
The Yeast Protein Kinase Sch9 Functions as a Central Nutrient-Responsive Hub That Calibrates Metabolic and Stress-Related Responses
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Structure of Sch9 and Confirmed Phosphosites
3. Lipid-Dependent Regulation of Sch9
4. The Role of Sch9 in Metabolic Reprogramming and Stress Responses
5. The Involvement of Sch9 in Proteasomal Degradation and Autophagy
6. The Role of Sch9 on Longevity Modulation
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kingsbury, J.M.; Sen, N.D.; Cardenas, M.E. Branched-Chain Aminotransferases Control TORC1 Signaling in Saccharomyces cerevisiae. PLoS Genet. 2015, 11, e1005714. [Google Scholar] [CrossRef] [PubMed]
- Péli-Gulli, M.-P.; Sardu, A.; Panchaud, N.; Raucci, S.; De Virgilio, C. Amino Acids Stimulate TORC1 through Lst4-Lst7, a GTPase-Activating Protein Complex for the Rag Family GTPase Gtr2. Cell Rep. 2015, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracka, D.; Jozefczuk, S.; Rudroff, F.; Sauer, U.; Hall, M.N. Nitrogen Source Activates TOR (Target of Rapamycin) Complex 1 via Glutamine and Independently of Gtr/Rag Proteins. J. Biol. Chem. 2014, 289, 25010–25020. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, M.; Maeda, T. An in Vitro TORC1 Kinase Assay That Recapitulates the Gtr-Independent Glutamine-Responsive TORC1 Activation Mechanism on Yeast Vacuoles. Mol. Cell. Biol. 2017, 37, e00075-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukai, H.; Araki, Y.; Kira, S.; Oikawa, Y.; May, A.I.; Noda, T. Gtr/Ego-Independent TORC1 Activation Is Achieved through a Glutamine-Sensitive Interaction with Pib2 on the Vacuolar Membrane. PLoS Genet. 2018, 14, e1007334. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Hall, M.N. Nutrient Sensing and TOR Signaling in Yeast and Mammals. EMBO J. 2017, 36, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.; Soulard, A.; Huber, A.; Lippman, S.; Mukhopadhyay, D.; Deloche, O.; Wanke, V.; Anrather, D.; Ammerer, G.; Riezman, H.; et al. Sch9 Is a Major Target of TORC1 in Saccharomyces Cerevisiae. Mol. Cell 2007, 26, 663–674. [Google Scholar] [CrossRef]
- Huber, A.; French, S.L.; Tekotte, H.; Yerlikaya, S.; Stahl, M.; Perepelkina, M.P.; Tyers, M.; Rougemont, J.; Beyer, A.L.; Loewith, R. Sch9 Regulates Ribosome Biogenesis via Stb3, Dot6 and Tod6 and the Histone Deacetylase Complex RPD3L. EMBO J. 2011, 30, 3052–3064. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Weisman, L.S. The Vacuole/Lysosome Is Required for Cell-Cycle Progression. eLife 2015, 4, e08160. [Google Scholar] [CrossRef]
- Jorgensen, P.; Rupeš, I.; Sharom, J.R.; Schneper, L.; Broach, J.R.; Tyers, M. A Dynamic Transcriptional Network Communicates Growth Potential to Ribosome Synthesis and Critical Cell Size. Genes Dev. 2004, 18, 2491. [Google Scholar] [CrossRef] [Green Version]
- Roosen, J.; Engelen, K.; Marchal, K.; Mathys, J.; Griffioen, G.; Cameroni, E.; Thevelein, J.M.; De Virgilio, C.; De Moor, B.; Winderickx, J. PKA and Sch9 Control a Molecular Switch Important for the Proper Adaptation to Nutrient Availability. Mol. Microbiol. 2005, 55, 862–880. [Google Scholar] [CrossRef]
- Swinnen, E.; Ghillebert, R.; Wilms, T.; Winderickx, J. Molecular Mechanisms Linking the Evolutionary Conserved TORC1-Sch9 Nutrient Signalling Branch to Lifespan Regulation in Saccharomyces Cerevisiae. FEMS Yeast Res. 2014, 14, 17–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorimitsu, T.; Zaman, S.; Broach, J.R.; Klionsky, D.J. Protein Kinase A and Sch9 Cooperatively Regulate Induction of Autophagy in Saccharomyces Cerevisiae. Mol. Biol. Cell 2007, 18, 4180–4189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, T.; Cameron, S.; Sass, P.; Wigler, M. SCH9, a Gene of Saccharomyces Cerevisiae That Encodes a Protein Distinct from, but Functionally and Structurally Related to, CAMP-Dependent Protein Kinase Catalytic Subunits. Genes Dev. 1988, 2, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Malia, P.C.; Hatakeyama, R.; Nicastro, R.; Hu, Z.; Péli-Gulli, M.P.; Gao, J.; Nishimura, T.; Eskes, E.; Stefan, C.J.; et al. TORC1 Determines Fab1 Lipid Kinase Function at Signaling Endosomes and Vacuoles. Curr. Biol. 2021, 31, 297–309.e8. [Google Scholar] [CrossRef] [PubMed]
- Novarina, D.; Guerra, P.; Milias-Argeitis, A. Vacuolar Localization via the N-Terminal Domain of Sch9 Is Required for TORC1-Dependent Phosphorylation and Downstream Signal Transduction. J. Mol. Biol. 2021, 433, 167326. [Google Scholar] [CrossRef] [PubMed]
- Wilms, T.; Swinnen, E.; Eskes, E.; Dolz-Edo, L.; Uwineza, A.; Van Essche, R.; Rosseels, J.; Zabrocki, P.; Cameroni, E.; Franssens, V.; et al. The Yeast Protein Kinase Sch9 Adjusts V-ATPase Assembly/Disassembly to Control PH Homeostasis and Longevity in Response to Glucose Availability. PLoS Genet. 2017, 13, e1006835. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Zhang, X.; Lester, R.L.; Dickson, R.C. The Sphingoid Long Chain Base Phytosphingosine Activates AGC-Type Protein Kinases in Saccharomyces Cerevisiae Including Ypk1, Ypk2, and Sch9. J. Biol. Chem. 2005, 280, 22679–22687. [Google Scholar] [CrossRef] [Green Version]
- Friant, S.; Lombardi, R.; Schmelzle, T.; Hall, M.N.; Riezman, H. Sphingoid Base Signaling via Pkh Kinases Is Required for Endocytosis in Yeast. EMBO J. 2001, 20, 6783–6792. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Gruhler, A.; Liu, Y.; Jensen, O.N.; Dickson, R.C. The Sphingolipid Long-Chain Base-Pkh1/2-Ypk1/2 Signaling Pathway Regulates Eisosome Assembly and Turnover. J. Biol. Chem. 2008, 283, 10433–10444. [Google Scholar] [CrossRef] [Green Version]
- Roelants, F.M.; Baltz, A.G.; Trott, A.E.; Fereres, S.; Thorner, J. A Protein Kinase Network Regulates the Function of Aminophospholipid Flippases. Proc. Natl. Acad. Sci. USA 2010, 107, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Roelants, F.M.; Leskoske, K.L.; Martinez Marshall, M.N.; Locke, M.N.; Thorner, J. The TORC2-Dependent Signaling Network in the Yeast Saccharomyces Cerevisiae. Biomolecules 2017, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelants, F.M.; Torrance, P.D.; Thorner, J. Differential Roles of PDK1- and PDK2-Phosphorylation Sites in the Yeast AGC Kinases Ypk1, Pkc1 and Sch9. Microbiol. Read. Engl. 2004, 150, 3289–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voordeckers, K.; Kimpe, M.; Haesendonckx, S.; Louwet, W.; Versele, M.; Thevelein, J.M. Yeast 3-Phosphoinositide-Dependent Protein Kinase-1 (PDK1) Orthologs Pkh1-3 Differentially Regulate Phosphorylation of Protein Kinase A (PKA) and the Protein Kinase B (PKB)/S6K Ortholog Sch9. J. Biol. Chem. 2011, 286, 22017–22027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deprez, M.-A.; Caligaris, M.; Rosseels, J.; Hatakeyama, R.; Ghillebert, R.; Sampaio-Marques, B.; Mudholkar, K.; Eskes, E.; Meert, E.; Ungermann, C.; et al. The Nutrient-Responsive CDK Pho85 Primes the Sch9 Kinase for Its Activation by TORC1. PLoS Genet. 2023, 19, e1010641. [Google Scholar] [CrossRef]
