

Seasonal Variations in Fungal Communities on the Surfaces of Lan Na Sandstone Sculptures and Their Biodeterioration Capacities


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Culture-Dependent Study
2.2.1. Biodeterioration Study
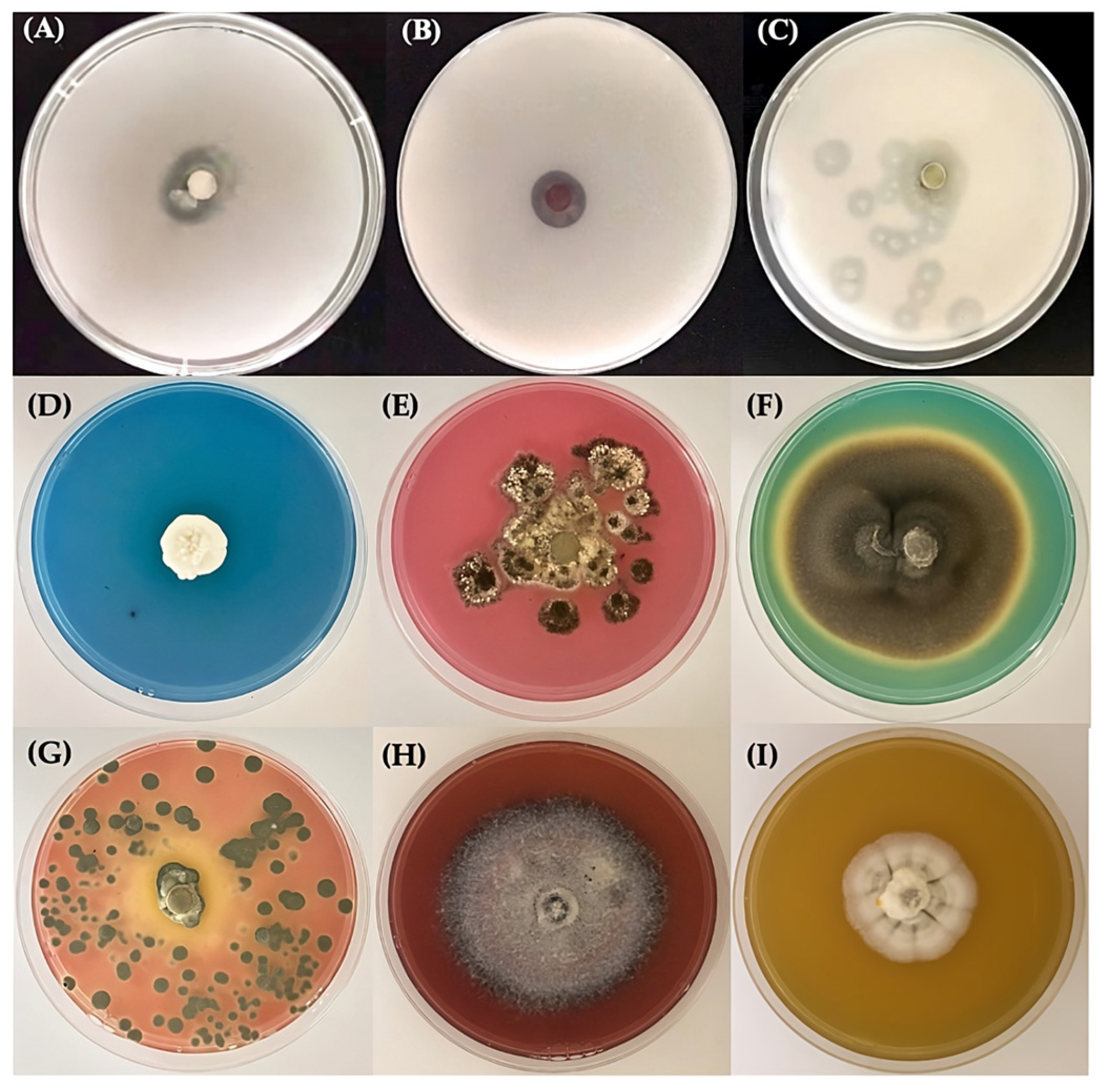
Drought Tolerance
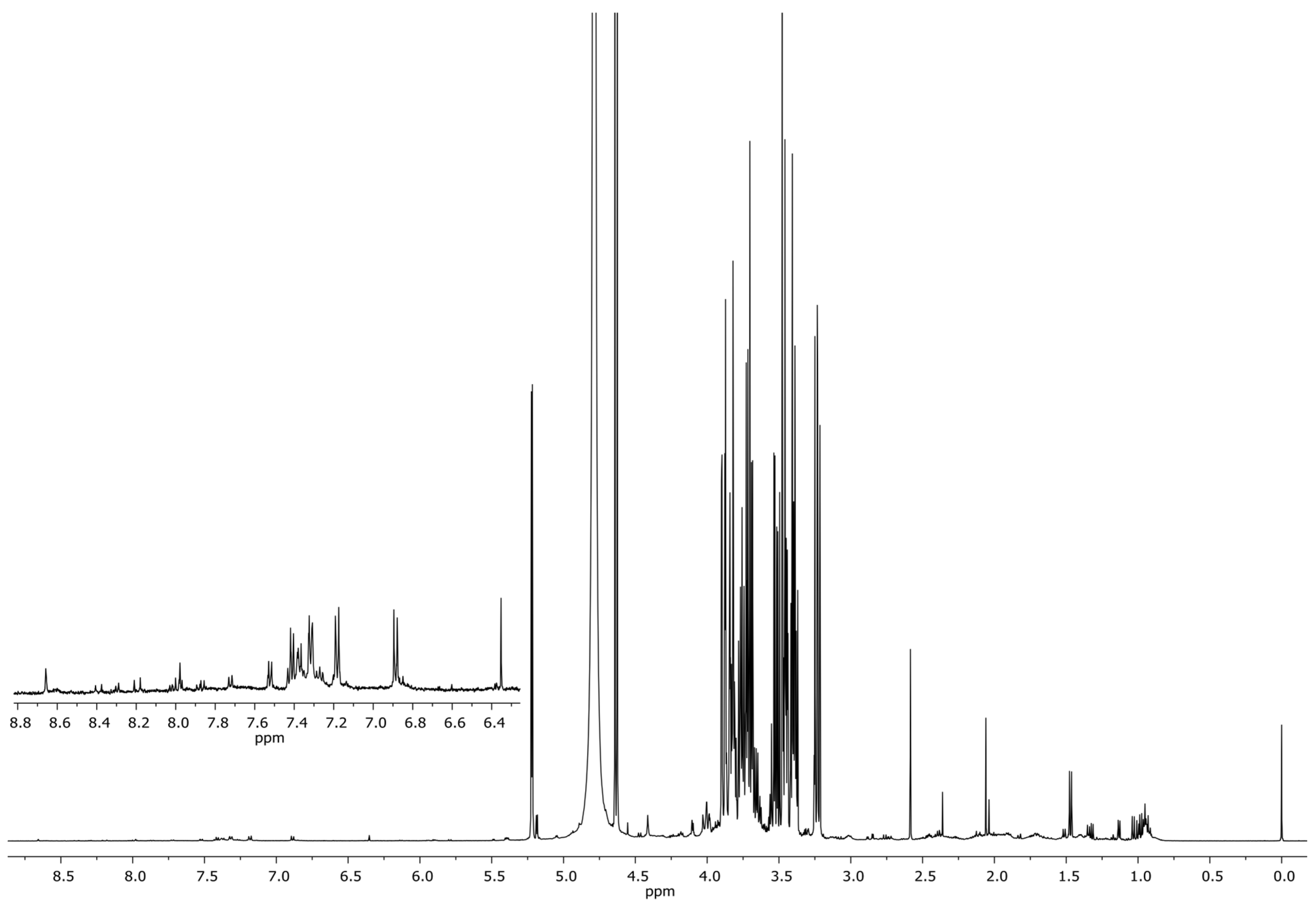
Production of Organic Acids
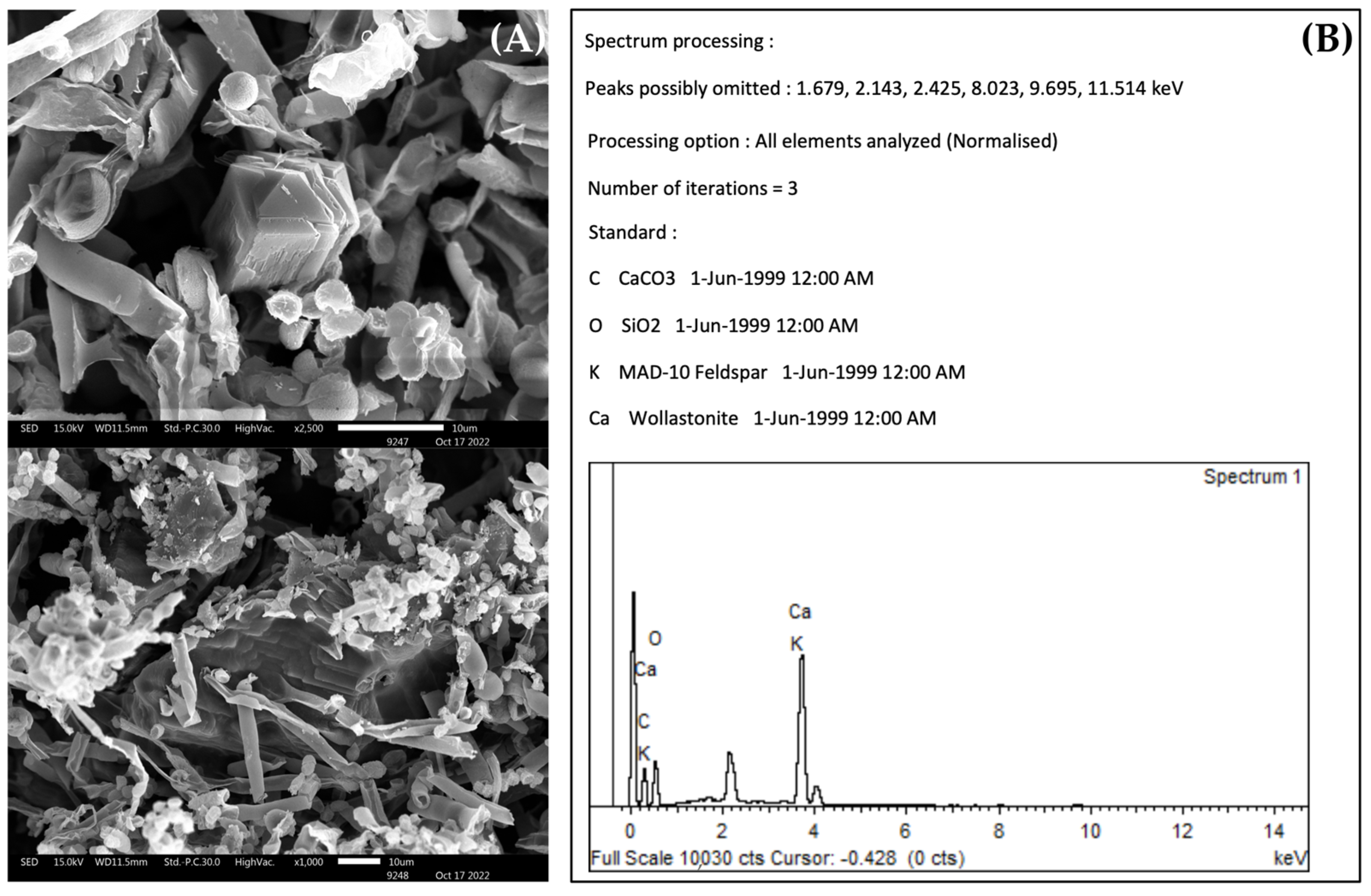
The Biomineralization of Calcium Carbonate
Calcium Carbonate Dissolution
Siderophore Production
2.2.2. Molecular Identification of Fungal Isolates
2.3. Culture-Independent Study
2.3.1. Preparation of Genomic DNA for Next-Generation Sequencing
2.3.2. Next-Generation Sequencing (NGS) of DNA for Metagenomic Sequencing
2.3.3. Alpha Diversity and Beta Diversity Analysis
2.3.4. Network Analysis
2.3.5. Functional Prediction Analysis
2.3.6. DNA Sequence Deposition
3. Results
3.1. Fungal Isolates and Drought Tolerance
3.2. The Production of Organic Acids
3.3. The Biomineralization and Dissolution of Calcium Carbonate
3.4. Siderophore Production
3.5. The Molecular Identification of Fungal Isolates
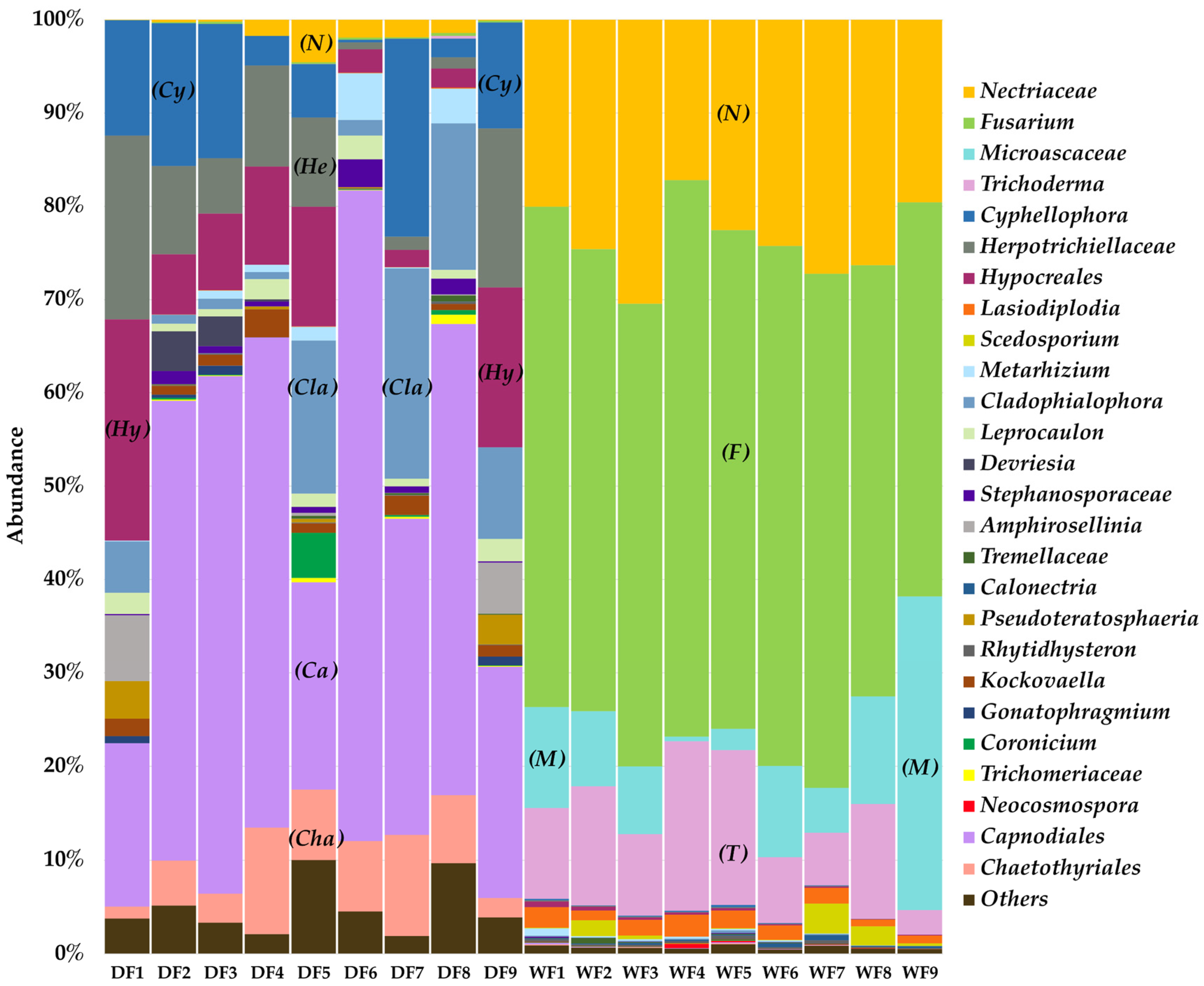
3.6. Fungal Community Structures in Sandstone Sculptures in Wet and Dry Seasons, Characterized via a Culture-Independent Molecular Technique
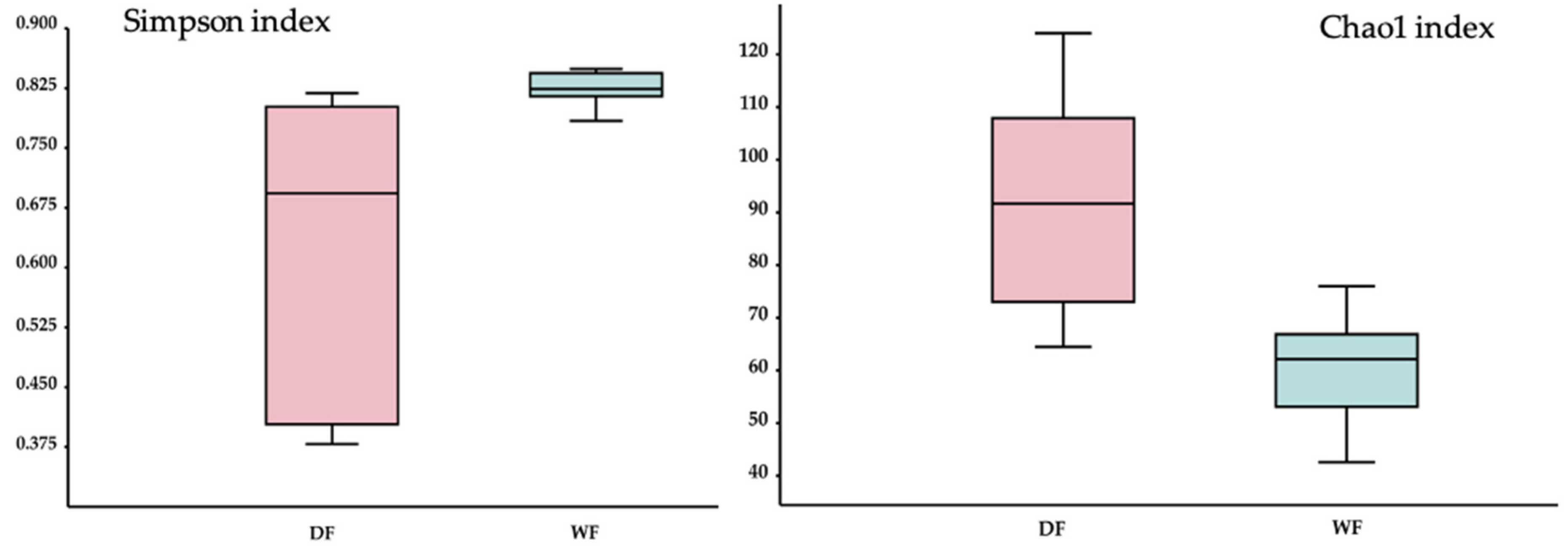
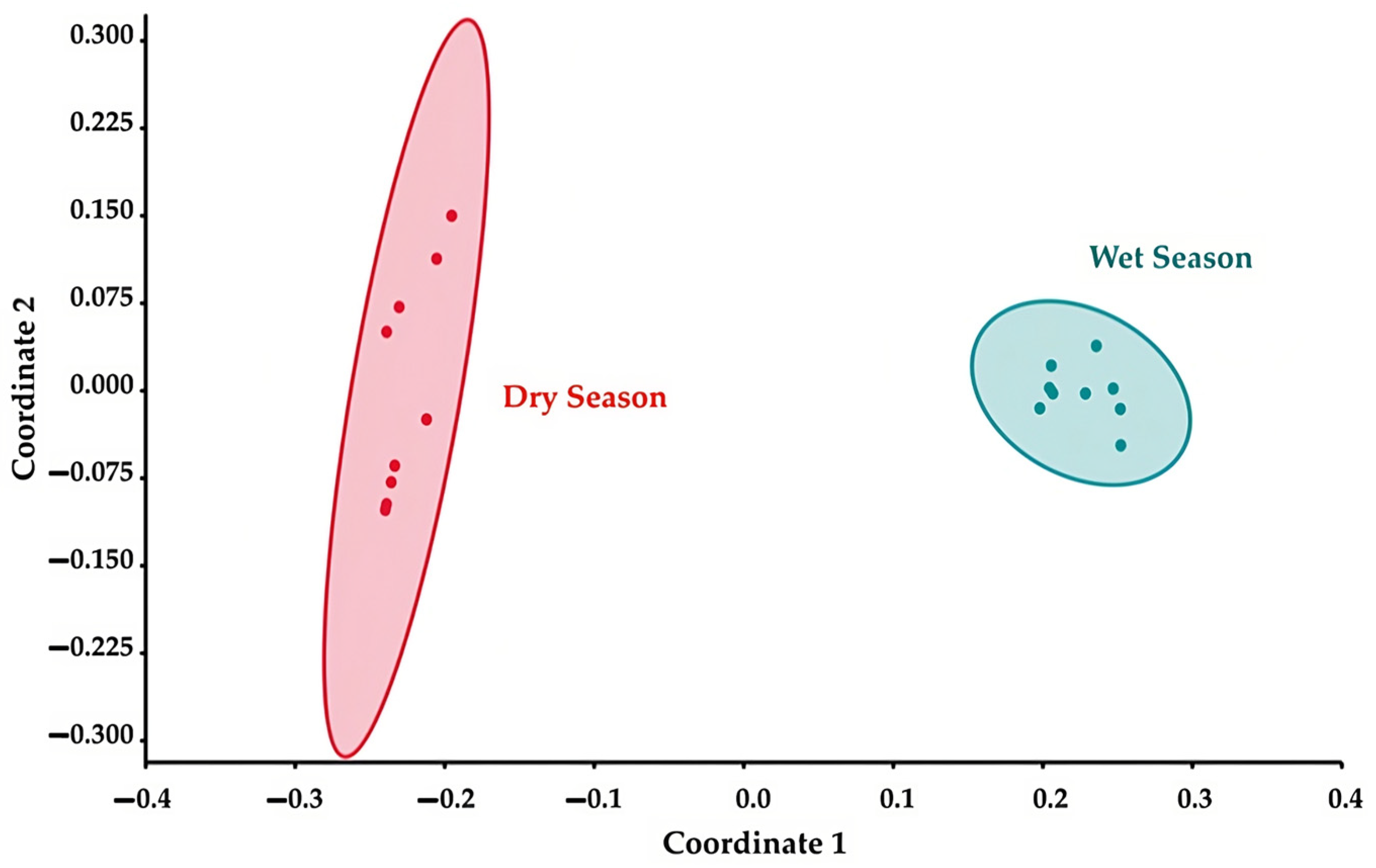
3.7. Alpha and Beta Diversity
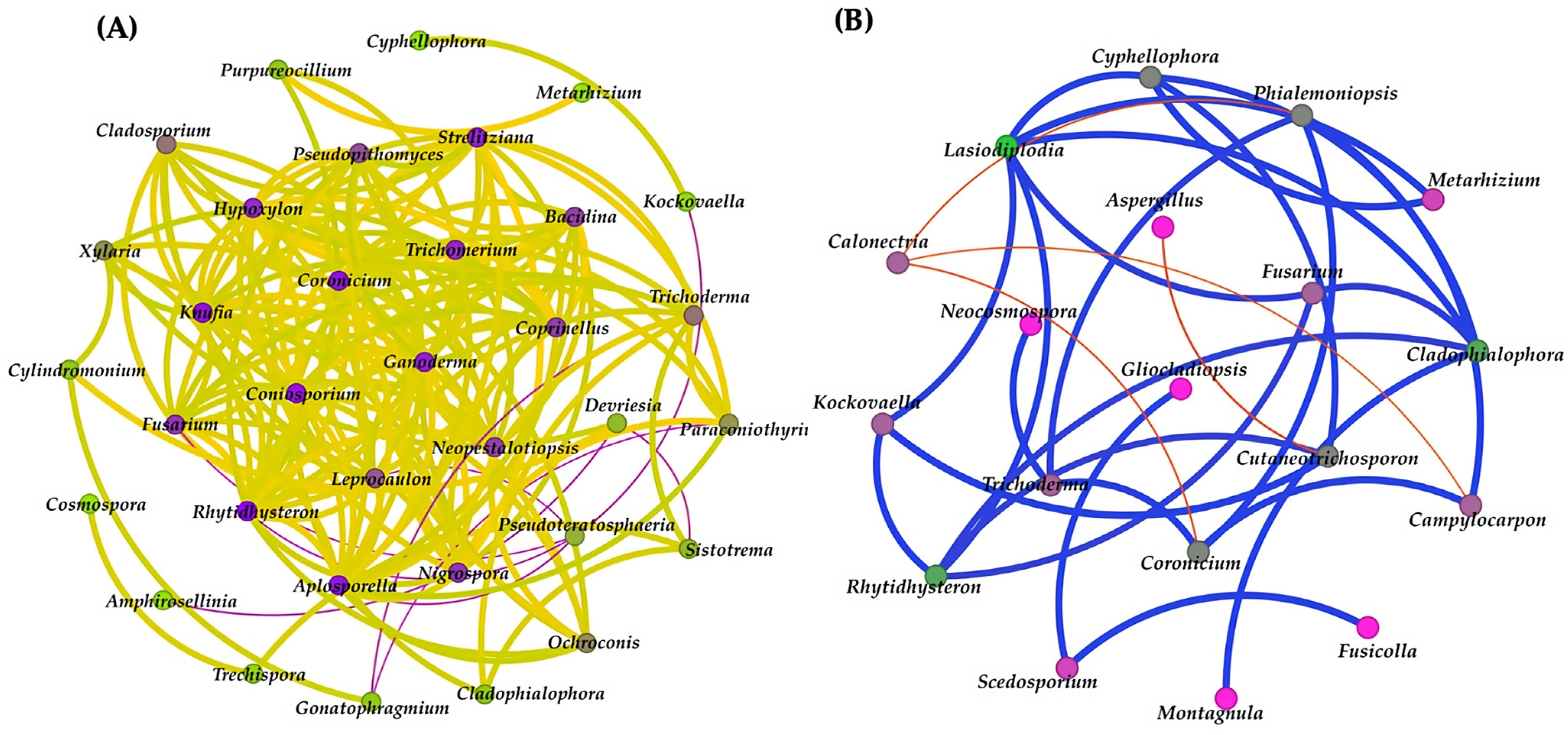
3.8. Network Analysis
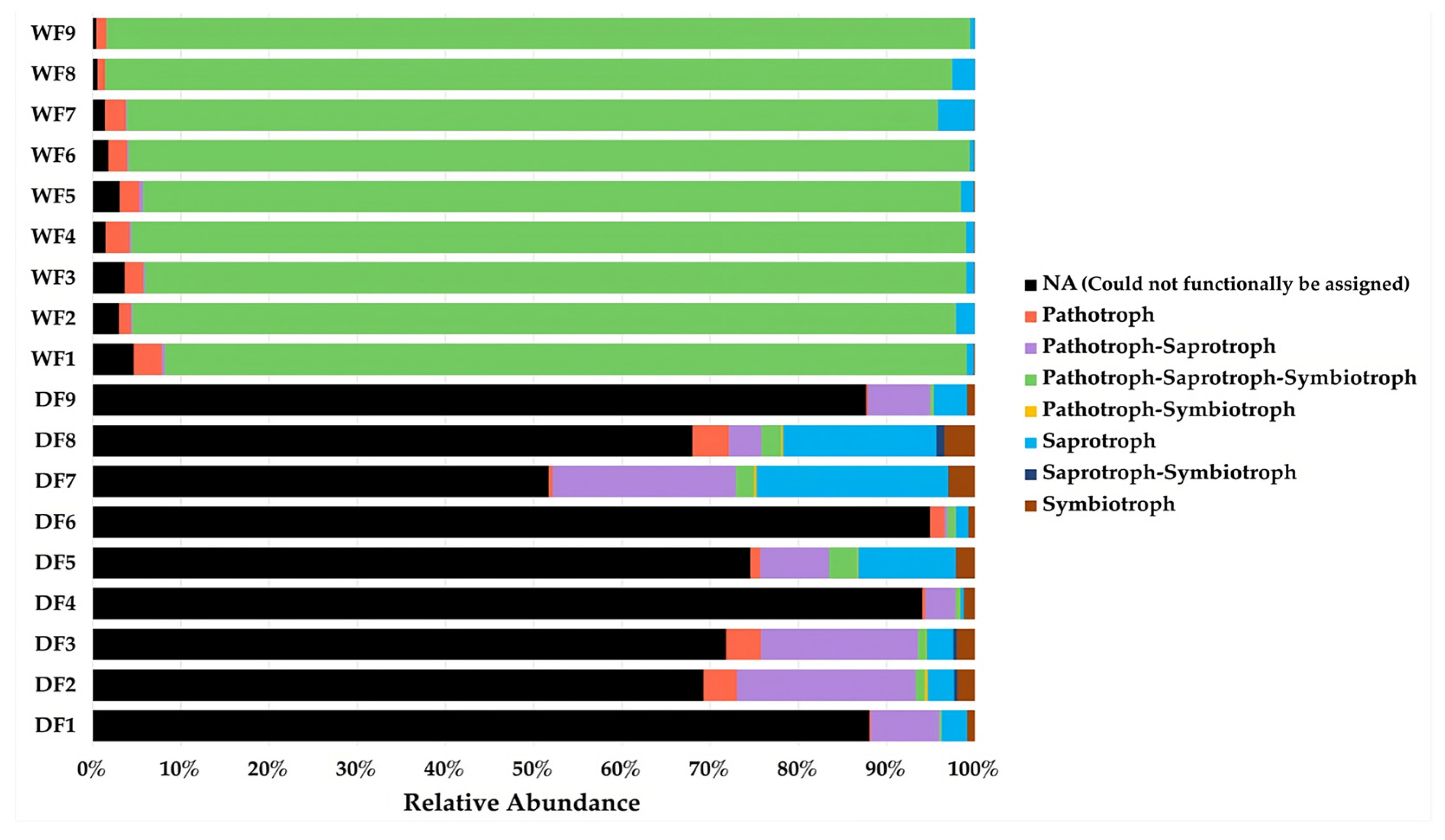
3.9. Functional Prediction Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, G.; Gong, C.; Gu, J.; Katayama, Y.; Someya, T.; Gu, J.D. Biochemical reactions and mechanisms involved in the biodeterioration of stone world cultural heritage under the tropical climate conditions. Int. Biodeterior. Biodegrad. 2019, 143, e104723. [Google Scholar] [CrossRef]
- Evidente, A. Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants. Microorganisms 2023, 11, 843. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Wu, F.; He, D.; Gu, J.D.; Feng, H.; Chen, T.; Liu, G.; Wang, W. Bacterial and fungal communities in the sandstone biofilms of two famous Buddhist grottoes in China. Int. Biodeterior. Biodegrad. 2021, 163, e105267. [Google Scholar] [CrossRef]
- Griffin, P.S.; Indictor, N.; Koestler, R.J. The Biodeterioration of Stone: A Review of Deterioration Mechanisms, Conservation Case Histories, and Treatment. Int. Biodeter. 1991, 28, 187–207. [Google Scholar] [CrossRef]
- Potysz, A.; Bartz, W. Bioweathering of minerals and dissolution assessment by experimental simulations—Implications for sandstone rocks: A review. Constr. Build. Mater. 2022, 316, e125862. [Google Scholar] [CrossRef]
