
Electrical Conductivity and pH Are Two of the Main Factors Influencing the Composition of Arbuscular Mycorrhizal Fungal Communities in the Vegetation Succession Series of Songnen Saline-Alkali Grassland
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description and Sample Collection
2.2. Soil Physical and Chemical Properties and Enzyme Activities
2.3. DNA Extraction and PCR Amplification
2.4. Illumina Miseq Sequencing
2.5. Spore Morphological Identification
2.6. Determination of Root Colonization of Plants in the Wild and Establishment of Colonization System
2.7. Data Analysis and Processing
3. Results
3.1. Spore Morphological Identification of AMF
3.2. Analysis of Soil Physical and Chemical Properties
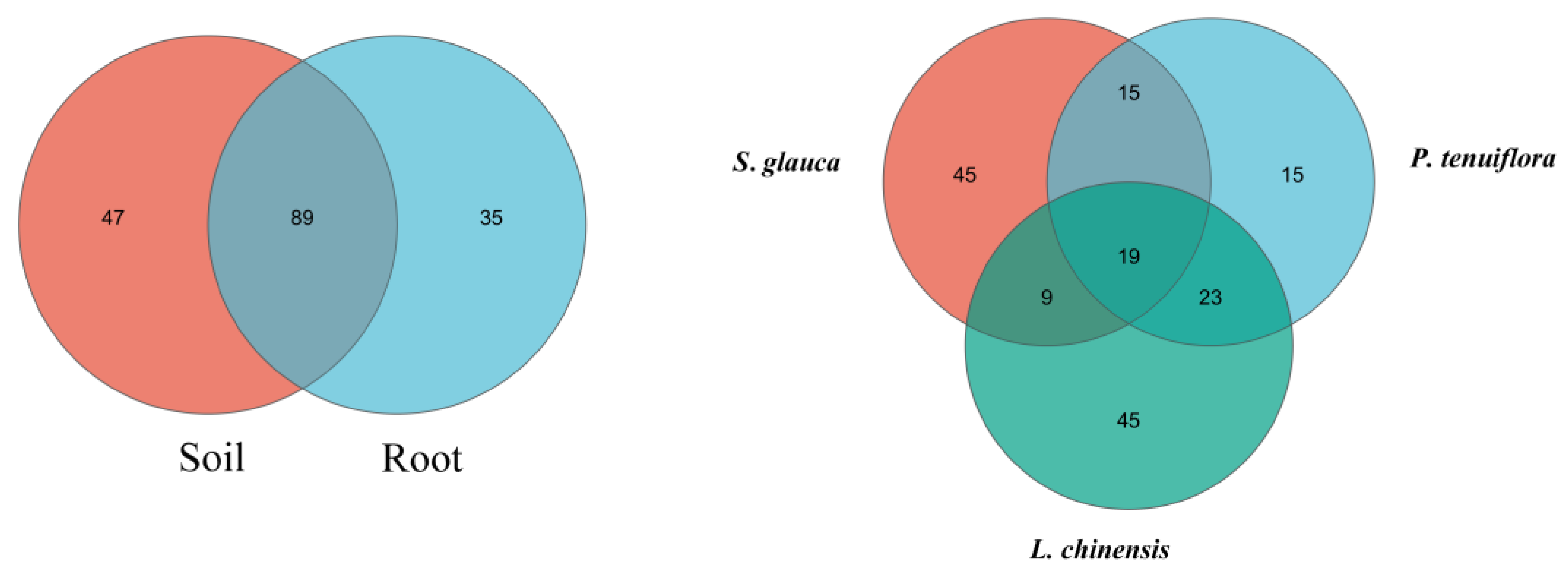
3.3. Changes in the Amount and Richness of AMF OUT
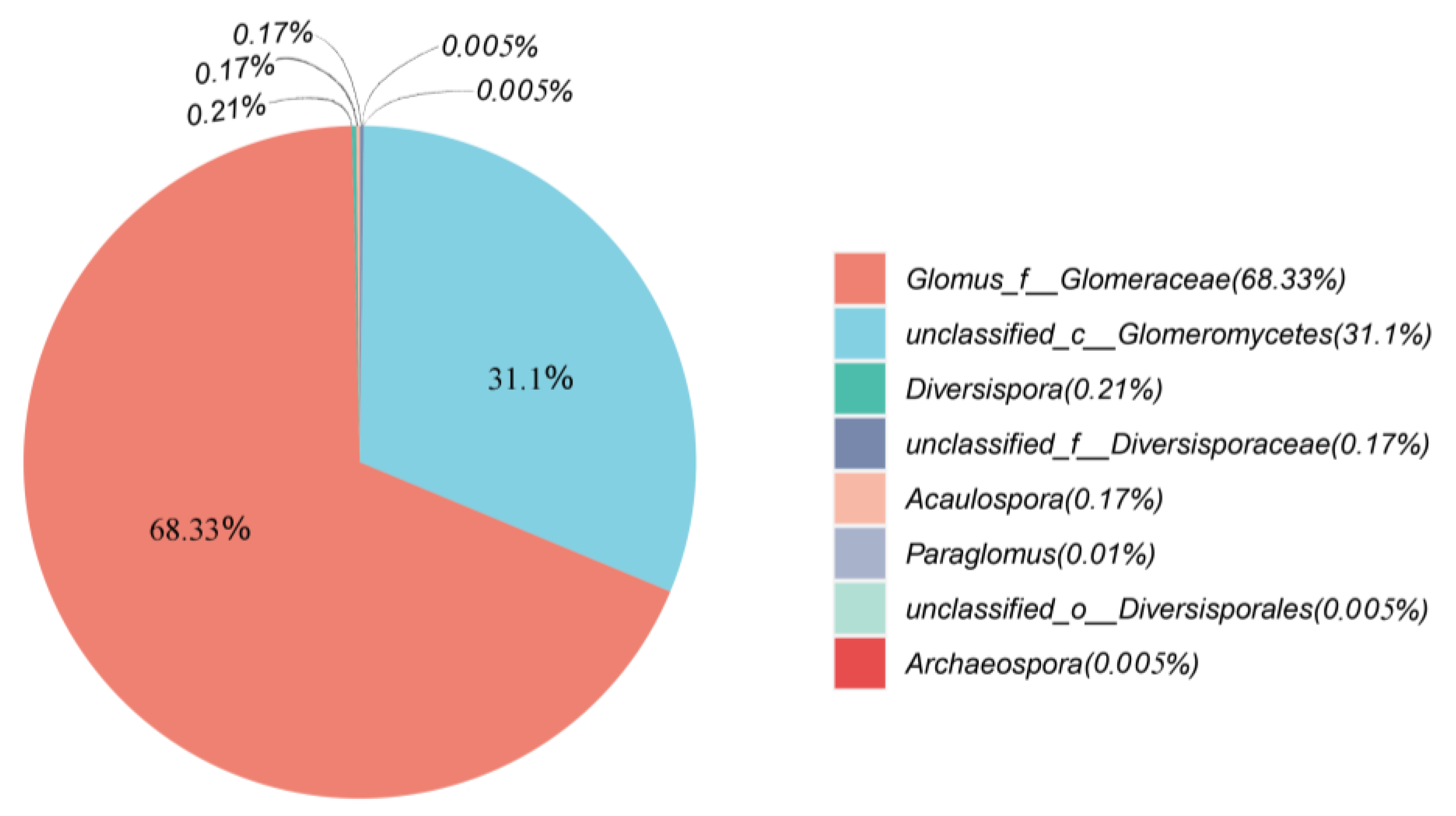
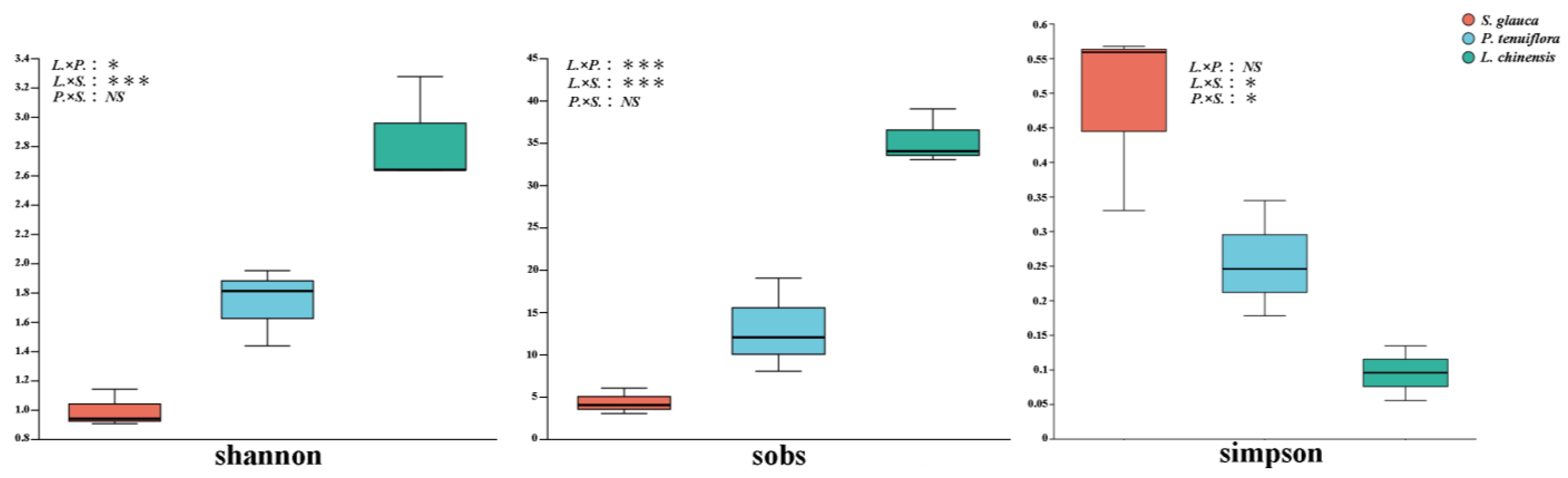
3.4. Changes in AMF Diversity and Community Structure of Plant Community Succession Sequence
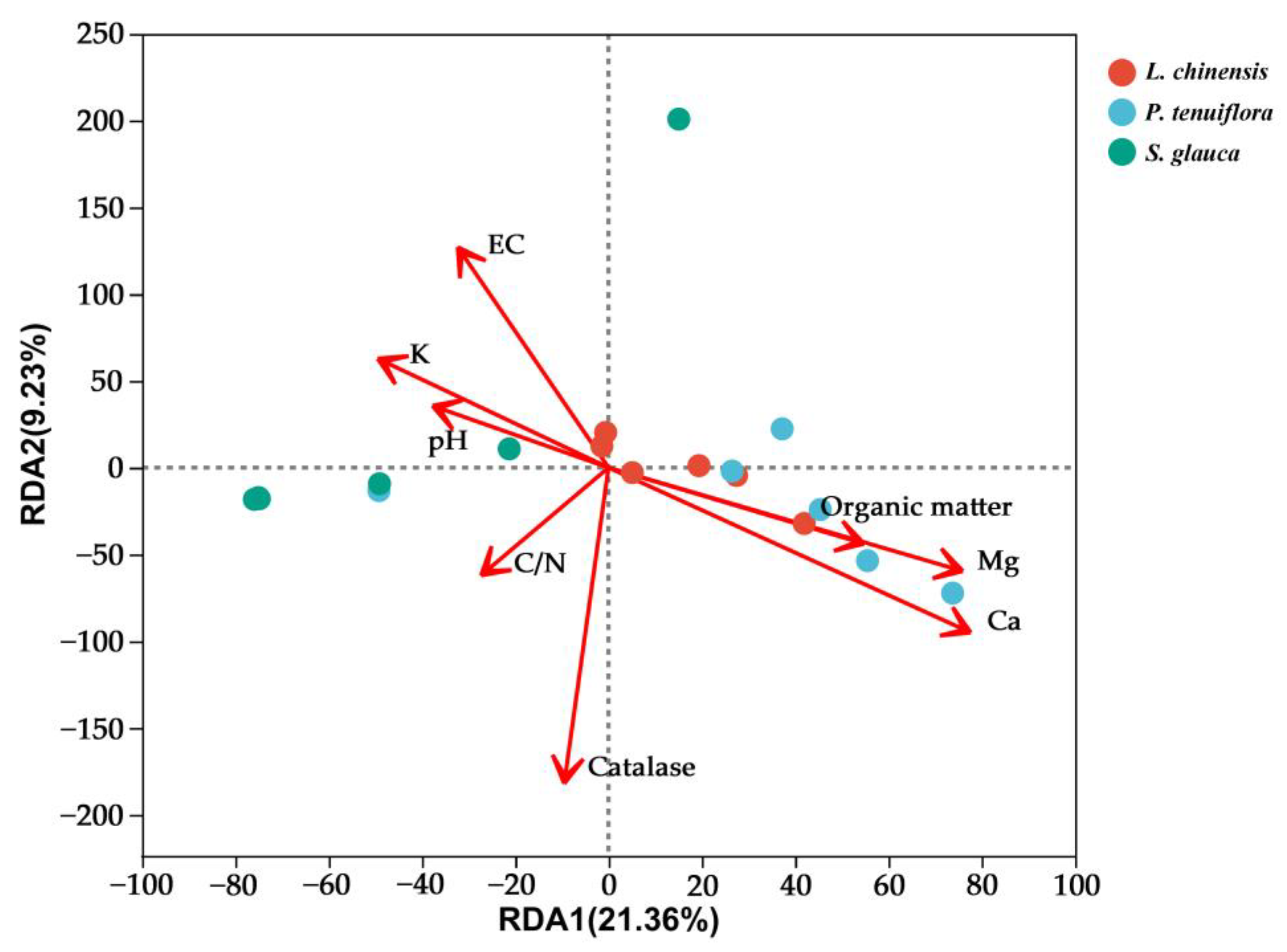
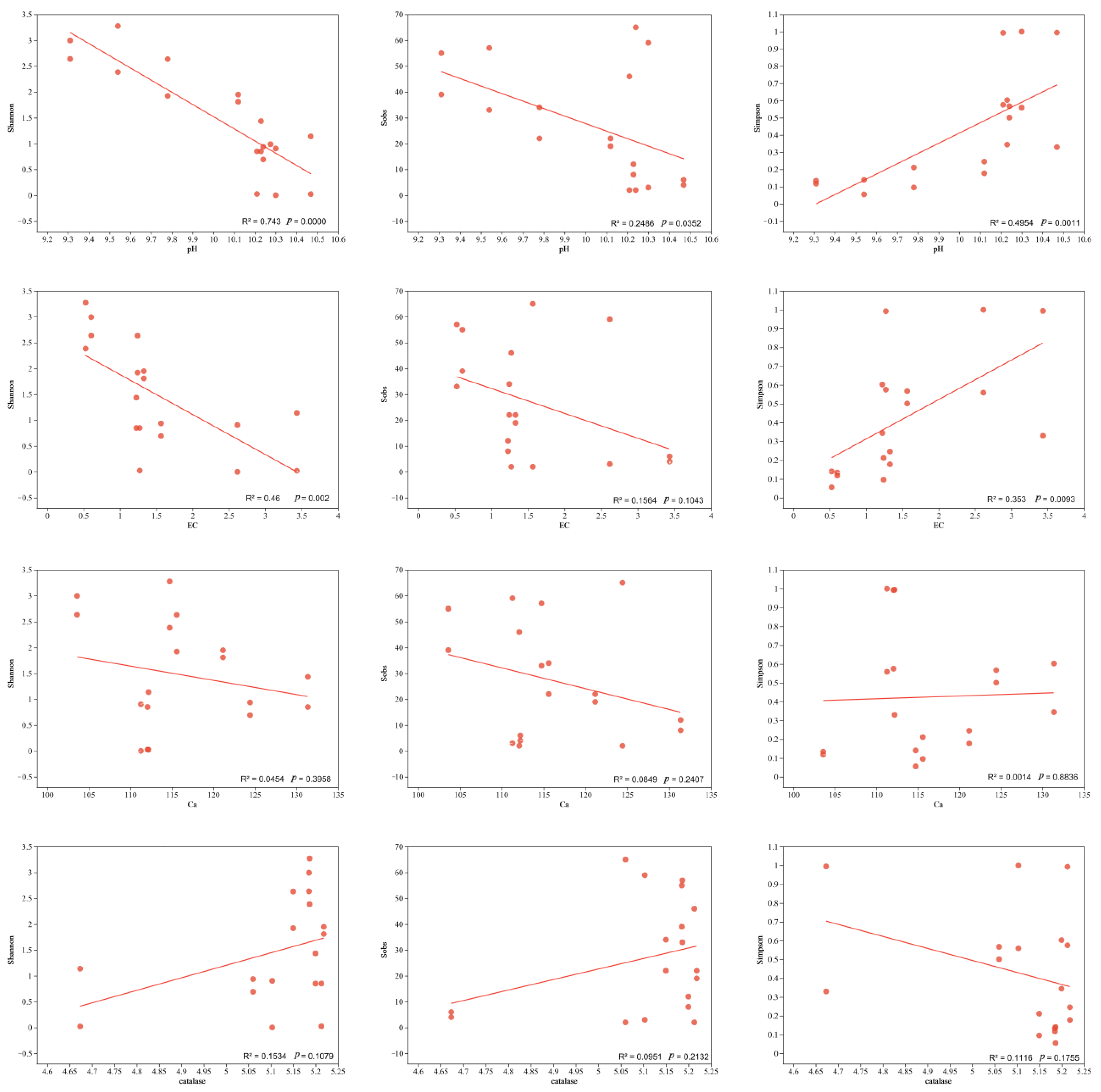
3.5. Effects of Environmental Factors on AMF Community Structure and Diversity
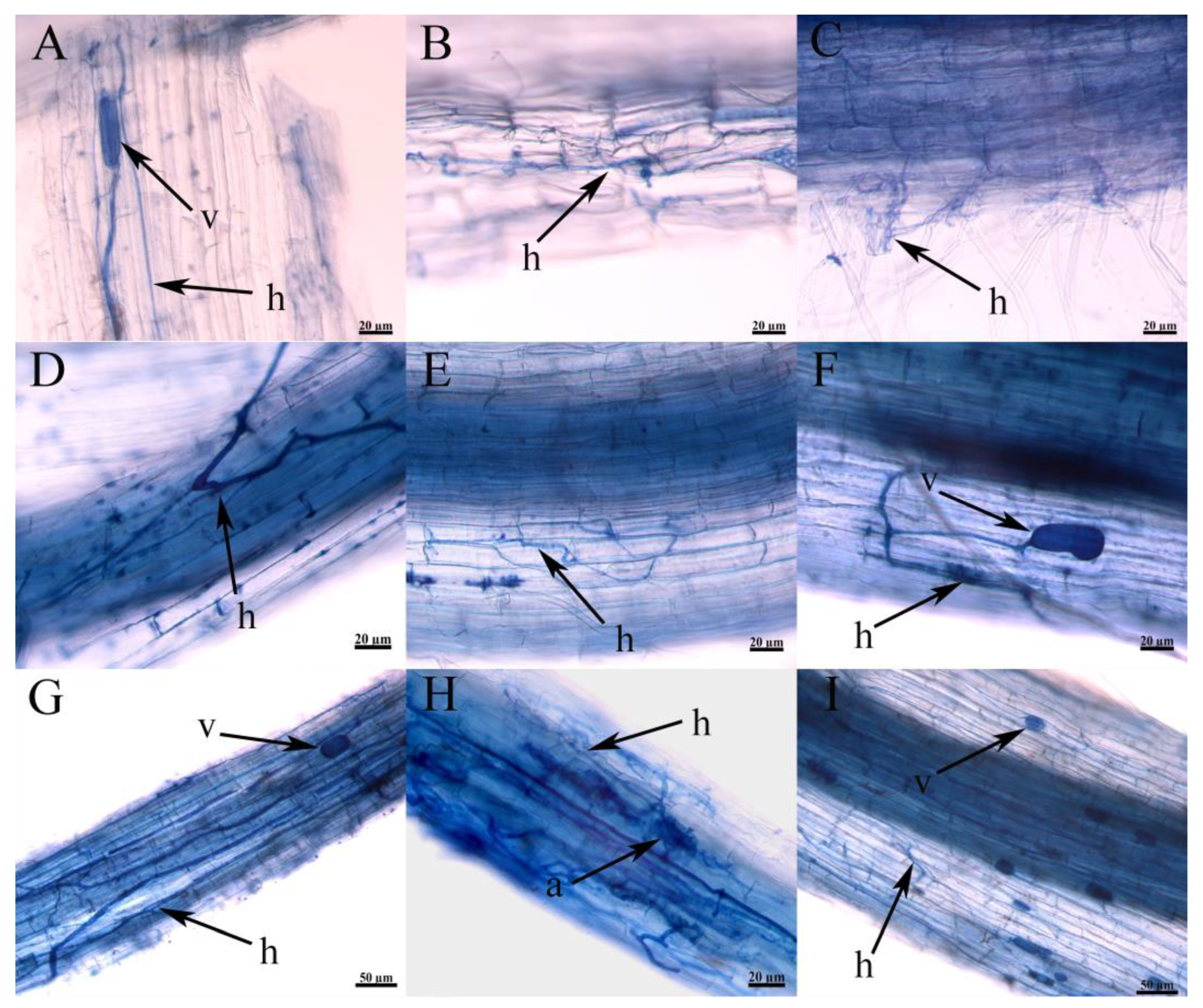
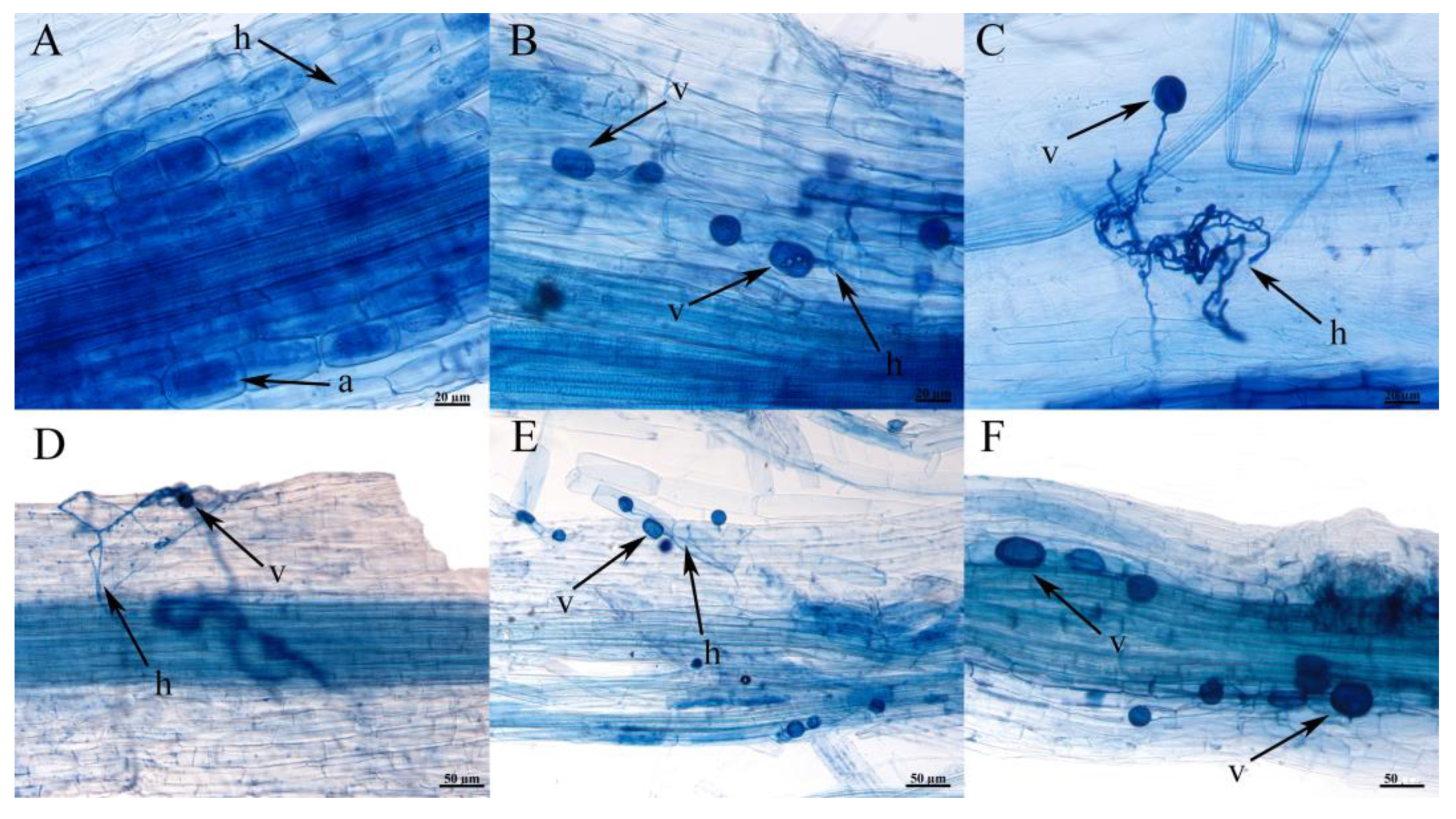
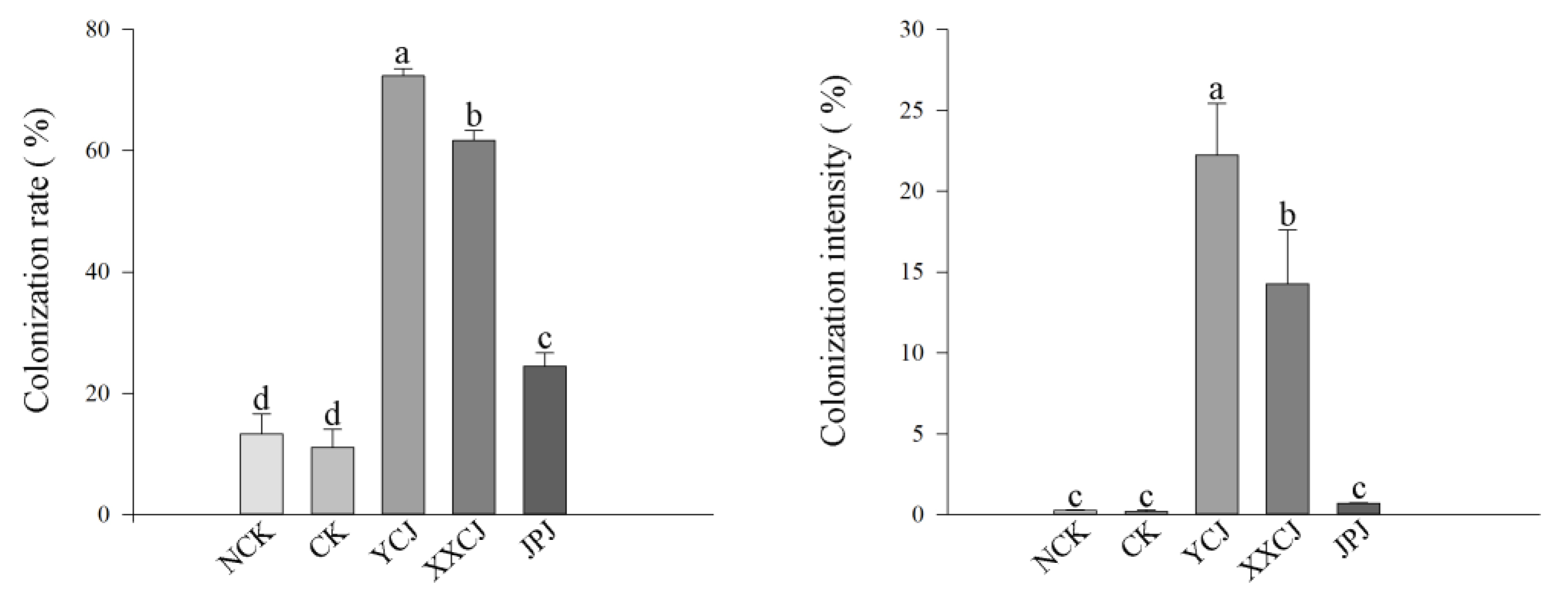
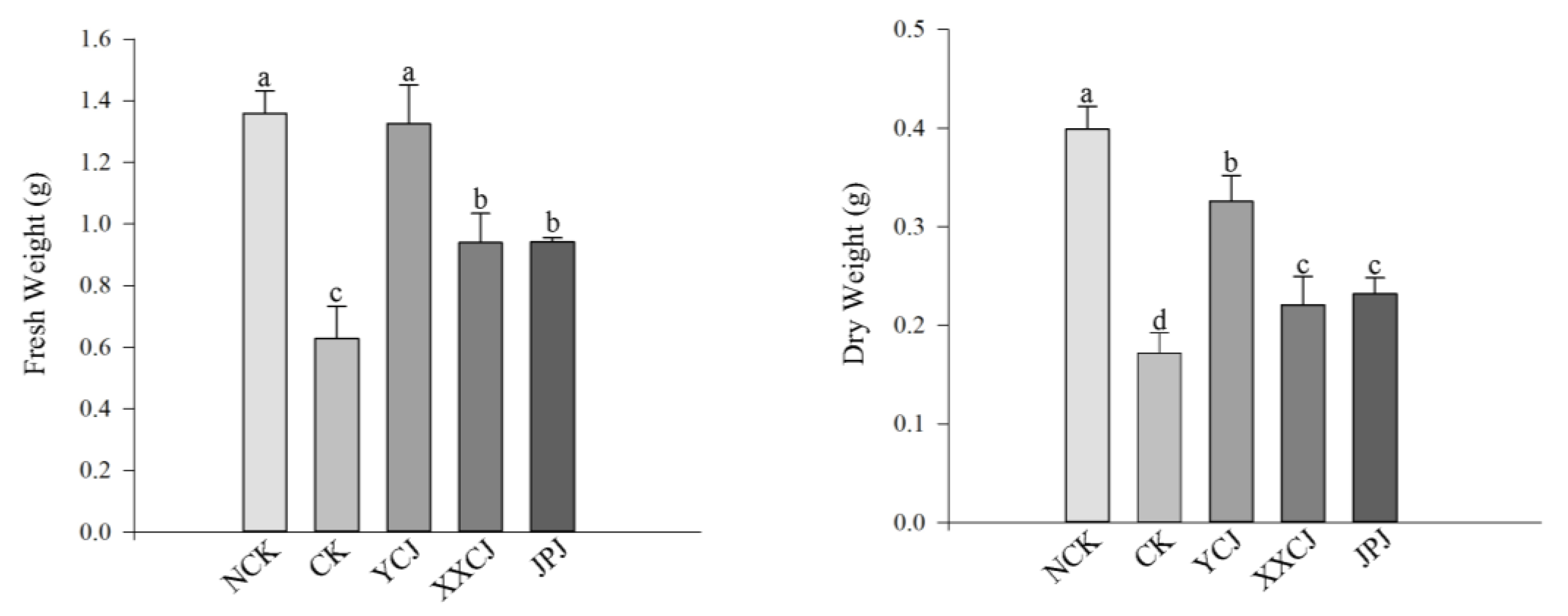
3.6. Root Colonization of Plants in the Wild and Verifying the Ecological Matching Phenomenon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, C.X.; Chen, F.; Yue, Y.N.; Yan, X.F. Diversity characteristics of arbuscular mycorrhizal fungi in the rhizosphere of twenty-six species of plants in Songnen saline-alkaline grassland. Pratacultural Sci. 2015, 32, 2008–2020. [Google Scholar]
- Liu, Y.J.; Fang, L.L.; Zhao, W.N.; Yang, C.X. Effects of Different Arbuscular Mycorrhizal Fungi on Physiology of Viola prionantha under Salt Stress. Phyton 2023, 92, 55–69. [Google Scholar] [CrossRef]
- Song, C.C.; He, Y.; Deng, W. Ecological Geochemistry of Salt-Marsh Soil in Songnen Plain; Science Press: Beijing, China, 2003. [Google Scholar]
- Wang, S.R.; Huang, Y.X. Research progress on saline-alkali land improvement in Songnen Plain. Soils Crop. 2023, 12, 206–217. [Google Scholar]
- Wang, C.Y. Salt Soil in Northeast China; Science Press: Beijing, China, 2004. [Google Scholar]
- Xu, Z.Q.; Xu, X.O. The Cause of Formation and Characteristics of Soda Saline-Alkaline Land of the Songnen Plain and the Study Progress of Control Measures. Soil Water Conserv. China 2018, 2, 54–59. [Google Scholar]