- Jin, Y.; Jin, N.; Oikawa, Y.; Benyair, R.; Koizumi, M.; Wilson, T.E.; Ohsumi, Y.; Weisman, L.S. Bur1 Functions with TORC1 for Vacuole-Mediated Cell Cycle Progression. EMBO Rep. 2022, 23, e53477. [Google Scholar] [CrossRef]
- Caligaris, M.; Nicastro, R.; Hu, Z.; Tripodi, F.; Hummel, J.E.; Pillet, B.; Deprez, M.-A.; Winderickx, J.; Rospert, S.; Coccetti, P.; et al. Snf1/AMPK Fine-Tunes TORC1 Signaling in Response to Glucose Starvation. eLife 2023, 12, e84319. [Google Scholar] [CrossRef]
- Lu, J.-Y.; Lin, Y.-Y.; Sheu, J.-C.; Wu, J.-T.; Lee, F.-J.; Chen, Y.; Lin, M.-I.; Chiang, F.-T.; Tai, T.-Y.; Berger, S.L.; et al. Acetylation of Yeast AMPK Controls Intrinsic Aging Independently of Caloric Restriction. Cell 2011, 146, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudholkar, K.; Fitzke, E.; Prinz, C.; Mayer, M.P.; Rospert, S. The Hsp70 Homolog Ssb Affects Ribosome Biogenesis via the TORC1-Sch9 Signaling Pathway. Nat. Commun. 2017, 8, 937. [Google Scholar] [CrossRef] [Green Version]
- Soulard, A.; Cremonesi, A.; Moes, S.; Schütz, F.; Jenö, P.; Hall, M.N. The Rapamycin-Sensitive Phosphoproteome Reveals That TOR Controls Protein Kinase A toward Some but Not All Substrates. Mol. Biol. Cell 2010, 21, 3475–3486. [Google Scholar] [CrossRef] [Green Version]
- Galello, F.; Portela, P.; Moreno, S.; Rossi, S. Characterization of Substrates That Have a Differential Effect on Saccharomyces Cerevisiae Protein Kinase A Holoenzyme Activation. J. Biol. Chem. 2010, 285, 29770–29779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plank, M. Interaction of TOR and PKA Signaling in S. Cerevisiae. Biomolecules 2022, 12, 210. [Google Scholar] [CrossRef]
- Dokládal, L.; Stumpe, M.; Hu, Z.; Jaquenoud, M.; Dengjel, J.; De Virgilio, C. Phosphoproteomic Responses of TORC1 Target Kinases Reveal Discrete and Convergent Mechanisms That Orchestrate the Quiescence Program in Yeast. Cell Rep. 2021, 37, 110149. [Google Scholar] [CrossRef]
- Smets, B.; Ghillebert, R.; De Snijder, P.; Binda, M.; Swinnen, E.; De Virgilio, C.; Winderickx, J. Life in the Midst of Scarcity: Adaptations to Nutrient Availability in Saccharomyces Cerevisiae. Curr. Genet. 2010, 56, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Pozza, F.; Pletcher, S.D.; Gendron, C.M.; Longo, V.D. Regulation of Longevity and Stress Resistance by Sch9 in Yeast. Science 2001, 292, 288–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio-Marques, B.; Burhans, W.C.; Ludovico, P. Longevity Pathways and Maintenance of the Proteome: The Role of Autophagy and Mitophagy during Yeast Ageing. Microb. Cell Graz Austria 2014, 1, 118–127. [Google Scholar] [CrossRef]
- Wei, M.; Fabrizio, P.; Hu, J.; Ge, H.; Cheng, C.; Li, L.; Longo, V.D. Life Span Extension by Calorie Restriction Depends on Rim15 and Transcription Factors Downstream of Ras/PKA, Tor, and Sch9. PLoS Genet. 2008, 4, e13. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Lu, K.-Y.; Tao, S.-C.; Yang, T.-C.; Ho, Y.-H.; Lee, C.-H.; Lin, C.-C.; Juan, H.-F.; Huang, H.-C.; Yang, C.-Y.; Chen, M.-S.; et al. Profiling Lipid-Protein Interactions Using Nonquenched Fluorescent Liposomal Nanovesicles and Proteome Microarrays. Mol. Cell. Proteom. MCP 2012, 11, 1177–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobko, A. Systems Biology of AGC Kinases in Fungi. Sci. STKE Signal Transduct. Knowl. Environ. 2006, 2006, re9. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.R.; Komander, D.; Alessi, D.R. The Nuts and Bolts of AGC Protein Kinases. Nat. Rev. Mol. Cell Biol. 2010, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Truebestein, L.; Hornegger, H.; Anrather, D.; Hartl, M.; Fleming, K.D.; Stariha, J.T.B.; Pardon, E.; Steyaert, J.; Burke, J.E.; Leonard, T.A. Structure of Autoinhibited Akt1 Reveals Mechanism of PIP3-Mediated Activation. Proc. Natl. Acad. Sci. USA 2021, 118, e2101496118. [Google Scholar] [CrossRef] [PubMed]
- Frech, M.; Andjelkovic, M.; Ingley, E.; Reddy, K.K.; Falck, J.R.; Hemmings, B.A. High Affinity Binding of Inositol Phosphates and Phosphoinositides to the Pleckstrin Homology Domain of RAC/Protein Kinase B and Their Influence on Kinase Activity. J. Biol. Chem. 1997, 272, 8474–8481. [Google Scholar] [CrossRef] [Green Version]
- James, S.R.; Downes, C.P.; Gigg, R.; Grove, S.J.; Holmes, A.B.; Alessi, D.R. Specific Binding of the Akt-1 Protein Kinase to Phosphatidylinositol 3,4,5-Trisphosphate without Subsequent Activation. Biochem. J. 1996, 315 Pt 3, 709–713. [Google Scholar] [CrossRef]
- Thomas, C.C.; Deak, M.; Alessi, D.R.; van Aalten, D.M.F. High-Resolution Structure of the Pleckstrin Homology Domain of Protein Kinase b/Akt Bound to Phosphatidylinositol (3,4,5)-Trisphosphate. Curr. Biol. 2002, 12, 1256–1262. [Google Scholar] [CrossRef]
- Corbalan-Garcia, S.; Gómez-Fernández, J.C. Signaling through C2 Domains: More than One Lipid Target. Biochim. Biophys. Acta 2014, 1838, 1536–1547. [Google Scholar] [CrossRef] [Green Version]
- Thorner, J. TOR Complex 2 Is a Master Regulator of Plasma Membrane Homeostasis. Biochem. J. 2022, 479, 1917–1940. [Google Scholar] [CrossRef]
- Kamada, Y.; Fujioka, Y.; Suzuki, N.N.; Inagaki, F.; Wullschleger, S.; Loewith, R.; Hall, M.N.; Ohsumi, Y. Tor2 Directly Phosphorylates the AGC Kinase Ypk2 to Regulate Actin Polarization. Mol. Cell. Biol. 2005, 25, 7239–7248. [Google Scholar] [CrossRef] [Green Version]
- Leskoske, K.L.; Roelants, F.M.; Martinez Marshall, M.N.; Hill, J.M.; Thorner, J. The Stress-Sensing TORC2 Complex Activates Yeast AGC-Family Protein Kinase Ypk1 at Multiple Novel Sites. Genetics 2017, 207, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Martinez Marshall, M.N.; Emmerstorfer-Augustin, A.; Leskoske, K.L.; Zhang, L.H.; Li, B.; Thorner, J. Analysis of the Roles of Phosphatidylinositol-4,5-Bisphosphate and Individual Subunits in Assembly, Localization, and Function of Saccharomyces Cerevisiae Target of Rapamycin Complex 2. Mol. Biol. Cell 2019, 30, 1555–1574. [Google Scholar] [CrossRef] [PubMed]
