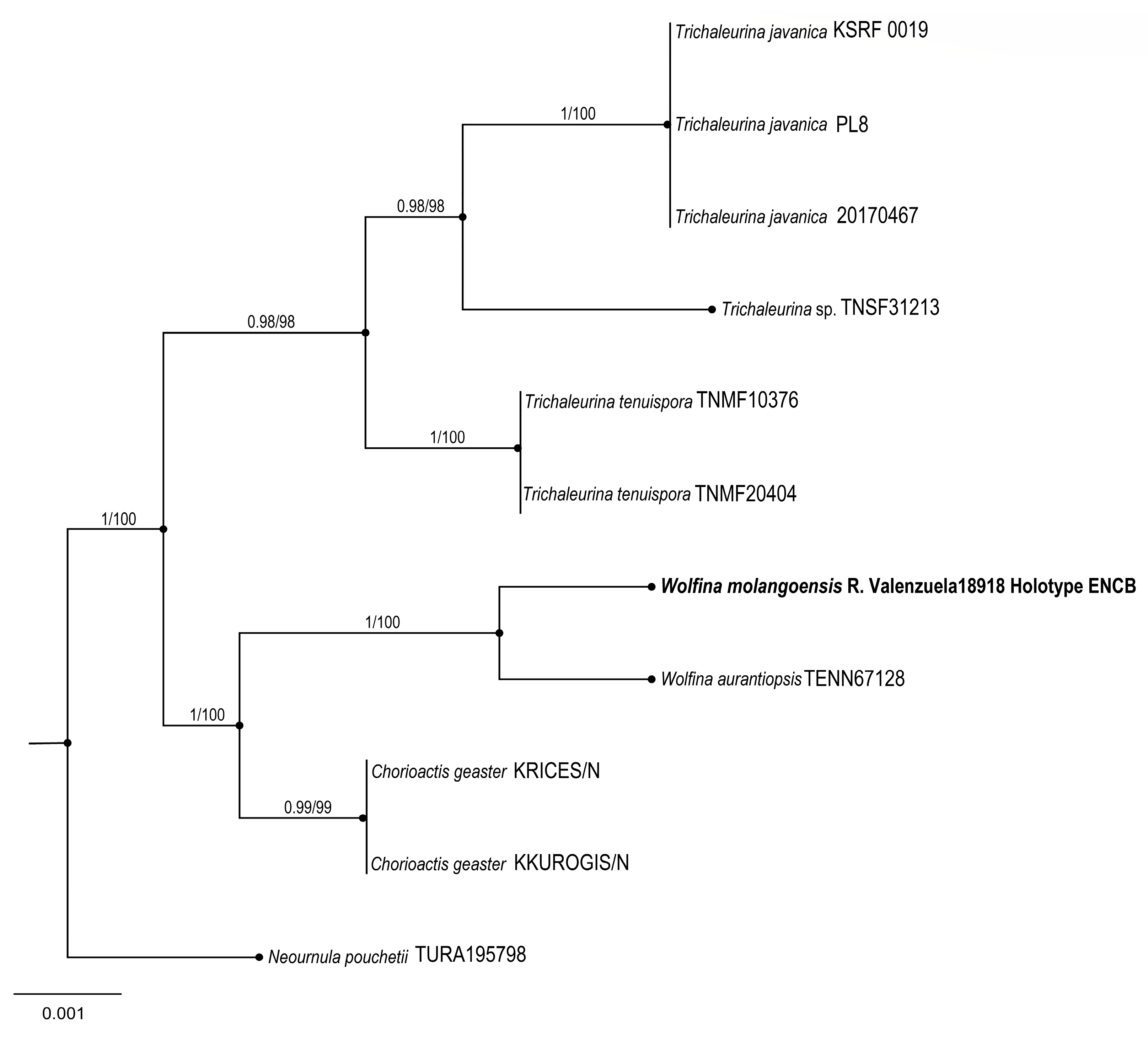
Wolfina molangoensis R. Valenz. & Raymundo
MycoBank: MB842045
Diagnosis: Apothecia 20–60 mm diameter, cup-shaped to discoid, external surface black, external hairs velvety; asci 400–450 × 22–24 μm, cylindrical, operculate; ascospores 35–40 × 14–18 μm, elliptical to cylindrical, hyaline, with granular content, smooth, thick-walled.
Type: MEXICO: Hidalgo. Molango municipality, Laguna de Atezca, 20°48′32″ N, 98°44′52″ W, alt. 1281 m, 31 May 2018, R. Valenzuela 18918 (ENCB, Holotype).
GenBank: ITS: OQ877257
Etymology: The name refers to the Molango locality in Hidalgo state.