- Yan, X.F.; Sun, G.R. Study on Physiological Ecology of Puccinellia Tenuiflora; Science Press: Beijing, China, 2000. [Google Scholar]
- Chi, C.M. Study on Stress of Soda-Saline Soil in Songnen Plain; Southwest Jiaotong University Press: Chengdu, China, 2014. [Google Scholar]
- Li, R.Q.; Wang, Y.X.; Sun, Y.L.; Zhang, L.; Chen, A.P. Effects of salt stress on the growth, physiology, and biochemistry of five Bromus inermis varieties. Acta Prataculturae Sin. 2023, 32, 99–111. [Google Scholar] [CrossRef]
- Yin, L.J.; Zhang, W.Z.; Shi, D.C. Effects of soil salinization and non-capillary porosity on biomass and dominance of Leymus chinensis community. Acta Ecol. Sin. 1992, 12, 302–309. [Google Scholar]
- Garcia-Franco, N.; Wiesmeier, M.; Hurtarte, L.; Fella, F.; Martinez-Mena, M. Pruning residues incorporation and reduced tillage improve soil organic matter stabilization and structure of salt-affected soils in a semi-arid Citrus tree orchard. Soil Tillage Res. 2021, 213, 105129. [Google Scholar] [CrossRef]
- Guo, Y.J.; Du, Q.F.; Li, G.D.; Ni, Y.; Zhang, Z.; Ren, W.B.; Huo, X.Y. Soil phosphorus fractions and arbuscular mycorrhizal fungi diversity following long-term grazing exclusion on semi-arid steppes in Inner Mongolia. Geoderma 2016, 269, 79–90. [Google Scholar] [CrossRef]
- Chen, X.N.; Aili, Y.L.N.E.; Gao, W.L. Effects of inoculation with arbuscular mycorrhizal fungi on the growth and chlorophyll fluorescence characteristics of the seedlings of two typical desert riparian forest plant species under salt stress. Chin. J. Ecol. 2023, 6, 16–34. [Google Scholar]
- Xu, J.M.; Fang, L.L.; Yang, C.X. Infection and Species Diversity of Fungi in the Rhizosphere of Syneilesis aconitifolia in Songnen Saline-alkali Grassland. J. Northwest For. Univ. 2023, 38, 173–179. [Google Scholar]
- Zhao, W.N.; Yang, C.X. Investigation of AM Fungal Diversity in the Rhizosphere of Viola philippica in Songnen Saline-Alkali Grassland. J. Northeast For. Univ. 2021, 9, 016. [Google Scholar]
- Coutinho, E.S.; Beiroz, W.; Barbosa, M.; Xavier, J.H.D.; Fernandes, G.W. Arbuscular Mycorrhizal Fungi in the Rhizosphere of Saplings Used in the Restoration of the Rupestrian Grassland. Biology 2019, 37, 152–162. [Google Scholar] [CrossRef]
- Heijden, M.G.A.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T.; Wiemken, A.; Sanders, I.R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytol. 2006, 172, 739–752. [Google Scholar] [CrossRef]