- Roelants, F.M.; Breslow, D.K.; Muir, A.; Weissman, J.S.; Thorner, J. Protein Kinase Ypk1 Phosphorylates Regulatory Proteins Orm1 and Orm2 to Control Sphingolipid Homeostasis in Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 2011, 108, 19222–19227. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cron, P.; Thompson, V.; Good, V.M.; Hess, D.; Hemmings, B.A.; Barford, D. Molecular Mechanism for the Regulation of Protein Kinase B/Akt by Hydrophobic Motif Phosphorylation. Mol. Cell 2002, 9, 1227–1240. [Google Scholar] [CrossRef]
- Chu, N.; Salguero, A.L.; Liu, A.Z.; Chen, Z.; Dempsey, D.R.; Ficarro, S.B.; Alexander, W.M.; Marto, J.A.; Li, Y.; Amzel, L.M.; et al. Akt Kinase Activation Mechanisms Revealed Using Protein Semisynthesis. Cell 2018, 174, 897–907.e14. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, R.; Péli-Gulli, M.P.; Hu, Z.; Jaquenoud, M.; Garcia Osuna, G.M.; Sardu, A.; Dengjel, J.; De Virgilio, C. Spatially Distinct Pools of TORC1 Balance Protein Homeostasis. Mol. Cell 2019, 73, 325–338.e8. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; Mao, K.; Jin, Y.; Tevzadze, G.; Kauffman, E.J.; Park, S.; Bridges, D.; Loewith, R.; Saltiel, A.R.; Klionsky, D.J.; et al. Roles for PI(3,5)P2 in Nutrient Sensing through TORC1. Mol. Biol. Cell 2014, 25, 1171–1185. [Google Scholar] [CrossRef]
- Lemmon, M.A. Phosphoinositide Recognition Domains. Traffic Cph. Den. 2003, 4, 201–213. [Google Scholar] [CrossRef]
- Posor, Y.; Jang, W.; Haucke, V. Phosphoinositides as Membrane Organizers. Nat. Rev. Mol. Cell Biol. 2022, 23, 797–816. [Google Scholar] [CrossRef]
- Lemmon, M.A. Membrane Recognition by Phospholipid-Binding Domains. Nat. Rev. Mol. Cell Biol. 2008, 9, 99–111. [Google Scholar] [CrossRef]
- Takeda, E.; Jin, N.; Itakura, E.; Kira, S.; Kamada, Y.; Weisman, L.S.; Noda, T.; Matsuura, A. Vacuole-Mediated Selective Regulation of TORC1-Sch9 Signaling Following Oxidative Stress. Mol. Biol. Cell 2018, 29, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Dove, S.K.; Cooke, F.T.; Douglas, M.R.; Sayers, L.G.; Parker, P.J.; Michell, R.H. Osmotic Stress Activates Phosphatidylinositol-3,5-Bisphosphate Synthesis. Nature 1997, 390, 187–192. [Google Scholar] [CrossRef]
- Jin, N.; Jin, Y.; Weisman, L.S. Early Protection to Stress Mediated by CDK-Dependent PI3,5P2 Signaling from the Vacuole/Lysosome. J. Cell Biol. 2017, 216, 2075–2090. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Clapp, K.; Tarsio, M.; Kane, P.M. Interaction of the Late Endo-Lysosomal Lipid PI(3,5)P2 with the Vph1 Isoform of Yeast V-ATPase Increases Its Activity and Cellular Stress Tolerance. J. Biol. Chem. 2019, 294, 9161–9171. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Kane, P.M. Regulation of V-ATPase Activity and Organelle PH by Phosphatidylinositol Phosphate Lipids. Front. Cell Dev. Biol. 2020, 8, 510. [Google Scholar] [CrossRef] [PubMed]
- Dechant, R.; Saad, S.; Ibáñez, A.J.; Peter, M. Cytosolic PH Regulates Cell Growth through Distinct Gtpases, Arf1 and Gtr1, to Promote Ras/PKA and TORC1 Activity. Mol. Cell 2014, 55, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechant, R.; Binda, M.; Lee, S.S.; Pelet, S.; Winderickx, J.; Peter, M. Cytosolic PH Is a Second Messenger for Glucose and Regulates the PKA Pathway through V-ATPase. EMBO J. 2010, 29, 2515–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laribee, R.N.; Hosni-Ahmed, A.; Workman, J.J.; Chen, H. Ccr4-Not Regulates RNA Polymerase I Transcription and Couples Nutrient Signaling to the Control of Ribosomal RNA Biogenesis. PLoS Genet. 2015, 11, e1005113. [Google Scholar] [CrossRef] [Green Version]
- Lenssen, E.; Oberholzer, U.; Labarre, J.; De Virgilio, C.; Collart, M.A. Saccharomyces Cerevisiae Ccr4-Not Complex Contributes to the Control of Msn2p-Dependent Transcription by the Ras/CAMP Pathway. Mol. Microbiol. 2002, 43, 1023–1037. [Google Scholar] [CrossRef] [Green Version]
- Laribee, R.N. Ccr4-Not as a Mediator of Environmental Signaling: A Jack of All Trades and Master of All. Curr. Genet. 2021, 67, 707–713. [Google Scholar] [CrossRef]
- Qie, B.; Lyu, Z.; Lyu, L.; Liu, J.; Gao, X.; Liu, Y.; Duan, W.; Zhang, N.; Du, L.; Liu, K. Sch9 Regulates Intracellular Protein Ubiquitination by Controlling Stress Responses. Redox Biol. 2015, 5, 290–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechant, R.; Peter, M. The N-Terminal Domain of the V-ATPase Subunit “a” Is Regulated by PH In Vitro and In Vivo. Channels 2011, 5, 4–8. [Google Scholar] [CrossRef]
- Saliba, E.; Evangelinos, M.; Gournas, C.; Corrillon, F.; Georis, I.; André, B. The Yeast H+-ATPase Pma1 Promotes Rag/Gtr-Dependent TORC1 Activation in Response to H+-Coupled Nutrient Uptake. eLife 2018, 7, e31981. [Google Scholar] [CrossRef]
- Ren, J.; Hannun, Y.A. Metabolism and Roles of Sphingolipids in Yeast Saccharomyces Cerevisiae. In Biogenesis of Fatty Acids, Lipids and Membranes; Geiger, O., Ed.; Handbook of Hydrocarbon and Lipid Microbiology; Springer: Cham, Switzerland, 2019; pp. 341–361. ISBN 978-3-319-43676-0. [Google Scholar] [CrossRef]
- Roelants, F.M.; Torrance, P.D.; Bezman, N.; Thorner, J. Pkh1 and Pkh2 Differentially Phosphorylate and Activate Ypk1 and Ykr2 and Define Protein Kinase Modules Required for Maintenance of Cell Wall Integrity. Mol. Biol. Cell 2002, 13, 3005–3028. [Google Scholar] [CrossRef] [Green Version]
- Swinnen, E.; Wilms, T.; Idkowiak-Baldys, J.; Smets, B.; De Snijder, P.; Accardo, S.; Ghillebert, R.; Thevissen, K.; Cammue, B.; De Vos, D.; et al. The Protein Kinase Sch9 Is a Key Regulator of Sphingolipid Metabolism in Saccharomyces Cerevisiae. Mol. Biol. Cell 2014, 25, 196–211. [Google Scholar] [CrossRef]
- Matmati, N.; Hannun, Y.A. Thematic Review Series: Sphingolipids. ISC1 (Inositol Phosphosphingolipid-Phospholipase C), the Yeast Homologue of Neutral Sphingomyelinases. J. Lipid Res. 2008, 49, 922–928. [Google Scholar] [CrossRef] [Green Version]