Apothecia cup-shaped to discoid, sessile, 20–60 mm in diameter; hymenium shallow, pale orange (6A5) to peach (7A4), external surface black, velvety, convoluted, flesh thick, firm, corky when dried; external hairs cylindrical, 4–8 μm diameter, septate, walls up to 1 μm thick, brown, entire, smooth with apex lanceolate; ectal excipulum with pseudoparenchymatous texture, epidermoid cells with thick wall, dark brown; medullar excipulum intricate texture, hyphae hyaline, simple septate, 2–4 μm, wide hyphae; subhymenium of thick texture intricata, septate, 3–4 μm wide hyphae, arranged perpendicular to the asci; paraphyses 3–4 μm diameter, filiform, septate, anastomosing; asci 400–450 × 22–24 μm, cylindrical, operculate, with acute apex, walls up to 2 μm thick, octosporic, hyaline and inamyloid, tapering base and flexuous; ascospores 35–40 × 14–18 μm, elliptical to cylindrical, hyaline with granular content, sharp ends, thick-walled and smooth.
Habitat: Grows on branches of angiosperms.
Additional specimens examined: MEXICO: Hidalgo, Laguna de Atezca, 20°48′32″ N, 98°44′52″ W, 1281 m, 1 June 2018, T. Raymundo 7640 (ENCB, Paratype).
Taxonomical notes: Morphological and phylogenetically, this new species is close to
W. aurantiopsis (Ellis) Seaver ex Eckblad; however,
W. aurantiopsis forms apothecia 25–45 mm with yellow to ochraceous hymenium and ascospores 25–32 × 10–15 μm, elliptical to cylindrical, hyaline with granular content, rounded ends with thin-walled and striate. Argnello et al. [
26] noted that it might be restricted to the eastern USA. We found differences in the size and form of spores between species.
3.1.6. Sordariomycetes, Xylariales, Xylariaceae
- 6.
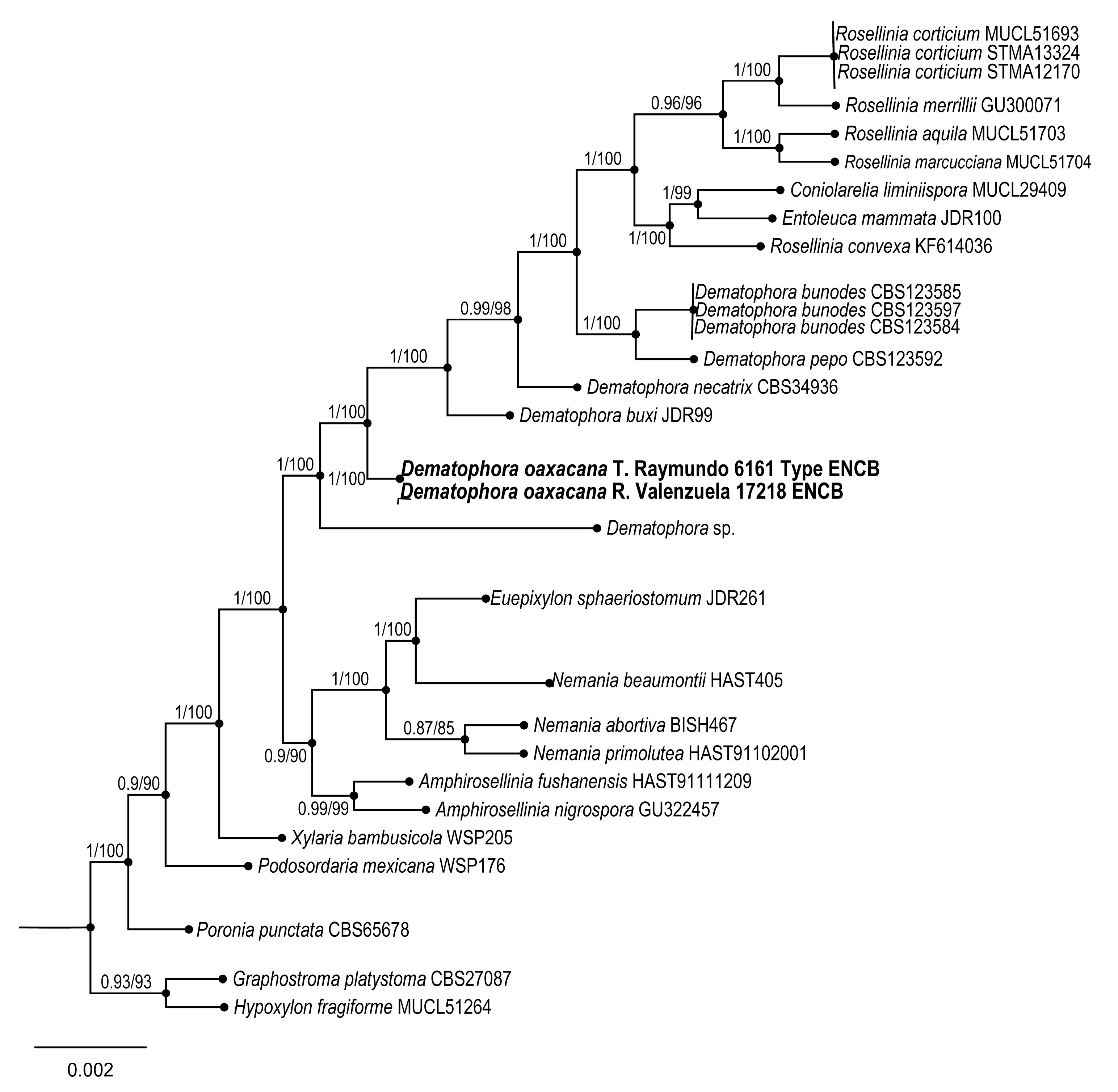
Dematophora oaxacana Sánchez-Flores, R. Valenz. & Raymundo sp. nov.