- Tian, H.; Drijber, R.A.; Zhang, J.L.; Li, X.L. Impact of long-term nitrogen fertilization and rotation with soybean on the diversity and phosphorus metabolism of indigenous arbuscular mycorrhizal fungi within the roots of maize (Zea mays L.). Agric. Ecosyst. Environ. 2013, 164, 53–61. [Google Scholar] [CrossRef]
- Bencherifa, B.; Ammar, B.; Yolande, D.; Anissa, L.S. Soil and seasons affect arbuscular mycorrhizal fungi associated with Tamarix rhizosphere in arid and semi-arid steppes. Appl. Soil Ecol. 2016, 107, 180–190. [Google Scholar] [CrossRef]
- Rua, M.A.; Antoninka, A.; Antunes, P.M.; Chaudhary, V. Home-field advantage? Evidence of local adaptation among plants, soil, and arbuscular mycorrhizal fungi through meta-analysis. BMC Evol. Biol. 2016, 16, 122. [Google Scholar] [CrossRef]
- Remke, M.J.; Johnson, N.C.; Wright, J.; Williamson, M.; Bowker, M.A. Sympatric pairings of dryland grass populations, mycorrhizal fungi and associated soil biota enhance mutualism and ameliorate drought stress. J. Ecol. 2020, 109, 1210–1223. [Google Scholar] [CrossRef]
- Rekret, P.; Maherali, H. Local adaptation to mycorrhizal fungi in geographically close Lobelia siphilitica populations. Oecologia 2019, 190, 127–138. [Google Scholar] [CrossRef]
- Liz, K.; James, D.B. The missing link in grassland restoration: Arbuscular mycorrhizal fungi inoculation increases plant diversity and accelerates succession. J. Appl. Ecol. 2016, 54, 1301–1309. [Google Scholar]
- He, X.H.; Critchley, C.; Bledsoe, C. Nitrogen transfer within and between plants through common mycorrhizal networks (CMNs). Crit. Rev. Plant Sci. 2003, 22, 531–567. [Google Scholar] [CrossRef]
- Merrild, M.P.; Ambus, P.; Rosendahl, S.; Jakobsen, I. Common arbuscular mycorrhizal networks amplify competition for phosphorus between seedlings and established plants. New Phytol. 2013, 200, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.L.; Zhang, L.D.; Miao, Y.L. Effect of soil salinity-alkalinity stress and arbuscular mycorrhizal fungi on the inter-specific relationship between Leymus chinensis and Chloris virgata. J. Northeast Norm. Univ. Nat. Sci. Ed. 2014, 46, 124–129. [Google Scholar]
- Lei, G.; Hao, Z.P.; Chen, B.D. Effects of indigenous AM fungi and neighboring plants on the growth and phosphorus nutrition of Leymus chinensis. Acta Ecol. Sin. 2013, 33, 1071–1079. [Google Scholar] [CrossRef]
- Chen, S.G.; Li, H.M.; Xie, X.M. Study on Soil Sampling Techiques in Annual Crop Dryland. Agric. Res. Appl. 2022, 35, 53–58. [Google Scholar]
- Qin, J.; Shu, J.; Liu, J.; Shi, F.; Meng, Y.F. Study on the Effects of Different Media and Sampling Method on the Counting of Eurotium cristatum in Fuzhuan Tea of Jingyang. China Food Saf. Mag. 2022, 26, 66–68. [Google Scholar]