- Rego, A.; Cooper, K.F.; Snider, J.; Hannun, Y.A.; Costa, V.; Côrte-Real, M.; Chaves, S.R. Acetic Acid Induces Sch9p-Dependent Translocation of Isc1p from the Endoplasmic Reticulum into Mitochondria. Biochim. Biophys. Acta BBA-Mol. Cell Biol. Lipids 2018, 1863, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.C.; Sumanasekera, C.; Lester, R.L. Functions and Metabolism of Sphingolipids in Saccharomyces Cerevisiae. Prog. Lipid Res. 2006, 45, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, M.; Obara, K.; Kihara, A. Sphingolipid Synthesis Is Involved in Autophagy in Saccharomyces Cerevisiae. Biochem. Biophys. Res. Commun. 2011, 410, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Principles of Bioactive Lipid Signalling: Lessons from Sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Beeler, T.J.; Fu, D.; Rivera, J.; Monaghan, E.; Gable, K.; Dunn, T.M. SUR1 (CSG1/BCL21), a Gene Necessary for Growth of Saccharomyces Cerevisiae in the Presence of High Ca2+ Concentrations at 37 Degrees C, Is Required for Mannosylation of Inositolphosphorylceramide. Mol. Gen. Genet. MGG 1997, 255, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Uemura, S.; Kihara, A.; Iwaki, S.; Inokuchi, J.; Igarashi, Y. Regulation of the Transport and Protein Levels of the Inositol Phosphorylceramide Mannosyltransferases Csg1 and Csh1 by the Ca2+-Binding Protein Csg2. J. Biol. Chem. 2007, 282, 8613–8621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.D.; Osório, H.; Sims, K.J.; Almeida, T.; Alves, M.; Bielawski, J.; Amorim, M.A.; Moradas-Ferreira, P.; Hannun, Y.A.; Costa, V. Role for Sit4p-Dependent Mitochondrial Dysfunction in Mediating the Shortened Chronological Lifespan and Oxidative Stress Sensitivity of Isc1p-Deficient Cells. Mol. Microbiol. 2011, 81, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, V.; Medeiros, T.C.; Vilaça, R.; Pereira, A.T.; Chaves, S.R.; Côrte-Real, M.; Moradas-Ferreira, P.; Costa, V. Ceramide Signalling Impinges on Sit4p and Hog1p to Promote Mitochondrial Fission and Mitophagy in Isc1p-Deficient Cells. Cell. Signal. 2015, 27, 1840–1849. [Google Scholar] [CrossRef]
- Sun, Y.; Miao, Y.; Yamane, Y.; Zhang, C.; Shokat, K.M.; Takematsu, H.; Kozutsumi, Y.; Drubin, D.G. Orm Protein Phosphoregulation Mediates Transient Sphingolipid Biosynthesis Response to Heat Stress via the Pkh-Ypk and Cdc55-PP2A Pathways. Mol. Biol. Cell 2012, 23, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Muir, A.; Ramachandran, S.; Roelants, F.M.; Timmons, G.; Thorner, J. TORC2-Dependent Protein Kinase Ypk1 Phosphorylates Ceramide Synthase to Stimulate Synthesis of Complex Sphingolipids. eLife 2014, 3, e03779. [Google Scholar] [CrossRef]
- Iwaki, S.; Kihara, A.; Sano, T.; Igarashi, Y. Phosphorylation by Pho85 Cyclin-Dependent Kinase Acts as a Signal for the down-Regulation of the Yeast Sphingoid Long-Chain Base Kinase Lcb4 during the Stationary Phase. J. Biol. Chem. 2005, 280, 6520–6527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, J.A.; Estruch, F.; Córcoles-Sáez, I.; Del Poeta, M.; Rieger, R.; Stenzel, I.; Randez-Gil, F. Pho85 and PI(4,5)P2 Regulate Different Lipid Metabolic Pathways in Response to Cold. Biochim. Biophys. Acta BBA-Mol. Cell Biol. Lipids 2020, 1865, 158557. [Google Scholar] [CrossRef] [PubMed]
- Borklu Yucel, E.; Ulgen, K.O. Assessment of Crosstalks between the Snf1 Kinase Complex and Sphingolipid Metabolism in S. cerevisiae via Systems Biology Approaches. Mol. Biosyst. 2013, 9, 2914–2931. [Google Scholar] [CrossRef]
- Knupp, J.; Martinez-Montañés, F.; Van Den Bergh, F.; Cottier, S.; Schneiter, R.; Beard, D.; Chang, A. Sphingolipid Accumulation Causes Mitochondrial Dysregulation and Cell Death. Cell Death Differ. 2017, 24, 2044–2053. [Google Scholar] [CrossRef] [Green Version]
- Guerra, P.; Vuillemenot, L.-A.P.E.; van Oppen, Y.B.; Been, M.; Milias-Argeitis, A. TORC1 and PKA Activity towards Ribosome Biogenesis Oscillates in Synchrony with the Budding Yeast Cell Cycle. J. Cell Sci. 2022, 135, jcs260378. [Google Scholar] [CrossRef] [PubMed]
- Lippman, S.I.; Broach, J.R. Protein Kinase A and TORC1 Activate Genes for Ribosomal Biogenesis by Inactivating Repressors Encoded by Dot6 and Its Homolog Tod6. Proc. Natl. Acad. Sci. USA 2009, 106, 19928–19933. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Bodenmiller, B.; Uotila, A.; Stahl, M.; Wanka, S.; Gerrits, B.; Aebersold, R.; Loewith, R. Characterization of the Rapamycin-Sensitive Phosphoproteome Reveals That Sch9 Is a Central Coordinator of Protein Synthesis. Genes Dev. 2009, 23, 1929–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocklin, R.; Goebl, M. Nutrient Sensing Kinases PKA and Sch9 Phosphorylate the Catalytic Domain of the Ubiquitin-Conjugating Enzyme Cdc34. PLoS ONE 2011, 6, e27099. [Google Scholar] [CrossRef] [Green Version]
- Askree, S.H.; Yehuda, T.; Smolikov, S.; Gurevich, R.; Hawk, J.; Coker, C.; Krauskopf, A.; Kupiec, M.; McEachern, M.J. A Genome-Wide Screen for Saccharomyces Cerevisiae Deletion Mutants That Affect Telomere Length. Proc. Natl. Acad. Sci. USA 2004, 101, 8658–8663. [Google Scholar] [CrossRef]
- Connelly, C.J.; Vidal-Cardenas, S.; Goldsmith, S.; Greider, C.W. The Bur1 Cyclin-Dependent Kinase Regulates Telomere Length in Saccharomyces Cerevisiae. Yeast Chichester Engl. 2022, 39, 177–192. [Google Scholar] [CrossRef]
- Zaragoza, D.; Ghavidel, A.; Heitman, J.; Schultz, M.C. Rapamycin Induces the G0 Program of Transcriptional Repression in Yeast by Interfering with the TOR Signaling Pathway. Mol. Cell. Biol. 1998, 18, 4463–4470. [Google Scholar] [CrossRef] [Green Version]
- Sanz, P. Snf1 Protein Kinase: A Key Player in the Response to Cellular Stress in Yeast. Biochem. Soc. Trans. 2003, 31, 178–181. [Google Scholar] [CrossRef]
- Hughes Hallett, J.E.; Luo, X.; Capaldi, A.P. State Transitions in the TORC1 Signaling Pathway and Information Processing in Saccharomyces cerevisiae. Genetics 2014, 198, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Vaga, S.; Chumnanpuen, P.; Kumar, R.; Vemuri, G.N.; Aebersold, R.; Nielsen, J. Mapping the Interaction of Snf1 with TORC1 in Saccharomyces Cerevisiae. Mol. Syst. Biol. 2011, 7, 545. [Google Scholar] [CrossRef]
- Hughes Hallett, J.E.; Luo, X.; Capaldi, A.P. Snf1/AMPK Promotes the Formation of Kog1/Raptor-Bodies to Increase the Activation Threshold of TORC1 in Budding Yeast. eLife 2015, 4, e09181. [Google Scholar] [CrossRef] [PubMed]
- De Virgilio, C. The Essence of Yeast Quiescence. FEMS Microbiol. Rev. 2012, 36, 306–339. [Google Scholar] [CrossRef] [Green Version]
- Cameroni, E.; Hulo, N.; Roosen, J.; Winderickx, J.; De Virgilio, C. The Novel Yeast PAS Kinase Rim 15 Orchestrates G0-Associated Antioxidant Defense Mechanisms. Cell Cycle 2004, 3, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedruzzi, I.; Dubouloz, F.; Cameroni, E.; Wanke, V.; Roosen, J.; Winderickx, J.; De Virgilio, C. TOR and PKA Signaling Pathways Converge on the Protein Kinase Rim15 to Control Entry into G0. Mol. Cell 2003, 12, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Reinders, A.; Bürckert, N.; Boller, T.; Wiemken, A.; De Virgilio, C. Saccharomyces Cerevisiae CAMP-Dependent Protein Kinase Controls Entry into Stationary Phase through the Rim15p Protein Kinase. Genes Dev. 1998, 12, 2943–2955. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Shen, Y.; Gao, W.; Dong, J. Role of Sch9 in Regulating Ras-CAMP Signal Pathway in Saccharomyces Cerevisiae. FEBS Lett. 2011, 585, 3026–3032. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Gao, W. Mechanisms of Protein Kinase Sch9 Regulating Bcy1 in Saccharomyces Cerevisiae. FEMS Microbiol. Lett. 2012, 331, 10–16. [Google Scholar] [CrossRef]