- Shimadzu, H.; Dornelas, M.; Henderson, P.A.; Magurran, A.E. Diversity is maintained by seasonal variation in species abundance. BMC Biol. 2013, 11, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Bhadauria, S.; Kumar, V.; Chauhan, R.S. Biodeterioration of sandstone under the influence of different humidity lev-els in laboratory conditions. Build. Environ. 2009, 44, 1276–1284. [Google Scholar] [CrossRef]
- Luo, S.; He, B.; Zeng, Q.; Li, N.; Yang, L. Effects of seasonal variation on soil microbial community structure and enzyme ac-tivity in a Masson pine forest in Southwest China. J. Mt. Sci. 2020, 17, 1398–1409. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, F.; Liu, Y.; Wu, M.; Wang, S.; Sun, H.J.; Liu, G.; Zhang, Y.; Cui, X.; Zhang, W.; et al. The Distribution and Influencing Factors of Hypolithic Microbial Communities in the Hexi Corridor. Microorganisms 2023, 11, 1212. [Google Scholar] [CrossRef]
- Wu, G.; Wang, Y.; Swift, G.; Chen, J. Laboratory Investigation of the Effects of Temperature on the Mechanical Properties of Sandstone. Geotech. Geol. Eng. 2013, 31, 809–816. [Google Scholar] [CrossRef]
- Tiyapairat, S.; Mahabhumi, T. Quaternary Geology of Changwat Phayao Sheet (4947 II) and Amphoe Mae Chai Sheet (4947 I); Department of Mineral Resources: Bangkok, Thailand, 1990; Volume 143, pp. 36–37. [Google Scholar]
- Bunopas, S.; Khositanont, S. Did Shan-Thai twice marry Indochina and then India?: A Review. Bull. Earth Sci. Thail. (BEST) 2008, 1, 1–27. [Google Scholar]
- Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics Low Template DNA Collection View Project. In PCR Protocols: A Guide to Methods and Applications; Wiley: Hobboken, NJ, USA, 2021; Volume 38, pp. 315–322. [Google Scholar]
- Usyk, M.; Zolnik, C.P.; Patel, H.; Levi, M.H.; Burk, R.D. Novel ITS1 Fungal Primers for Characterization of the Mycobiome. mSphere 2017, 17, e00488. [Google Scholar] [CrossRef] [Green Version]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks Visuali-zation and Exploration of Large Graphs. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal. Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Milagres, A.M.; Machuca, A.; Napoleaó Napoleaó, D. Detection of siderophore production from several fungi and bacteria by a modification of chrome azurol S (CAS) agar plate assay. J. Microbiol. Methods 1999, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal. Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Liu, X.; Liu, X.; Song, W. Fungal communities in the biofilms colonizing the basalt sculptures of the Lei-zhou Stone Dogs and assessment of a conservation measure. Herit. Sci. 2021, 9, 36. [Google Scholar] [CrossRef]