- Bao, S.D. Soil Agriculture Chemical Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Guan, Y.M. Soil Enzymes and Research Methods; China Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Wright, S.; Upadhyaya, A. Extraction of an abundant an unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575e–586. [Google Scholar] [CrossRef]
- Wang, Y.S.; Zhang, S.B.; Zhang, M.Q. Resources and Germplasm Resources of Arbuscular Mycorrhizal Fungi in China; China Agriculture Press: Beijing, China, 2012. [Google Scholar]
- Philips, J.M. Hayman D S. Improved procedures for clearing and staining vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Liu, P.; Li, M.J. Plant Physiology Experiments: Determination of Enzyme Activity and Growth Indicators, 3rd ed.; Science Press: Beijing, China, 2016; pp. 208–216. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Bo, G.; Hong, X.Z.; Wang, X.H.; Yang, S.S.; Chen, M.; Hou, A.X.; Cagle, G.A.; Han, G.X. Salt is a main factor shaping community composition of arbuscular mycorrhizal fungi along a vegetation successional series in the Yellow River Delta. Catena 2020, 185, 104318. [Google Scholar]
- Song, F.Q.; Nu, L.C.; Chang, W.; Fan, X.X.; Liu, X.; Gao, Y. Diversity of Root Nodule Endophytes from Elaeagnus angustifolia L. and Arbuscular Mycorrhizal. Chin. Agric. Sci. Bull. 2016, 32, 6–13. [Google Scholar]
- Zhang, M.Q.; Wang, Y.S.; Zhang, C.; Huang, L. Ecological distribution of some genera and species of VA mycorrhizal fungi in northern China. Mycosystema 1994, 3. [Google Scholar]
- Juniper, S.; Abbott, L.K. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza 2006, 16, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, N.; Huang, S.; Zhang, L.; Gao, X.; Yang, C. The effect of dominant AMF in the rhizosphere of Iris lactea in Songnen saline-alkali grassland on salt tolerance physiology of Petunia hybrida. Fresenius Environ. Bull. 2019, 28, 5099–5108. [Google Scholar]
- Ugyen, W.T.; Nuttapol, N.; Phanthipha, R.; Prakrit, N.; Lompong, K. Dry-Season Soil and Co-Cultivated Host Plants Enhanced Propagation of Arbuscular Mycorrhizal Fungal Spores from Sand Dune Vegetation in Trap Culture. J. Fungi 2022, 8, 1061. [Google Scholar]
- Cong, W.; Yu, J.J.; Yu, H.M.; Ding, Y.; Zhang, Y.G. Diversity and Community Assembly of Forest Soil Microorganisms in Different Climatic Zones. Sci. Silvae Sin. 2022, 58, 70–79. [Google Scholar]