- Sánchez-Adriá, I.E.; Sanmartín, G.; Prieto, J.A.; Estruch, F.; Randez-Gil, F. Slt2 Is Required to Activate ER-Stress-Protective Mechanisms through TORC1 Inhibition and Hexosamine Pathway Activation. J. Fungi 2022, 8, 92. [Google Scholar] [CrossRef]
- Lee, P.; Kim, M.S.; Paik, S.-M.; Choi, S.-H.; Cho, B.-R.; Hahn, J.-S. Rim15-Dependent Activation of Hsf1 and Msn2/4 Transcription Factors by Direct Phosphorylation in Saccharomyces Cerevisiae. FEBS Lett. 2013, 587, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Bontron, S.; Jaquenoud, M.; Vaga, S.; Talarek, N.; Bodenmiller, B.; Aebersold, R.; De Virgilio, C. Yeast Endosulfines Control Entry into Quiescence and Chronological Life Span by Inhibiting Protein Phosphatase 2A. Cell Rep. 2013, 3, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Dalgaard, J.Z.; Millar, J.B.A.; Arumugam, P. The Rim15-Endosulfine-PP2ACdc55 Signalling Module Regulates Entry into Gametogenesis and Quiescence via Distinct Mechanisms in Budding Yeast. PLoS Genet. 2014, 10, e1004456. [Google Scholar] [CrossRef] [PubMed]
- Talarek, N.; Bontron, S.; De Virgilio, C. Quantification of MRNA Stability of Stress-Responsive Yeast Genes Following Conditional Excision of Open Reading Frames. RNA Biol. 2013, 10, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Talarek, N.; Cameroni, E.; Jaquenoud, M.; Luo, X.; Bontron, S.; Lippman, S.; Devgan, G.; Snyder, M.; Broach, J.R.; De Virgilio, C. Initiation of the TORC1-Regulated G0 Program Requires Igo1/2, Which License Specific MRNAs to Evade Degradation via the 5’-3’ MRNA Decay Pathway. Mol. Cell 2010, 38, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swinnen, E.; Rosseels, J.; Winderickx, J. The Minimum Domain of Pho81 Is Not Sufficient to Control the Pho85-Rim15 Effector Branch Involved in Phosphate Starvation-Induced Stress Responses. Curr. Genet. 2005, 48, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Wanke, V.; Pedruzzi, I.; Cameroni, E.; Dubouloz, F.; De Virgilio, C. Regulation of G0 Entry by the Pho80-Pho85 Cyclin-CDK Complex. EMBO J. 2005, 24, 4271–4278. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Paik, S.-M.; Shin, C.-S.; Huh, W.-K.; Hahn, J.-S. Regulation of Yeast Yak1 Kinase by PKA and Autophosphorylation-Dependent 14-3-3 Binding. Mol. Microbiol. 2011, 79, 633–646. [Google Scholar] [CrossRef]
- Schmelzle, T.; Beck, T.; Martin, D.E.; Hall, M.N. Activation of the RAS/Cyclic AMP Pathway Suppresses a TOR Deficiency in Yeast. Mol. Cell. Biol. 2004, 24, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Cho, B.-R.; Joo, H.-S.; Hahn, J.-S. Yeast Yak1 Kinase, a Bridge between PKA and Stress-Responsive Transcription Factors, Hsf1 and Msn2/Msn4. Mol. Microbiol. 2008, 70, 882–895. [Google Scholar] [CrossRef]
- Zhang, N.; Quan, Z.; Rash, B.; Oliver, S.G. Synergistic Effects of TOR and Proteasome Pathways on the Yeast Transcriptome and Cell Growth. Open Biol. 2013, 3, 120137. [Google Scholar] [CrossRef] [Green Version]
- Moriya, H.; Shimizu-Yoshida, Y.; Omori, A.; Iwashita, S.; Katoh, M.; Sakai, A. Yak1p, a DYRK Family Kinase, Translocates to the Nucleus and Phosphorylates Yeast Pop2p in Response to a Glucose Signal. Genes Dev. 2001, 15, 1217–1228. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.E.; Soulard, A.; Hall, M.N. TOR Regulates Ribosomal Protein Gene Expression via PKA and the Forkhead Transcription Factor FHL1. Cell 2004, 119, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffioen, G.; Branduardi, P.; Ballarini, A.; Anghileri, P.; Norbeck, J.; Baroni, M.D.; Ruis, H. Nucleocytoplasmic Distribution of Budding Yeast Protein Kinase a Regulatory Subunit Bcy1 Requires Zds1 and Is Regulated by Yak1-Dependent Phosphorylation of Its Targeting Domain. Mol. Cell. Biol. 2001, 21, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Z.; Cao, L.; Tang, Y.; Yan, Y.; Oliver, S.G.; Zhang, N. The Yeast GSK-3 Homologue Mck1 Is a Key Controller of Quiescence Entry and Chronological Lifespan. PLoS Genet. 2015, 11, e1005282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffioen, G.; Swinnen, S.; Thevelein, J.M. Feedback Inhibition on Cell Wall Integrity Signaling by Zds1 Involves Gsk3 Phosphorylation of a CAMP-Dependent Protein Kinase Regulatory Subunit. J. Biol. Chem. 2003, 278, 23460–23471. [Google Scholar] [CrossRef] [Green Version]
- Mizunuma, M.; Hirata, D.; Miyaoka, R.; Miyakawa, T. GSK-3 Kinase Mck1 and Calcineurin Coordinately Mediate Hsl1 down-Regulation by Ca2+ in Budding Yeast. EMBO J. 2001, 20, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Tang, Y.; Quan, Z.; Zhang, Z.; Oliver, S.G.; Zhang, N. Chronological Lifespan in Yeast Is Dependent on the Accumulation of Storage Carbohydrates Mediated by Yak1, Mck1 and Rim15 Kinases. PLoS Genet. 2016, 12, e1006458. [Google Scholar] [CrossRef] [Green Version]
- Hirata, Y.; Andoh, T.; Asahara, T.; Kikuchi, A. Yeast Glycogen Synthase Kinase-3 Activates Msn2p-Dependent Transcription of Stress Responsive Genes. Mol. Biol. Cell 2003, 14, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Gutin, J.; Joseph-Strauss, D.; Sadeh, A.; Shalom, E.; Friedman, N. Genetic Screen of the Yeast Environmental Stress Response Dynamics Uncovers Distinct Regulatory Phases. Mol. Syst. Biol. 2019, 15, e8939. [Google Scholar] [CrossRef]
- Lee, J.; Moir, R.D.; McIntosh, K.B.; Willis, I.M. TOR Signaling Regulates Ribosome and TRNA Synthesis via LAMMER/Clk and GSK-3 Family Kinases. Mol. Cell 2012, 45, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Cao, L. Starvation Signals in Yeast Are Integrated to Coordinate Metabolic Reprogramming and Stress Response to Ensure Longevity. Curr. Genet. 2017, 63, 839–843. [Google Scholar] [CrossRef] [Green Version]
- Peterson, P.P.; Liu, Z. Identification and Characterization of Rapidly Accumulating Sch9Δ Suppressor Mutations in Saccharomyces Cerevisiae. G3 Genes 2021, 11, jkab134. [Google Scholar] [CrossRef]
- Balciunas, D.; Ronne, H. Yeast Genes GIS1-4: Multicopy Suppressors of the Gal- Phenotype of Snf1 Mig1 Srb8/10/11 Cells. Mol. Gen. Genet. MGG 1999, 262, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Estruch, F.; Carlson, M. Two Homologous Zinc Finger Genes Identified by Multicopy Suppression in a SNF1 Protein Kinase Mutant of Saccharomyces Cerevisiae. Mol. Cell. Biol. 1993, 13, 3872–3881. [Google Scholar] [CrossRef] [PubMed]