- Gutarowska, B.; Czyżowska, A. The ability of filamentous fungi to produce acids on indoor building materials. Ann. Microbiol. 2009, 59, 807–813. [Google Scholar] [CrossRef]
- Del, S.; De La Rosa-García, C.; Ortega-Morales, O.; Gaylarde, C.C.; Beltrán-García, M.; Quintana-Owen, P.; Reyes-Estebanez, M. Influence of fungi in the weathering of limestone of Mayan monuments Autor para correspondencia: B.O. Ortega-Morales beorte-ga@uacam.mx ORIGINAL. Rev. Mex. Micol. 2011, 33, 43–51. [Google Scholar]
- Poole, R.K. Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 1999; Volume 41, pp. 1–236. [Google Scholar]
- Zhang, Y.; Su, M.; Wu, F.; Gu, J.D.; Li, J.; He, D.; Guo, Q.; Cui, H.; Zhang, Q.; Feng, H. Diversity and Composition of Culturable Mi-croorganisms and Their Biodeterioration Potentials in the Sandstone of Beishiku Temple, China. Microorganisms 2023, 11, 429. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M.; Bahri-Esfahani, J.; Li, Q.; Rhee, Y.J.; Wei, Z.; Fomina, M.; Liang, X. Oxalate production by fungi: Significance in geo-mycology, biodeterioration and bioremediation. Fungal. Biol. Rev. 2014, 28, 36–55. [Google Scholar] [CrossRef]
- Yu, C.; Wang, X.; Huang, S.; Zhang, X.; An, X.; Xu, M.; Lian, B. Characterisation of products from fungally modified wollas-tonite and the simulation experiment of Pb2+ fixation. Sci. Total Environ. 2021, 760, e143357. [Google Scholar] [CrossRef]
- Renshaw, J.C.; Robson, G.D.; Trinci, A.P.J.; Wiebe, M.G.; Livens, F.R.; Collison, D.; Taylor, R.J. Fungal siderophores: Structures, functions and applications. Mycol. Res. 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Haas, H. Molecular genetics of fungal siderophore biosynthesis and uptake: The role of siderophores in iron uptake and storage. Appl. Microbiol. Biotechnol. 2003, 62, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Holinsworth, B.; Martin, J.D. Siderophore production by marine-derived fungi. BioMetals 2009, 22, 625–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, B.N.; Callicott, K.A.; Cotty, P.J. Conservation and loss of a putative iron utilization gene cluster among genotypes of aspergillus flavus. Microorganisms 2021, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Y.; Kong, S. Effects of Trichoderma asperellum and its siderophores on endogenous auxin in Arabidopsis thaliana under iron-deficiency stress. Int. Microbiol. 2020, 23, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.; Holmström, S.J.M. Siderophores in environmental research: Roles and applications. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Hussein, K.A.; Joo, J.H. Zinc ions affect siderophore production by fungi isolated from the panax ginseng rhi-zosphere. J. Microbiol. Biotechnol. 2019, 29, 105–113. [Google Scholar] [CrossRef]