- Qadir, M.; Schubert, S. Degradation processes and nutrient constraints in sodic soils. Land Degrad. Develop. 2002, 13, 275–294. [Google Scholar] [CrossRef]
- Wang, Q.B.; Yu, W.X.; Wang, N.Y.; Liu, Z.G.; Qi, X.Y. Spatial Distribution Characteristics of Soil Organic Carbon, Nitrogen and Carbon-nitrogen Ratio in Main Drylands of Jilin Province. Ournal Anhui Agric. Sci. 2020, 50, 40–42. [Google Scholar]
- Xie, X.F.; Pu, L.J.; Wang, Q.Q. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 2017, 607, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.L.; Lam, S.K.; Wolf, B. Trade-offs between soil carbon sequestration and reactive nitrogen losses under straw return in global agroecosystems. Glob. Chang. Biol. 2018, 24, 5919–5932. [Google Scholar] [CrossRef]
- Yang, C.D.; Lu, S.G. Straw and straw biochar differently affect phosphorus availability, enzyme activity and microbial functional genes in an Ultisol. Sci. Total Environ. 2022, 805, 150325. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, R.; Bharti, A.; Ramesh, A.; Prakash, A.; Sharma, M.P. Glomalin related protein and C16:1ω5 PLFA associated with AM fungi as potential signatures for assessing the soil C sequestration under contrasting soil management practices. Eur. J. Soil Biol. 2021, 103, 103286. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Liang, C.; Ai, Z.; Wu, Y.; Xu, H.; Xue, S.; Liu, G. Glomalin-related soil protein affects soil aggregation and recovery of soil nutrient following natural revegetation on the Loess Plateau. Geoderma 2020, 357, 113921. [Google Scholar] [CrossRef]
- Liu, W.J.; Ma, K.; Wang, B.B.; He, Y.L.; Jiang, Q.F.; Mi, Y. Effects of no-tillage, mulching, and organic fertilizer application on the diversity of arbuscular mycorrhizal fungi in winter wheat field. Chin. J. Ecol. 2022, 41, 1101–1108. [Google Scholar]
- Zhu, L.; Guo, K.X.; Lan, L.Y. Effects of the conversion of forest types on diversity of arbuscular mycorrhizal fungi in subalpine soil. Chin. J. Ecol. 2020, 39, 3943–3951. [Google Scholar]
- Wen, Y.X.; Zhao, B.; Luo, Q.Y.; Jia, Y.L.; Feng, T.; Wang, Q. Distribution and ecological role in close-to-nature restoration of arbuscular mycorrhizal fungi in Tibetan Plateau alpine grassland. Mycosystema 2021, 40, 2562–2578. [Google Scholar]
- Erica, L.; Jing, P.; Franco, M.; Huang, C.H.; Valeria, B.; Xue, X.; Raffaella, B.; Anna, T. Native Arbuscular Mycorrhizal Fungi Characterization from Saline Lands in Arid Oases, Northwest China. J. Fungi 2020, 6, 80. [Google Scholar]