- De Wever, V.; Reiter, W.; Ballarini, A.; Ammerer, G.; Brocard, C. A Dual Role for PP1 in Shaping the Msn2-Dependent Transcriptional Response to Glucose Starvation. EMBO J. 2005, 24, 4115–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, J.-S.; Thiele, D.J. Activation of the Saccharomyces Cerevisiae Heat Shock Transcription Factor under Glucose Starvation Conditions by Snf1 Protein Kinase. J. Biol. Chem. 2004, 279, 5169–5176. [Google Scholar] [CrossRef] [Green Version]
- Mayordomo, I.; Estruch, F.; Sanz, P. Convergence of the Target of Rapamycin and the Snf1 Protein Kinase Pathways in the Regulation of the Subcellular Localization of Msn2, a Transcriptional Activator of STRE (Stress Response Element)-Regulated Genes. J. Biol. Chem. 2002, 277, 35650–35656. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, R.; Tripodi, F.; Gaggini, M.; Castoldi, A.; Reghellin, V.; Nonnis, S.; Tedeschi, G.; Coccetti, P. Snf1 Phosphorylates Adenylate Cyclase and Negatively Regulates Protein Kinase A-Dependent Transcription in Saccharomyces Cerevisiae. J. Biol. Chem. 2015, 290, 24715–24726. [Google Scholar] [CrossRef] [Green Version]
- Dauner, K.; Eid, W.; Raghupathy, R.; Presley, J.F.; Zha, X. MTOR Complex 1 Activity Is Required to Maintain the Canonical Endocytic Recycling Pathway against Lysosomal Delivery. J. Biol. Chem. 2017, 292, 5737–5747. [Google Scholar] [CrossRef] [Green Version]
- Dobzinski, N.; Chuartzman, S.G.; Kama, R.; Schuldiner, M.; Gerst, J.E. Starvation-Dependent Regulation of Golgi Quality Control Links the TOR Signaling and Vacuolar Protein Sorting Pathways. Cell Rep. 2015, 12, 1876–1886. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.B.; Ott, E.M.; Keener, J.M.; Curtiss, M.; Sandrin, V.; Babst, M. Regulation of Membrane Protein Degradation by Starvation-Response Pathways. Traffic 2012, 13, 468–482. [Google Scholar] [CrossRef] [Green Version]
- MacGurn, J.A.; Hsu, P.-C.; Smolka, M.B.; Emr, S.D. TORC1 Regulates Endocytosis via Npr1-Mediated Phosphoinhibition of a Ubiquitin Ligase Adaptor. Cell 2011, 147, 1104–1117. [Google Scholar] [CrossRef] [Green Version]
- Durchschlag, E.; Reiter, W.; Ammerer, G.; Schüller, C. Nuclear Localization Destabilizes the Stress-Regulated Transcription Factor Msn2. J. Biol. Chem. 2004, 279, 55425–55432. [Google Scholar] [CrossRef] [Green Version]
- Irniger, S.; Braus, G.H. Controlling Transcription by Destruction: The Regulation of Yeast Gcn4p Stability. Curr. Genet. 2003, 44, 8–18. [Google Scholar] [CrossRef]
- Zhang, N.; Oliver, S.G. The Transcription Activity of Gis1 Is Negatively Modulated by Proteasome-Mediated Limited Proteolysis. J. Biol. Chem. 2010, 285, 6465–6476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, A.; Bertolotti, A. Regulation of Proteasome Assembly and Activity in Health and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 697–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, A.; Bertolotti, A. An Evolutionarily Conserved Pathway Controls Proteasome Homeostasis. Nature 2016, 536, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.D.; Cacioppo, R.; Agrotis, A.; Black, A.; Zhou, H.; Rousseau, A. Actin Remodelling Controls Proteasome Homeostasis upon Stress. Nat. Cell Biol. 2022, 24, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.-S.; Neef, D.W.; Thiele, D.J. A Stress Regulatory Network for Co-Ordinated Activation of Proteasome Expression Mediated by Yeast Heat Shock Transcription Factor. Mol. Microbiol. 2006, 60, 240–251. [Google Scholar] [CrossRef]
- Laporte, D.; Salin, B.; Daignan-Fornier, B.; Sagot, I. Reversible Cytoplasmic Localization of the Proteasome in Quiescent Yeast Cells. J. Cell Biol. 2008, 181, 737–745. [Google Scholar] [CrossRef]
- Waite, K.A.; Roelofs, J. Proteasome Granule Formation Is Regulated through Mitochondrial Respiration and Kinase Signaling. J. Cell Sci. 2022, 135, jcs259778. [Google Scholar] [CrossRef]
- Karmon, O.; Ben Aroya, S. Spatial Organization of Proteasome Aggregates in the Regulation of Proteasome Homeostasis. Front. Mol. Biosci. 2019, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; McLoughlin, F.; Vierstra, R.D. Autophagic Turnover of Inactive 26S Proteasomes in Yeast Is Directed by the Ubiquitin Receptor Cue5 and the Hsp42 Chaperone. Cell Rep. 2016, 16, 1717–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waite, K.A.; Burris, A.; Vontz, G.; Lang, A.; Roelofs, J. Proteaphagy Is Specifically Regulated and Requires Factors Dispensable for General Autophagy. J. Biol. Chem. 2022, 298, 101494. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, R.; De Virgilio, C. A Spatially and Functionally Distinct Pool of TORC1 Defines Signaling Endosomes in Yeast. Autophagy 2019, 15, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Yoshino, K.-I.; Kondo, C.; Kawamata, T.; Oshiro, N.; Yonezawa, K.; Ohsumi, Y. Tor Directly Controls the Atg1 Kinase Complex to Regulate Autophagy. Mol. Cell. Biol. 2010, 30, 1049–1058. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, R.; De Virgilio, C. TORC1 Specifically Inhibits Microautophagy through ESCRT-0. Curr. Genet. 2019, 65, 1243–1249. [Google Scholar] [CrossRef] [Green Version]
- Morshed, S.; Sharmin, T.; Ushimaru, T. TORC1 Regulates ESCRT-0 Complex Formation on the Vacuolar Membrane and Microautophagy Induction in Yeast. Biochem. Biophys. Res. Commun. 2020, 522, 88–94. [Google Scholar] [CrossRef]
- Li, J.; Hochstrasser, M. Selective Microautophagy of Proteasomes Is Initiated by ESCRT-0 and Is Promoted by Proteasome Ubiquitylation. J. Cell Sci. 2022, 135, jcs259393. [Google Scholar] [CrossRef]
- Stephan, J.S.; Yeh, Y.-Y.; Ramachandran, V.; Deminoff, S.J.; Herman, P.K. The Tor and PKA Signaling Pathways Independently Target the Atg1/Atg13 Protein Kinase Complex to Control Autophagy. Proc. Natl. Acad. Sci. USA 2009, 106, 17049–17054. [Google Scholar] [CrossRef]
- Montella-Manuel, S.; Pujol-Carrion, N.; de la Torre-Ruiz, M.A. The Cell Wall Integrity Receptor Mtl1 Contributes to Articulate Autophagic Responses When Glucose Availability Is Compromised. J. Fungi 2021, 7, 903. [Google Scholar] [CrossRef]
- Yao, W.; Li, Y.; Chen, Y.; Chen, Y.; Zhao, P.; Zhang, Y.; Jiang, Q.; Feng, Y.; Yang, F.; Wu, C.; et al. Mec1 Regulates PAS Recruitment of Atg13 via Direct Binding with Atg13 during Glucose Starvation-Induced Autophagy. Proc. Natl. Acad. Sci. USA 2023, 120, e2215126120. [Google Scholar] [CrossRef]