MycoBank: MB842051.
Diagnosis: Stromata 500–1100 × 400–700 µm, globose to subglobose, dark, solitary to gregarious, subiculum irregular extension, evanescent, carbonaceous, ascospores 20–29 × 10–13 (–14) µm, ovoid to asymmetrically ellipsoidal, with two cellular appendages, without germ slit.
Type: MEXICO: Oaxaca, Ixtlán de Juárez district, Santiago Comaltepec municipality, km 79 road Tuxtepec-Oaxaca, La Esperanza, Chinantla, 17°37′45″ N, 96°31′33″ W, 1130 m, 22 May 2017, T. Raymundo 6161 (ENCB, Holotype).
GenBank: ITS: OQ877258; nrLSU:OQ889487.
Etymology: The name refers to the state of Oaxaca, where this species was found.
Diagnosis: Stromata 500–1100 × 400–700 µm, globose to subglobose, dark to dark-brown, solitary, gregarious to cespitose, ostioles finely papillate to punctate; lack of subiculum; ectostroma dark, carbonaceous; endostroma 13–17 µm thick, light orange (5A4); perithecia not collapsed; asci dehiscent in 5% KOH; ascospores 20–29 × 10–13 (–14) µm, ovoid to asymmetrically ellipsoidal, brown, without germ slit, with flat sides ends, two cellular appendages, dehiscent in 5% KOH; external cellular appendage 3–5 µm tall and 5–6 µm wide, subglobose, hyaline; internal cellular appendage 1–2 µm tall and 2–3 µm wide, conical to subglobose, hyaline.
Habitat: Gregarious growing on decaying wood.
Distribution: Only known to be found in the state of Oaxaca.
Additional specimens examined: MEXICO, Oaxaca, Ixtlán de Juárez district, Santiago Comaltepec municipality, El Relámpago, La Esperanza, 17°35′28.1″ N, 96°53′52.2″ W, 1399 m, 29 May 2016, T. Raymundo 6161 (ENCB) and 6164 (ENCB). km 79 road Tuxtepec-Oaxaca, La Esperanza, Chinantla, 17°97′45″ N, 96°31′33″ W, 1130 m, 22 May 2017; 15 May 2015, R. Valenzuela 16111 (ENCB), 16145 (ENCB), T. Raymundo 5710 (ENCB). Loc. cit., 29 May 2016, R. Valenzuela 16667 (ENCB). Loc. cit., 22 May 2017, R. Valenzuela 17218 (ENCB), 17225 (ENCB), 17231 (ENCB), 17243 (ENCB), T. Raymundo 6587 (ENCB). Loc. cit., 23 May 2017, B. Nuñez 4 (ENCB), T. Raymundo 6607 (ENCB). Loc. cit., 30 April 2018, A. Cobos-Villagrán 1134 (ENCB). Paraje San Bernardo, La Esperanza, 17°37′55.4″ N, 96°22′1.5″ W, 25 September 2016, 1600 m, A. Trejo-Arana 17 (ENCB). Villa Alta district, Santiago Camotlán municipality, 5 km of Santiago Camotlán to San Juan Yatzuna, 24 March 2017, T. Raymundo (ENCB). Road Río Blanco, 25 March 2013, Galicia-Ávila 58 (ENCB). Santiago Camotlán, 25 March 2013, Escudero-Leyva 160 (ENCB).