- Haug, I.; Setaro, S.; Suarez, J.P. Global AM fungi are dominating mycorrhizal communities in a tropical premontane dry forest in Laipuna, South Ecuador. Mycol. Prog. 2021, 20, 1007. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Correia, P.M.; Cacador, I. Effects of salinity and flooding on the infectivity of salt marsh arbuscular mycorrhizal fungi in Aster tripolium L. Biol. Fertil. Soils 2003, 38, 137–143. [Google Scholar] [CrossRef]
- Santander, C.; Aroca, R.; Ruiz-lozano, J.M. Arbuscular mycorrhiza effects on plant performance under osmotic stress. Mycorrhiza 2017, 27, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gong, J. Differential effects of abiotic factors and host plant traits on diversity and community composition of root-colonizing arbuscular mycorrhizal fungi in a salt-stressed ecosystem. Mycorrhiza 2014, 24, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Yang, R. Symbiotic Adaptation of Plant and AM Fungi to Environmental Change along Tibetan Altitudinal Gradients; China Agricultural University: Beijing, China, 2017. [Google Scholar]
- Lin, L.T.; Ma, K.M. Applications of network analyses in the studies of symbiotic relationship between host plants and mycorrhizal fungi. Mycosystema 2019, 38, 1826–1839. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Stage | F% | RA% | IV% | ||
|---|---|---|---|---|---|---|---|
| III | II | I | |||||
| Glomus | G. multiforum | + | + | + | 37.04% | 2.97% | 20.01% |
| G. monosporum | + | 14.81% | 0.82% | 7.82% | |||
| G. badium | + | + | 25.93% | 1.65% | 13.79% | ||
| G. convolutum | + | + | 25.93% | 2.14% | 14.04% | ||
| G. reticulatum | + | + | + | 59.26% | 4.61% | 31.94% | |
| G. microaggregatum | + | + | + | 18.52% | 0.82% | 9.67% | |
| G. pansihalos | + | 7.41% | 0.49% | 3.95% | |||
| G. geosporum | + | + | 25.93% | 1.48% | 13.71% | ||
| G. halonatum | + | 7.41% | 0.33% | 3.87% | |||
| G. macrocarpum | + | + | 7.41% | 0.33% | 3.87% | ||
| G. hyderabadensis | + | 14.81% | 0.66% | 7.74% | |||
| G. reticulatum | + | + | + | 18.52% | 0.99% | 9.76% | |
| G. lamellosum | + | + | 18.52% | 0.82% | 9.67% | ||
| G. clarum | + | + | + | 66.67% | 6.10% | 36.39% | |
| G. fasciculatum | + | + | 29.63% | 1.81% | 15.72% | ||
| G. melanosporum | + | + | 22.22% | 1.48% | 11.85% | ||
| G. magnicaule | + | 3.70% | 0.16% | 1.93% | |||
| G. mosseae | + | 7.41% | 0.33% | 3.87% | |||
| G. fragile | + | 3.70% | 0.16% | 1.93% | |||
| G. versiforme | + | 3.70% | 0.16% | 1.93% | |||
| Acaulospora | Ac. delicata | + | 22.22% | 1.81% | 12.02% | ||
| Ac. laevis | + | + | + | 62.96% | 6.26% | 34.61% | |
