

Host Range and Loop-Mediated Isothermal Amplification Detection of Globisporangium sylvaticum from Guizhou, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Collection, Isolation, and Conservation
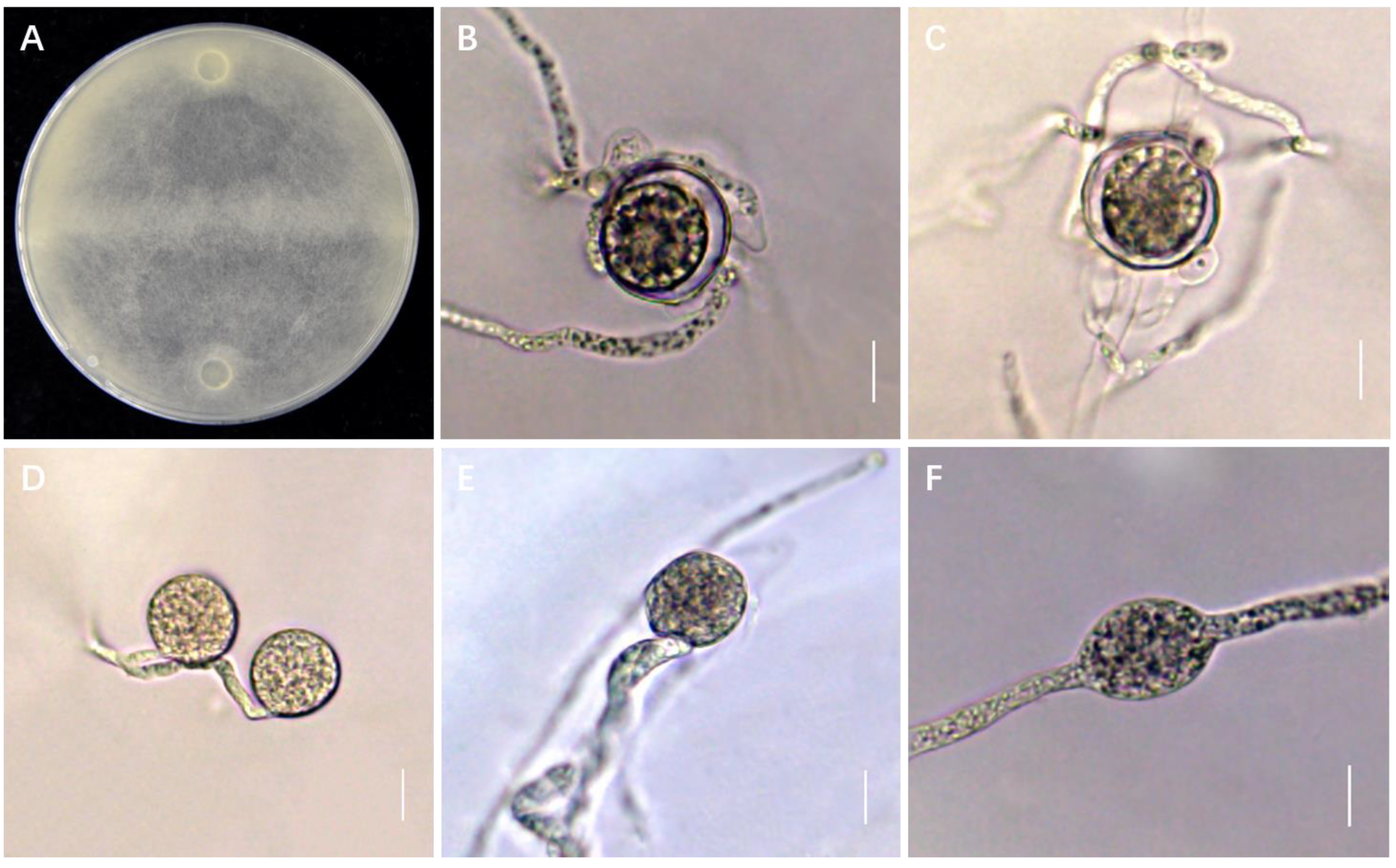