- Yi, C.; Tong, J.; Lu, P.; Wang, Y.; Zhang, J.; Sun, C.; Yuan, K.; Xue, R.; Zou, B.; Li, N.; et al. Formation of a Snf1-Mec1-Atg1 Module on Mitochondria Governs Energy Deprivation-Induced Autophagy by Regulating Mitochondrial Respiration. Dev. Cell 2017, 41, 59–71.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartholomew, C.R.; Suzuki, T.; Du, Z.; Backues, S.K.; Jin, M.; Lynch-Day, M.A.; Umekawa, M.; Kamath, A.; Zhao, M.; Xie, Z.; et al. Ume6 Transcription Factor Is Part of a Signaling Cascade That Regulates Autophagy. Proc. Natl. Acad. Sci. USA 2012, 109, 11206–11210. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Jin, M.; González-Rodríguez, P.; Füllgrabe, J.; Delorme-Axford, E.; Backues, S.K.; Joseph, B.; Klionsky, D.J. Rph1/KDM4 Mediates Nutrient-Limitation Signaling That Leads to the Transcriptional Induction of Autophagy. Curr. Biol. 2015, 25, 546–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Huang, Y.; Wen, X.; Yin, Z.; Zhang, Z.; Klionsky, D.J. How Cells Deal with the Fluctuating Environment: Autophagy Regulation under Stress in Yeast and Mammalian Systems. Antioxidants 2022, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Vlahakis, A.; Lopez Muniozguren, N.; Powers, T. Stress-Response Transcription Factors Msn2 and Msn4 Couple TORC2-Ypk1 Signaling and Mitochondrial Respiration to ATG8 Gene Expression and Autophagy. Autophagy 2017, 13, 1804–1812. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, T.; Irie, K. Msn2/4 Transcription Factors Positively Regulate Expression of Atg39 ER-Phagy Receptor. Sci. Rep. 2021, 11, 11919. [Google Scholar] [CrossRef]
- DeMille, D.; Badal, B.D.; Evans, J.B.; Mathis, A.D.; Anderson, J.F.; Grose, J.H. PAS Kinase Is Activated by Direct SNF1-Dependent Phosphorylation and Mediates Inhibition of TORC1 through the Phosphorylation and Activation of Pbp1. Mol. Biol. Cell 2015, 26, 569–582. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Habernig, L.; Broeskamp, F.; Aufschnaiter, A.; Diessl, J.; Atienza, I.; Matz, S.; Ruiz, F.A.; Büttner, S. Phosphate Restriction Promotes Longevity via Activation of Autophagy and the Multivesicular Body Pathway. Cells 2021, 10, 3161. [Google Scholar] [CrossRef]
- Yang, Z.; Geng, J.; Yen, W.-L.; Wang, K.; Klionsky, D.J. Positive or Negative Roles of Different Cyclin-Dependent Kinase Pho85-Cyclin Complexes Orchestrate Induction of Autophagy in Saccharomyces Cerevisiae. Mol. Cell 2010, 38, 250–264. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological Aging-Independent Replicative Life Span Regulation by Msn2/Msn4 and Sod2 in Saccharomyces Cerevisiae. FEBS Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M.; Powers, R.W.; Steffen, K.K.; Westman, E.A.; Hu, D.; Dang, N.; Kerr, E.O.; Kirkland, K.T.; Fields, S.; Kennedy, B.K. Regulation of Yeast Replicative Life Span by TOR and Sch9 in Response to Nutrients. Science 2005, 310, 1193–1196. [Google Scholar] [CrossRef] [Green Version]
- Mirisola, M.G.; Taormina, G.; Fabrizio, P.; Wei, M.; Hu, J.; Longo, V.D. Serine- and Threonine/Valine-Dependent Activation of PDK and Tor Orthologs Converge on Sch9 to Promote Aging. PLoS Genet. 2014, 10, e1004113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, Z.; Gao, X.; Wang, W.; Dang, J.; Yang, L.; Yan, M.; Ali, S.A.; Liu, Y.; Liu, B.; Yu, M.; et al. MTORC1-Sch9 Regulates Hydrogen Sulfide Production through the Transsulfuration Pathway. Aging 2019, 11, 8418–8432. [Google Scholar] [CrossRef] [PubMed]
- Picazo, C.; Orozco, H.; Matallana, E.; Aranda, A. Interplay among Gcn5, Sch9 and Mitochondria during Chronological Aging of Wine Yeast Is Dependent on Growth Conditions. PLoS ONE 2015, 10, e0117267. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.; Leitão-Correia, F.; Sousa, M.J.; Leão, C. Ammonium Is a Key Determinant on the Dietary Restriction of Yeast Chronological Aging in Culture Medium. Oncotarget 2015, 6, 6511–6523. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liu, S.Q.; Huang, D. Dietary Restriction Depends on Nutrient Composition to Extend Chronological Lifespan in Budding Yeast Saccharomyces Cerevisiae. PLoS ONE 2013, 8, e64448. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Gattazzo, C.; Battistella, L.; Wei, M.; Cheng, C.; McGrew, K.; Longo, V.D. Sir2 Blocks Extreme Life-Span Extension. Cell 2005, 123, 655–667. [Google Scholar] [CrossRef]
- Kennedy, B.K.; Smith, E.D.; Kaeberlein, M. The Enigmatic Role of Sir2 in Aging. Cell 2005, 123, 548–550. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Finch, C.E. Evolutionary Medicine: From Dwarf Model Systems to Healthy Centenarians? Science 2003, 299, 1342–1346. [Google Scholar] [CrossRef] [Green Version]
- Pedruzzi, I.; Bürckert, N.; Egger, P.; De Virgilio, C. Saccharomyces Cerevisiae Ras/CAMP Pathway Controls Post-Diauxic Shift Element-Dependent Transcription through the Zinc Finger Protein Gis1. EMBO J. 2000, 19, 2569–2579. [Google Scholar] [CrossRef] [PubMed]
- Bitterman, K.J.; Medvedik, O.; Sinclair, D.A. Longevity Regulation in Saccharomyces Cerevisiae: Linking Metabolism, Genome Stability, and Heterochromatin. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 376–399. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Liou, L.-L.; Moy, V.N.; Diaspro, A.; Valentine, J.S.; Gralla, E.B.; Longo, V.D. SOD2 Functions Downstream of Sch9 to Extend Longevity in Yeast. Genetics 2003, 163, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Ahuir, A.; Proft, M. The Sch9 Kinase Is a Chromatin-Associated Transcriptional Activator of Osmostress-Responsive Genes. EMBO J. 2007, 26, 3098–3108. [Google Scholar] [CrossRef]
- Piper, P.W.; Harris, N.L.; MacLean, M. Preadaptation to Efficient Respiratory Maintenance Is Essential Both for Maximal Longevity and the Retention of Replicative Potential in Chronologically Ageing Yeast. Mech. Ageing Dev. 2006, 127, 733–740. [Google Scholar] [CrossRef]
- Bonawitz, N.D.; Chatenay-Lapointe, M.; Pan, Y.; Shadel, G.S. Reduced TOR Signaling Extends Chronological Life Span via Increased Respiration and Upregulation of Mitochondrial Gene Expression. Cell Metab. 2007, 5, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, H.; Whiteway, M. Increased Respiration in the Sch9Delta Mutant Is Required for Increasing Chronological Life Span but Not Replicative Life Span. Eukaryot. Cell 2008, 7, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowart, L.A.; Shotwell, M.; Worley, M.L.; Richards, A.J.; Montefusco, D.J.; Hannun, Y.A.; Lu, X. Revealing a Signaling Role of Phytosphingosine-1-Phosphate in Yeast. Mol. Syst. Biol. 2010, 6, 349. [Google Scholar] [CrossRef] [PubMed]