| Ac. koskei | + | 11.11% | 0.49% | 5.8% | |||
| Ac. morrowiae | + | 25.93% | 1.32% | 13.63% | |||
| Ac. excavata | + | 11.11% | 0.49% | 5.8% | |||
| Ac. bireticulata | + | + | + | 40.74% | 2.80% | 21.77% | |
| Ac. denticulata | + | 3.70% | 0.16% | 1.93% | |||
| Ac. lacunosa | + | 3.70% | 0.16% | 1.93% | |||
| Ac. gerdemannii | + | + | 7.41% | 0.33% | 3.87% | ||
| Ac. dilatata | + | 7.41% | 0.33% | 3.87% | |||
| Ac. paulinae | + | 3.70% | 0.16% | 1.93% | |||
| Ac. undulata | + | + | 7.41% | 0.33% | 3.87% | ||
| Ac. rugosa | + | 3.70% | 0.16% | 1.93% | |||
| Ac. mellea | + | 3.70% | 0.16% | 1.93% | |||
| Claroideoglomus | Cl. walkeri | + | + | + | 70.37% | 5.93% | 38.15% |
| Cl. luteum | + | + | 40.74% | 2.47% | 21.61% | ||
| Cl. etunicatum | + | 7.41% | 0.33% | 3.87% | |||
| Cl. lamellosum | + | + | + | 51.85% | 4.45% | 28.15% | |
| Cl. claroideum | + | + | + | 70.07% | 6.75% | 38.41% | |
| Gigaspora | Gi. decipiens | + | 7.41% | 0.49% | 3.95% | ||
| Gi. margarita | + | 3.70% | 0.16% | 1.93% | |||
| Gi. ramisporophora | + | 3.70% | 0.16% | 1.93% | |||
| Septoglomus | Se. constrictum | + | + | + | 25.93% | 1.65% | 13.79% |
| Se. deserticola | + | + | 18.52% | 0.99% | 9.76% | ||
| Funneliformis | Fu. mosseae | + | 3.70% | 0.16% | 1.93% | ||
| Fu. verruculosum | + | 3.70% | 0.16% | 1.93% | |||
| Diversispora | Di. aurantia | + | 3.70% | 0.33% | 2.02% | ||
| Di. eburnea | + | + | + | 44.44% | 2.80% | 23.62% | |
| Pacispora | Pa. scintillans | + | + | + | 44.44% | 2.97% | 23.71% |
| Pa. chimonobambusae | + | 7.41% | 0.33% | 3.87% | |||
| Scutellospora | Scu. reticulata | + | 3.70% | 0.16% | 1.93% | ||
| Scu. calospora | + | 3.70% | 0.16% | 1.93% | |||
| Entrophospora | En. baltica | + | 7.41% | 0.33% | 3.87% | ||
| En. infrequens | + | 3.70% | 0.16% | 1.93% | |||
| Archaeospora | Ar. leptoticha | + | + | 7.41% | 0.33% | 3.87% | |
| Dominikia | Do. aurea | + | + | 7.41% | 0.33% | 3.87% | |
| Rhizophagus | Rh. intraradices | + | + | + | 62.96% | 5.11% | 34.04% |
| Racocetra | Ra. castanea | + | + | 25.93% | 1.48% | 13.71% | |
| Sclerocystis | Scl. sinuosa | + | + | 37.04% | 3.46% | 20.25% | |
| Paraglomus | Par. laccatum | + | + | 33.33% | 2.31% | 17.82% | |
| Ambispora | Am. leptoticha | + | + | + | 48.15% | 3.46% | 25.81% |
| Glomeraceae sp1 | + | + | + | 92.59% | 8.90% | 50.75% | |
| L. chinensis | P. tenuiflora | S. glauca | |
|---|---|---|---|
| pH | 9.55 ± 0.21 c | 10.19 ± 0.05 b | 10.34 ± 0.10 a |
| EC (dS·m−1) | 0.79 ± 0.34 c | 1.55 ± 0.18 b | 2.54 ± 0.81 a |
| OM (g·kg) | 57.71 ± 5.09 a | 21.59 ± 2.60 b | 11.85 ± 6.74 c |