2.3. Morphological Studies
2.4. DNA Extraction, PCR Analysis, and Multi-Locus Phylogeny
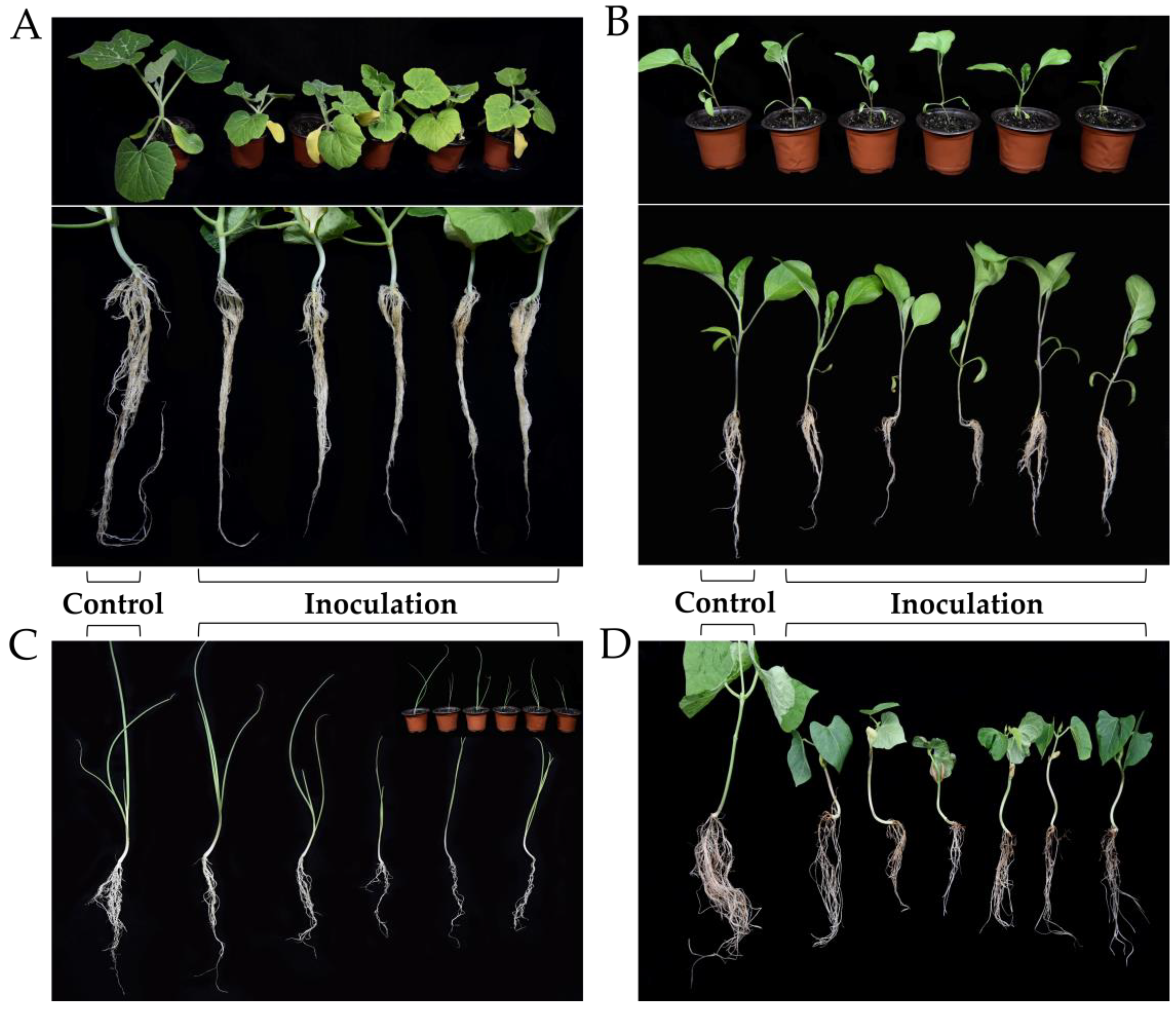
2.5. Koch’s Postulates
2.6. LAMP and PCR Primer Design
2.7. LAMP Reaction