- Ludovico, P.; Burhans, W.C. Reactive Oxygen Species, Ageing and the Hormesis Police. FEMS Yeast Res. 2014, 14, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, A.; Weinberger, M.; Silva, A.; Sampaio-Marques, B.; Almeida, B.; Leão, C.; Costa, V.; Rodrigues, F.; Burhans, W.C.; Ludovico, P. Caloric Restriction or Catalase Inactivation Extends Yeast Chronological Lifespan by Inducing H2O2 and Superoxide Dismutase Activity. Proc. Natl. Acad. Sci. USA 2010, 107, 15123–15128. [Google Scholar] [CrossRef]
- Pan, Y. Mitochondria, Reactive Oxygen Species, and Chronological Aging: A Message from Yeast. Exp. Gerontol. 2011, 46, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Suganuma, T.; Gogol, M.M.; Workman, J.L. Histone H3 Threonine 11 Phosphorylation by Sch9 and CK2 Regulates Chronological Lifespan by Controlling the Nutritional Stress Response. eLife 2018, 7, e36157. [Google Scholar] [CrossRef] [PubMed]
- Madia, F.; Gattazzo, C.; Wei, M.; Fabrizio, P.; Burhans, W.C.; Weinberger, M.; Galbani, A.; Smith, J.R.; Nguyen, C.; Huey, S.; et al. Longevity Mutation in SCH9 Prevents Recombination Errors and Premature Genomic Instability in a Werner/Bloom Model System. J. Cell Biol. 2008, 180, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, P.; Battistella, L.; Vardavas, R.; Gattazzo, C.; Liou, L.-L.; Diaspro, A.; Dossen, J.W.; Gralla, E.B.; Longo, V.D. Superoxide Is a Mediator of an Altruistic Aging Program in Saccharomyces Cerevisiae. J. Cell Biol. 2004, 166, 1055–1067. [Google Scholar] [CrossRef]
- Moreno-Torres, M.; Jaquenoud, M.; De Virgilio, C. TORC1 Controls G1-S Cell Cycle Transition in Yeast via Mpk1 and the Greatwall Kinase Pathway. Nat. Commun. 2015, 6, 8256. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Torres, M.; Jaquenoud, M.; Péli-Gulli, M.-P.; Nicastro, R.; De Virgilio, C. TORC1 Coordinates the Conversion of Sic1 from a Target to an Inhibitor of Cyclin-CDK-Cks1. Cell Discov. 2017, 3, 17012. [Google Scholar] [CrossRef]
- Weinberger, M.; Mesquita, A.; Caroll, T.; Marks, L.; Yang, H.; Zhang, Z.; Ludovico, P.; Burhans, W.C. Growth Signaling Promotes Chronological Aging in Budding Yeast by Inducing Superoxide Anions That Inhibit Quiescence. Aging 2010, 2, 709–726. [Google Scholar] [CrossRef] [Green Version]
- Zinzalla, V.; Graziola, M.; Mastriani, A.; Vanoni, M.; Alberghina, L. Rapamycin-Mediated G1 Arrest Involves Regulation of the Cdk Inhibitor Sic1 in Saccharomyces Cerevisiae. Mol. Microbiol. 2007, 63, 1482–1494. [Google Scholar] [CrossRef]
- Hers, I.; Vincent, E.E.; Tavaré, J.M. Akt Signalling in Health and Disease. Cell. Signal. 2011, 23, 1515–1527. [Google Scholar] [CrossRef]
- Liko, D.; Hall, M.N. MTOR in Health and in Sickness. J. Mol. Med. 2015, 93, 1061–1073. [Google Scholar] [CrossRef]
- Carling, D. AMPK Signalling in Health and Disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kitano, H. Biological Robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, T.; Mayo, A.E.; Tlusty, T.; Alon, U. Evolution of Bow-Tie Architectures in Biology. PLoS Comput. Biol. 2015, 11, e1004055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, T.; Graumann, J.; Bryan, J.; MacCoss, M.J.; Deshaies, R.J. Quantitative Profiling of Ubiquitylated Proteins Reveals Proteasome Substrates and the Substrate Repertoire Influenced by the Rpn10 Receptor Pathway. Mol. Cell. Proteom. MCP 2007, 6, 1885–1895. [Google Scholar] [CrossRef] [Green Version]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villén, J. Global Analysis of Phosphorylation and Ubiquitylation Cross-Talk in Protein Degradation. Nat. Methods 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.; McGraw, T.E. The Akt Kinases: Isoform Specificity in Metabolism and Cancer. Cell Cycle 2009, 8, 2502–2508. [Google Scholar] [CrossRef]
- Bos, J.L. Molecular Mechanisms of Signal Transduction; IOS Press: Amsterdam, The Netherlands, 2000; ISBN 978-1-58603-016-2. [Google Scholar]
- González, A.; Shimobayashi, M.; Eisenberg, T.; Merle, D.A.; Pendl, T.; Hall, M.N.; Moustafa, T. TORC1 Promotes Phosphorylation of Ribosomal Protein S6 via the AGC Kinase Ypk3 in Saccharomyces Cerevisiae. PLoS ONE 2015, 10, e0120250. [Google Scholar] [CrossRef] [Green Version]
- Yerlikaya, S.; Meusburger, M.; Kumari, R.; Huber, A.; Anrather, D.; Costanzo, M.; Boone, C.; Ammerer, G.; Baranov, P.V.; Loewith, R. TORC1 and TORC2 Work Together to Regulate Ribosomal Protein S6 Phosphorylation in Saccharomyces Cerevisiae. Mol. Biol. Cell 2016, 27, 397–409. [Google Scholar] [CrossRef]
- Odorizzi, G.; Babst, M.; Emr, S.D. Phosphoinositide Signaling and the Regulation of Membrane Trafficking in Yeast. Trends Biochem. Sci. 2000, 25, 229–235. [Google Scholar] [CrossRef]
- Fu, W.; Hall, M.N. Regulation of MTORC2 Signaling. Genes 2020, 11, 1045. [Google Scholar] [CrossRef]



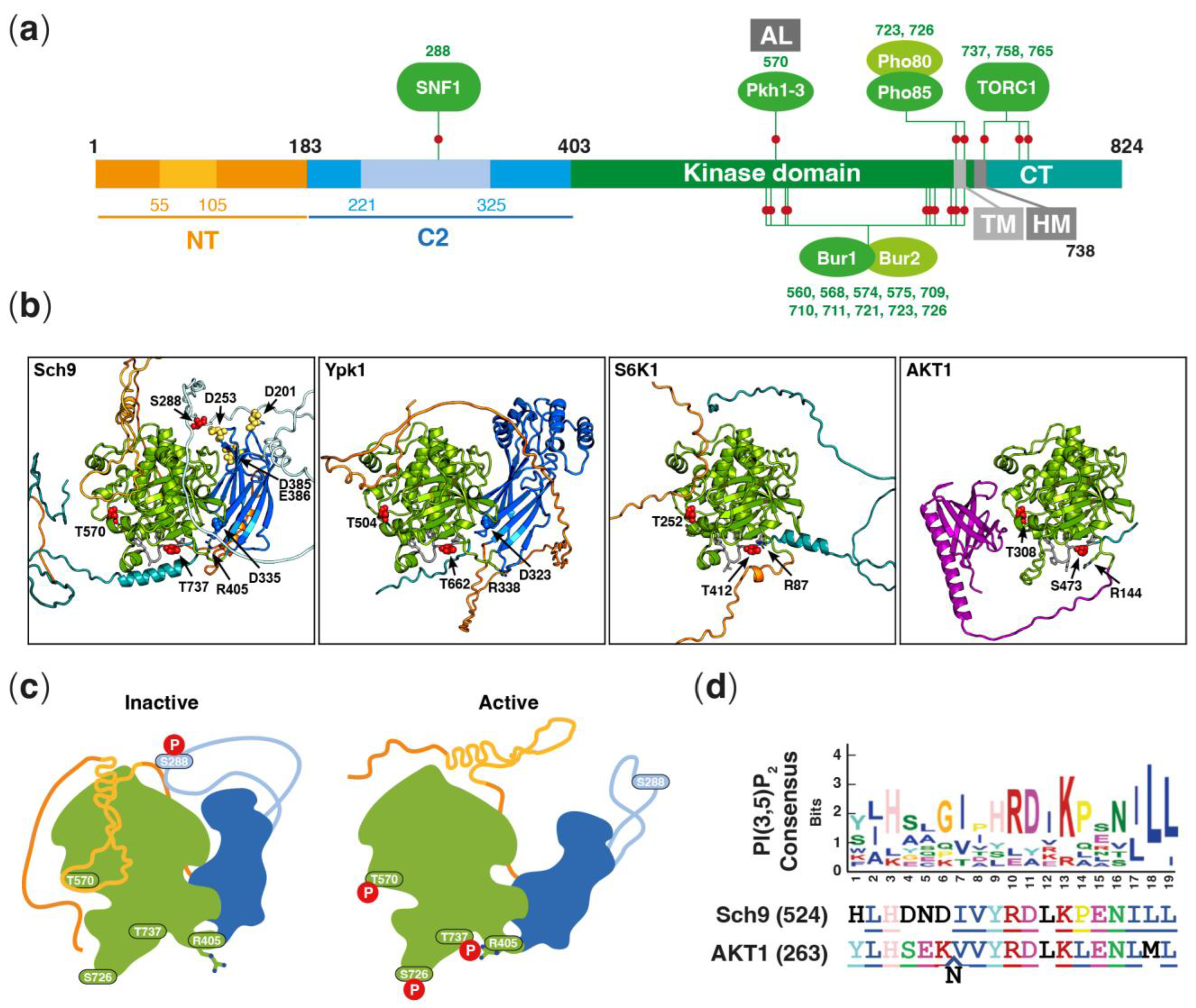{kind=link}
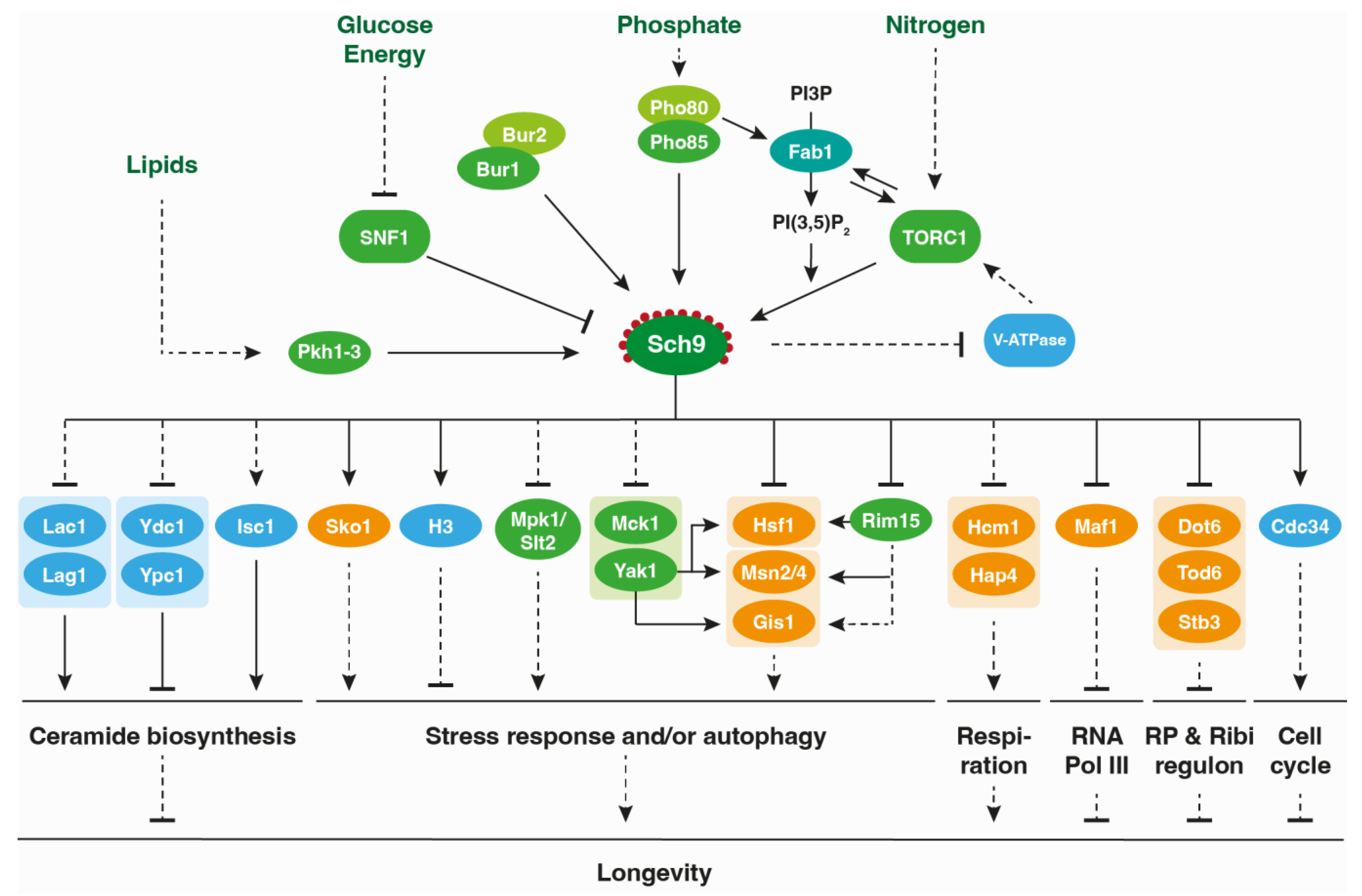{kind=link}
| Yeast | Mammalian | |||
|---|---|---|---|---|
| Region | Sch9 | Ypk1 | S6K1 | AKT1 |
| N-terminus | 1–183 | 1–117 | 1–83 | See PH domain |
| PH + Linker | - | - | - | 1–141 |
| C2 domain | 184–402 | 118–336 | - | - |
| C2 extended loop | 221–325 | - | - | - |
| Kinase domain | 403–738 | 337–663 | 84–413 | 142–474 |
| Activation loop | 570–574 (TFCGT) | 504–508 (TFCGT) | 252–256 (TFCGT) | 308–312 (TFCGT) |
| Hydrophobic motif | 733–738 (FAGFTF) | 658–663 (FGGWTY) | 408–413 (FLGFTY) | 469–474 (FPQFSY) |
| C-terminus | 739–824 | 664–680 | 414–525 | 475–480 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caligaris, M.; Sampaio-Marques, B.; Hatakeyama, R.; Pillet, B.; Ludovico, P.; De Virgilio, C.; Winderickx, J.; Nicastro, R. The Yeast Protein Kinase Sch9 Functions as a Central Nutrient-Responsive Hub That Calibrates Metabolic and Stress-Related Responses. J. Fungi 2023, 9, 787. https://doi.org/10.3390/jof9080787
Caligaris M, Sampaio-Marques B, Hatakeyama R, Pillet B, Ludovico P, De Virgilio C, Winderickx J, Nicastro R. The Yeast Protein Kinase Sch9 Functions as a Central Nutrient-Responsive Hub That Calibrates Metabolic and Stress-Related Responses. Journal of Fungi. 2023; 9(8):787. https://doi.org/10.3390/jof9080787
Chicago/Turabian StyleCaligaris, Marco, Belém Sampaio-Marques, Riko Hatakeyama, Benjamin Pillet, Paula Ludovico, Claudio De Virgilio, Joris Winderickx, and Raffaele Nicastro. 2023. "The Yeast Protein Kinase Sch9 Functions as a Central Nutrient-Responsive Hub That Calibrates Metabolic and Stress-Related Responses" Journal of Fungi 9, no. 8: 787. https://doi.org/10.3390/jof9080787