| OC (g·kg) | 33.47 ± 2.96 a | 12.52 ± 1.51 b | 6.87 ± 3.91 c |
| CO32− (cmol·kg−1) | 2.40 ± 1.41 b | 6.98 ± 0.86 a | 8.66 ± 2.68 a |
| HCO3− (cmol·kg−1) | 4.23 ± 3.15 a | 3.28 ± 1.92 a | 2.48 ± 1.72 a |
| N (g·kg) | 1.43 ± 0.15 a | 0.93 ± 0.17 b | 0.39 ± 0.04 c |
| P (g·kg) | 0.35 ± 0.09 a | 0.30 ± 0.06 ab | 0.28 ± 0.04 b |
| K (mg·kg) | 21.85 ± 3.44 b | 24.01 ± 0.81 ab | 25.16 ± 1.72 a |
| Na (mg·kg) | 144.28 ± 8.78 b | 168.06 ± 5.92 a | 165.61 ± 4.83 a |
| Ca (mg·kg) | 111.31 ± 6.16 b | 121.53 ± 8.53 a | 115.97 ± 6.43 ab |
| Mg (mg·kg) | 6.72 ± 0.52 a | 6.60 ± 0.81 a | 6.34 ± 0.65 a |
| C/N (g·kg) | 23.50 ± 1.86 a | 14.08 ± 4.12 b | 17.52 ± 9.44 b |
| Catalase (μmol·d−1·g−1) | 5.17 ± 0.02 a | 5.21 ± 0.01 a | 4.95 ± 0.21 b |
| Sucrase (mg·d−1·g−1) | 0.33 ± 0.12 a | 0.17 ± 0.02 b | 0.10 ± 0.03 b |
| Urease (μg·d−1·g−1) | 3.10 ± 0.96 b | 4.51 ± 0.20 a | 4.28 ± 0.17 a |
| E-GRSP (mg·kg) | 0.43 ± 0.09 a | 0.20 ± 0.06 b | 0.15 ± 0.07 b |
| T-GRSP (mg·kg) | 1.32 ± 0.16 a | 0.65 ± 0.11 b | 0.42 ± 0.08 c |
| L. chinensis | P. tenuiflora | S. glauca | |
|---|---|---|---|
| colonization rate (%) | 92.592 ± 7.026 a | 63.703 ± 11.110 b | 33.333 ± 8.820 c |
| colonization intensity (%) | 29.658 ± 11.727 a | 5.030 ± 5.266 b | 0.510 ± 0.425 b |
| arbuscular abundance (%) | 2.954 ± 1.672 a | 0.763 ± 1.760 b | 0.008 ± 0.013 b |
| vesicle abundance (%) | 8.774 ± 6.570 a | 0.630 ± 1.639 b | 0.038 ± 0.037 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, L.-L.; Liu, Y.-J.; Wang, Z.-H.; Lu, X.-Y.; Li, J.-H.; Yang, C.-X. Electrical Conductivity and pH Are Two of the Main Factors Influencing the Composition of Arbuscular Mycorrhizal Fungal Communities in the Vegetation Succession Series of Songnen Saline-Alkali Grassland. J. Fungi 2023, 9, 870. https://doi.org/10.3390/jof9090870
Fang L-L, Liu Y-J, Wang Z-H, Lu X-Y, Li J-H, Yang C-X. Electrical Conductivity and pH Are Two of the Main Factors Influencing the Composition of Arbuscular Mycorrhizal Fungal Communities in the Vegetation Succession Series of Songnen Saline-Alkali Grassland. Journal of Fungi. 2023; 9(9):870. https://doi.org/10.3390/jof9090870
Chicago/Turabian StyleFang, Lin-Lin, Ya-Jie Liu, Zi-He Wang, Xiao-Yu Lu, Jin-Hua Li, and Chun-Xue Yang. 2023. "Electrical Conductivity and pH Are Two of the Main Factors Influencing the Composition of Arbuscular Mycorrhizal Fungal Communities in the Vegetation Succession Series of Songnen Saline-Alkali Grassland" Journal of Fungi 9, no. 9: 870. https://doi.org/10.3390/jof9090870