- Oide, S.; Berthiller, F.; Wiesenberger, G.; Adam, G.; Turgeon, B.G. Individual and combined roles of malonichrome, ferricrocin, and TAFC siderophores in Fusarium graminearum pathogenic and sexual development. Front. Microbiol. 2014, 5, 759. [Google Scholar] [CrossRef]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic secondary metabolites from fungi. Toxins 2021, 13, 261. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 28, 187–207. [Google Scholar] [CrossRef]
- Mametova, L.F.; Mirek, A.; Kozii, Y.S. Pyritization of the Middle Carboniferous Sandstones of the Donbas. Mineral. J. 2020, 42, 14–19. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef] [PubMed]
- Trovão, J.; Tiago, I.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Catarino, L.; Gil, F.; Portugal, A. Description of Aeminia-ceae fam. Nov., Aeminium gen. Nov. and Aeminium ludgeri sp. Nov. (Capnodiales), isolated from a biodeteriorated art-piece in the Old Cathedral of Coimbra, Portugal. MycoKeys 2019, 45, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Suphaphimol, N.; Suwannarach, N.; Purahong, W.; Jaikang, C.; Pengpat, K.; Semakul, N.; Yimklan, S.; Jongjitngam, S.; Jindasu, S.; Thiangtham, S.; et al. Identification of Microorganisms Dwelling on the 19th Century Lan Na Mural Paintings from Northern Thailand Using Culture-Dependent and-Independent Approaches. Biology 2022, 11, 228. [Google Scholar] [CrossRef] [PubMed]
- Lombardozzi, V.; Castrignanò, T.; D’Antonio, M.; Casanova Municchia, A.; Caneva, G. An interactive database for an ecological analysis of stone biopitting. Int. Biodeter. Biodegrad. 2012, 73, 8–15. [Google Scholar] [CrossRef]
- Pląskowska, E.; Patejuk, K.; Lorenc, M.W.; Grzeszczuk, J. Lichens and Fungi on Sandstone Tombs at the Historical Jewish Cemetery in Wrocław (Poland). Stud. Conserv. 2023, 20, 1–9. [Google Scholar] [CrossRef]
- De Leo, F.; Marchetta, A.; Urzì, C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Appl. Sci. 2022, 12, 3969. [Google Scholar] [CrossRef]
- Feng, P.; Lu, Q.; Najafzadeh, M.J.; Gerrits Van Den Ende, A.H.G.; Sun, J.; Li, R.; Xi, L.; Vicente, V.A.; Lai, W.; Lu, C.; et al. Cy-phel-lophora and its relatives in Phialophora: Biodiversity and possible role in human infection. Fungal Divers. 2014, 65, 17–45. [Google Scholar] [CrossRef] [Green Version]
- Paiva, D.S.; Fernandes, L.; Pereira, E.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal). J. Fungi 2023, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.; Fritzlar, D.; Stannek, L.; Hoppert, M. Ascomycete fungi on dimension stone of the “Burg Gleichen”, Thuringia. Environ. Earth. Sci. 2011, 63, 1713–1722. [Google Scholar] [CrossRef] [Green Version]