Taxonomical notes: Ascospores measure 20–29.6 × 9.6–12 µm, without germline and double cell appendage. Phylogenetically, this species is close to
Dematophora buxi (Fabre) C. Lamb., Wittstein & M. Stadler, differing from the latter in its macro and microscopic characteristics, as a more persistent subicula, with narrower ascospores 19.8–30.1 × 6–8.9 µm, fusoid, with straight germline and rounded apices. It is also similar to
D. francisiae (L.E. Petrini) C. Lamb., Wittstein & M. Stadler; however, the latter has a persistent and felted subicula, 29–35 × 8–13 µm, longer ascospores, with a straight germline and rounded apices. Some species of
Dematophora were earlier considered under the genus
Rosellinia [
44].
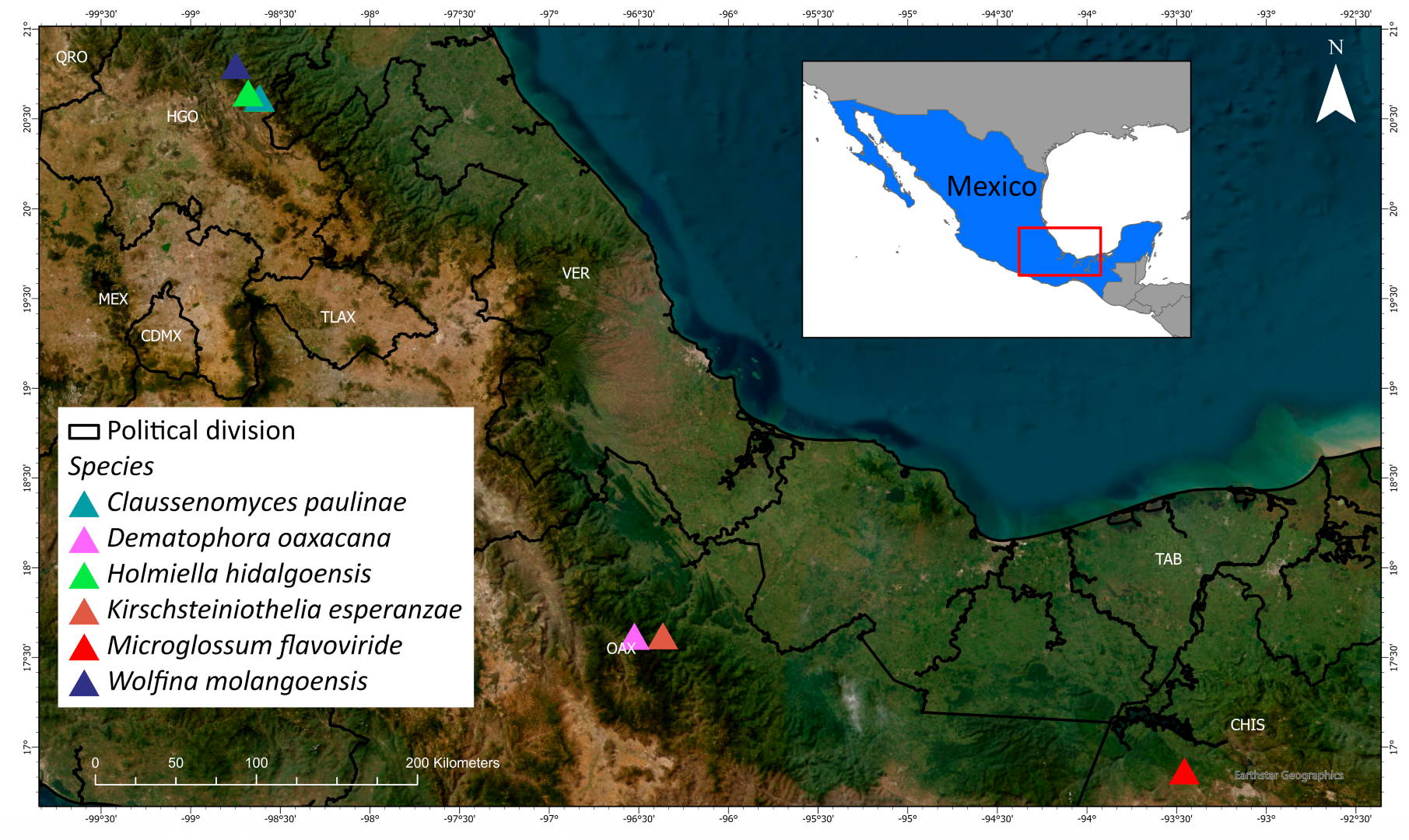
The distribution of the described new species is shown in
Figure 13.

 ,
, 
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}