- Joseph, E. Microorganisms in the Deterioration and Preservation of Cultural Heritage; Springer Nature: Berlin/Heidelberg, Germany, 2021; p. 367. [Google Scholar]
- Geweely, N.S.I.; Afifi, H.A.M. Bioremediation of some deterioration products from sandstone of archeological Karnak tem-ple using stimulated irradiated alkalo-thermophilic purified microbial enzymes. Geomicrobiol. J. 2011, 28, 56–67. [Google Scholar] [CrossRef]
- Pandey, A.; Shrivastav, A.; Bhatnagar, P.; Sarsaiya, S.; Awasthi, M.; Mandir, B.; Singh Chatri, B.; Mahal, R.; Mahal, K.; Ka Mandir, T. Diversity of Monument Deterioration-Causing Fungi at Gwalior Fort (M.P.) India. Ann. Environ. Sci. 2011, 5, 35–40. [Google Scholar]
- Li, J.; Deng, M.; Gao, L.; Yen, S.; Katayama, Y.; Gu, J.D. The active microbes and biochemical processes contributing to deteri-ora-tion of Angkor sandstone monuments under the tropical climate in Cambodia—A review. J. Cult. Herit. 2021, 47, 218–226. [Google Scholar] [CrossRef]
- Dupont, J.; Jacquet, C.; Dennetière, B.; Lacoste, S.; Bousta, F.; Orial, G.; Cruaud, C.; Couloux, A.; Roquebert, M.F. Invasion of the French paleolithic painted cave of Lascaux by members of the Fusarium solani species complex. Mycologia 2007, 4, 526–533. [Google Scholar] [CrossRef]
- Farooq, M. Mycobial Deterioration of Stone Monuments of Dharmarajika, Taxila. J. Microbiol. Exp. 2015, 2, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Rojas, T.I.; Aira, M.J.; Batista, A.; Cruz, I.L.; González, S. Fungal biodeterioration in historic buildings of Havana (Cuba). Grana 2012, 51, 44–51. [Google Scholar] [CrossRef]
- Abdelhafez, A.A.M.; El-Wekeel, F.M.; Ramadan, E.M.; Abed-Allah, A.A. Microbial deterioration of archaeological marble: Identification and treatment. Ann. Agric. Sci. 2012, 57, 137–144. [Google Scholar] [CrossRef]
- Le Thi, L.; Mertens, A.; Toan Vu, D.; Tuong Vu, D.; Le, P.; Minh, A.; Duc, N.; De Backer, S.; Swennen, R.; Vandelook, F.; et al. Genetic diversity of the pathogenic banana Fusarium wilt in northern Viet Nam. bioRxiv 2021, 5, 4–6. [Google Scholar]
- Villarino, M.; De la Lastra, E.; Basallote-Ureba, M.J.; Capote, N.; Larena, I.; Melgarejo, P.; De Cal, A. Charac-terization of fusarium solani populations associated with Spanish strawberry crops. Plant. Dis. 2019, 103, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Arie, T. Fusarium diseases of cultivated plants, control, diagnosis, and molecular and genetic studies. J. Pestic. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Fu, R.; Wu, B.; Liu, X.; Xiang, M. Rock-inhabiting fungi: Terminology, diversity, evolution, and adaptation mechanisms. Mycology 2022, 13, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, C.; Gueidan, C.; Selbmann, L.; Gorbushina, A.A.; Crous, P.W.; Groenewald, J.Z.; Muggia, L.; Grube, M.; Isola, D.; Schoch, C.L.; et al. Phylogeny of rock-inhabiting fungi related to Dothideomycetes. Stud. Mycol 2009, 64, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Talley, S.M.; Coley, P.D.; Kursar, T.A. The effects of weather on fungal abundance and richness among 25 communities in the Intermountain West. BMC Ecol. 2002, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- El-Derby, A.A.O.D.; Mansour, M.M.A.; Salem, M.Z.M. Investigation the microbial deterioration of sandstone from the osi-rion’s sarcophagus chamber as affected by rising ground water level. Mediterr. Archaeol. Archaeom. 2016, 16, 273–281. [Google Scholar]
- Erdmann, E.A.; Nitsche, S.; Gorbushina, A.A.; Schumacher, J. Genetic Engineering of the Rock Inhabitant Knufia petricola Pro-vides Insight Into the Biology of Extremotolerant Black Fungi. Front. Fungal Biol. 2022, 3, e862429. [Google Scholar] [CrossRef]
- Breitenbach, R.; Gerrits, R.; Dementyeva, P.; Knabe, N.; Schumacher, J.; Feldmann, I.; Radnik, J.; Ryo, M.; Gorbushina, A.A. The role of extracellular polymeric substances of fungal biofilms in mineral attachment and weathering. NPJ Mater. Degrad. 2022, 6, 42. [Google Scholar] [CrossRef]
- Ortega-Morales, B.O.; Narváez-Zapata, J.; Reyes-Estebanez, M.; Quintana, P.; De la Rosa-García, S.C.; Bullen, H.; Gómez-Cornelio, S.; Chan-Bacab, M.J. Bioweathering potential of cultivable fungi associated with semi-arid surface microhabitats of mayan buildings. Front. Microbiol. 2016, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the con-solidated species concept to resolve species in the teratosphaeriaceae. Pers. Mol. Phylogeny Evol. Fungi 2014, 33, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Castillo, J.A.; Oyama, K.; Vázquez-Garcidueñas, M.S.; Aguilar-Romero, R.; García-Oliva, F.; Vázquez-Marrufo, G. Fungal community and ligninolytic enzyme activities in Quercus deserticola Trel. litter from forest fragments with increasing levels of disturbance. Forests 2017, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Liu, X.; Cai, L.; Guo, L.; et al. Unveiling the hidden diversity of rock-inhabiting fungi: Chaetothyriales from China. J. Fungi 2020, 6, 187. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, J.; Kiyuna, T.; Nishijima, M.; An, K.D.; Nagatsuka, Y.; Tazato, N.; Handa, Y.; Hata-Tomita, J.; Sato, Y.; Kigawa, R.; et al. Polyphasic insights into the microbiomes of the Takamatsuzuka Tumulus and Kitora Tumulus. J. Gen. Appl. Microbiol. 2017, 63, 63–113. [Google Scholar] [CrossRef] [Green Version]
- Badali, H.; Gueidan, C.; Najafzadeh, M.J.; Bonifaz, A.; Gerrits van den Ende, A.H.G.; de Hoog, G.S. Biodiversity of the genus Cladophialophora. Stud. Mycol. 2008, 61, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Savković, Ž.; Stupar, M.; Unković, N.; Knežević, A.; Vukojević, J.; Grbić, M.L. Fungal Deterioration of Cultural Heritage Objects. In Biodegradation Technology of Organic and Inorganic Pollutants; Intechopen: London, UK, 2021; pp. 267–288. [Google Scholar]
- Cappitelli, F.; Nosanchuk, J.D.; Casadevall, A.; Toniolo, L.; Brusetti, L.; Florio, S.; Principi, P.; Borin, S.; Sorlini, C. Synthetic consolidants attacked by melanin-producing fungi: Case study of the biodeterioration of Milan (Italy) ca-thedral marble treated with acrylics. Appl. Environ. Microbiol. 2007, 73, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Zhong, Z.; Liu, H.; Lin, L.; Guo, M.; Guo, W.; Wang, Z.; Zhang, Q.; Feng, L.; Lu, G.; et al. Whole genome and transcrip-tome analysis reveal adaptive strategies and pathogenesis of Calonectria pseudoreteaudii to Eucalyptus. BMC Genom. 2018, 19, 358. [Google Scholar] [CrossRef] [Green Version]
- Ochi, S.; Yoshida, M.; Nakagawa, A.; Natsume, M. Identification, and activity of a phytotoxin produced by Calonectria ilic-icola, the causal agent of soybean red crown rot. Can. J. Plant. Pathol. 2011, 33, 347–354. [Google Scholar]
- Fu, Z.; Chen, Q.; Lei, P.; Xiang, W.; Ouyang, S.; Chen, L. Soil fungal communities and enzyme activities along local tree species diversity gradient in subtropical evergreen forest. Forests 2021, 12, 1321. [Google Scholar] [CrossRef]
- El-Gendi, H.; Saleh, A.K.; Badierah, R.; Redwan, E.M.; El-Maradny, Y.A.; El-Fakharany, E.M. A Comprehensive Insight into Fungal Enzymes: Structure, Classification, and Their Role in Mankind’s Challenges. J. Fungi 2022, 8, 23. [Google Scholar] [CrossRef]
- Kirtzel, J.; Madhavan, S.; Wielsch, N.; Blinne, A.; Hupfer, Y.; Linde, J.; Krause, K.; Svatoš, A.; Kothe, E. Enzymatic Bioweath-er-ing and Metal Mobilization from Black Slate by the Basidiomycete Schizophyllum commune. Front. Microbiol. 2018, 9, 2545. [Google Scholar] [CrossRef] [Green Version]
- Seifert, A.G.; Trumbore, S.; Xu, X.; Zhang, D.; Kothe, E.; Gleixner, G. Variable effects of labile carbon on the carbon use of dif-ferent microbial groups in black slate degradation. Geochim. Cosmochim Acta 2011, 75, 2557–2570. [Google Scholar] [CrossRef] [Green Version]
- Páramo-Aguilera, L.; Ortega-Morales, B.O.; Narváez-Zapata, J.A. Culturable fungi associated with urban stone surfaces in Mexico City. Electron. J. Biotechnol. 2012, 15, 4. [Google Scholar]
- Farah, N.; Karim, A.; Mohd, M.; Mohd, N.; Nor, I.M.; Zakaria, L. Saprophytic and Potentially Pathogenic Fusarium Species from Peat Soil in Perak and Pahang. Trop. Life Sci. Res. 2016, 27, 1. [Google Scholar]
- Isola, D.; Bartoli, F.; Meloni, P.; Caneva, G.; Zucconi, L. Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Appl. Sci. 2022, 12, 2038. [Google Scholar] [CrossRef]
- Bui, A.; Orr, D.; Lepori-Bui, M.; Konicek, K.; Young, H.S.; Moeller, H.V. Soil fungal community composition and functional similarity shift across distinct climatic conditions. FEMS. Microbiol. Ecol. 2020, 96, 193. [Google Scholar] [CrossRef]
- Aschenbrenner, I.A.; Cernava, T.; Berg, G.; Grube, M. Understanding microbial multi-species symbioses. Front. Microbiol. 2016, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.O.; Templer, P.H.; Sorensen, P.O.; Sanders-DeMott, R.; Groffman, P.M.; Bhatnagar, J.M. Soil Microbes Trade-Off Bio-geochemical Cycling for Stress Tolerance Traits in Response to Year-Round Climate Change. Front. Microbiol. 2020, 11, 616. [Google Scholar] [CrossRef]
- Liu, X.; Koestler, R.J.; Warscheid, T.; Katayama, Y.; Gu, J.D. Microbial deterioration and sustainable conservation of stone monuments and buildings. Nat. Sustain. 2020, 3, 991–1004. [Google Scholar] [CrossRef]
- Quamer, T.; Hassan, M.K.; Kamal, M.A. Role of Micro-Organisms in Biodeterioration of Sandstone in Heritage Buildings. J. Adv. Res. Const. Urban. Arch. 2020, 6, 1–9. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonthijun, P.; Mills, N.; Mills, N.; Yongsawas, R.; Sansupa, C.; Suwannarach, N.; Jaikang, C.; Motanated, K.; Chayapakdee, P.; Jongjitngam, S.; et al. Seasonal Variations in Fungal Communities on the Surfaces of Lan Na Sandstone Sculptures and Their Biodeterioration Capacities. J. Fungi 2023, 9, 833. https://doi.org/10.3390/jof9080833
Nonthijun P, Mills N, Mills N, Yongsawas R, Sansupa C, Suwannarach N, Jaikang C, Motanated K, Chayapakdee P, Jongjitngam S, et al. Seasonal Variations in Fungal Communities on the Surfaces of Lan Na Sandstone Sculptures and Their Biodeterioration Capacities. Journal of Fungi. 2023; 9(8):833. https://doi.org/10.3390/jof9080833
Chicago/Turabian StyleNonthijun, Paradha, Natasha Mills, Nantana Mills, Rujipas Yongsawas, Chakriya Sansupa, Nakarin Suwannarach, Churdsak Jaikang, Kannipa Motanated, Pattarasuda Chayapakdee, Surachai Jongjitngam, and et al. 2023. "Seasonal Variations in Fungal Communities on the Surfaces of Lan Na Sandstone Sculptures and Their Biodeterioration Capacities" Journal of Fungi 9, no. 8: 833. https://doi.org/10.3390/jof9080833