2.8. Specificity and Sensitivity of LAMP or PCR Primers in Detecting G. sylvaticum
2.9. Detection of G. sylvaticum in Plant Roots
3. Results
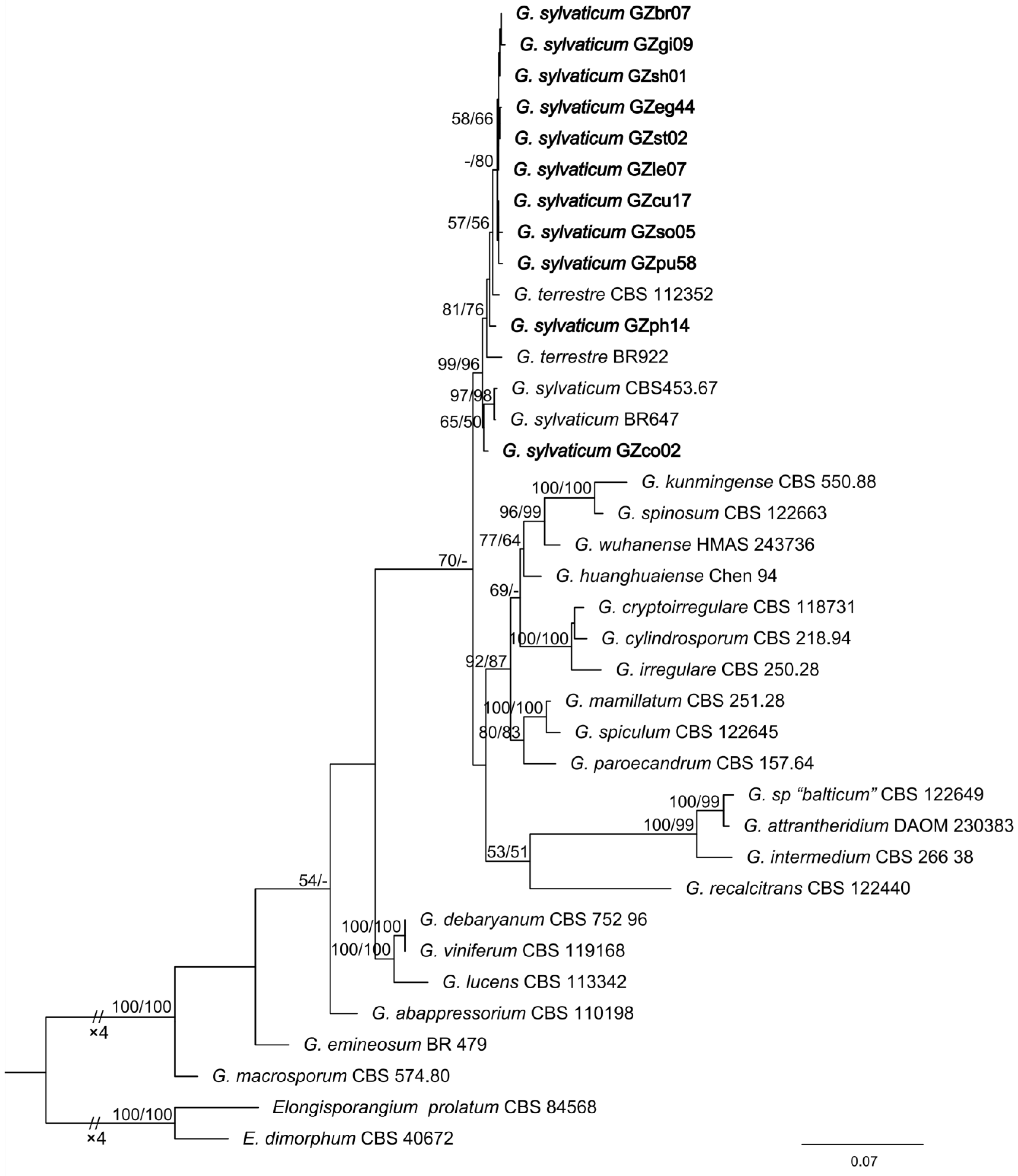
3.1. Identification of G. sylvaticum
3.2. Host Plants of G. sylvaticum in Guizhou, China
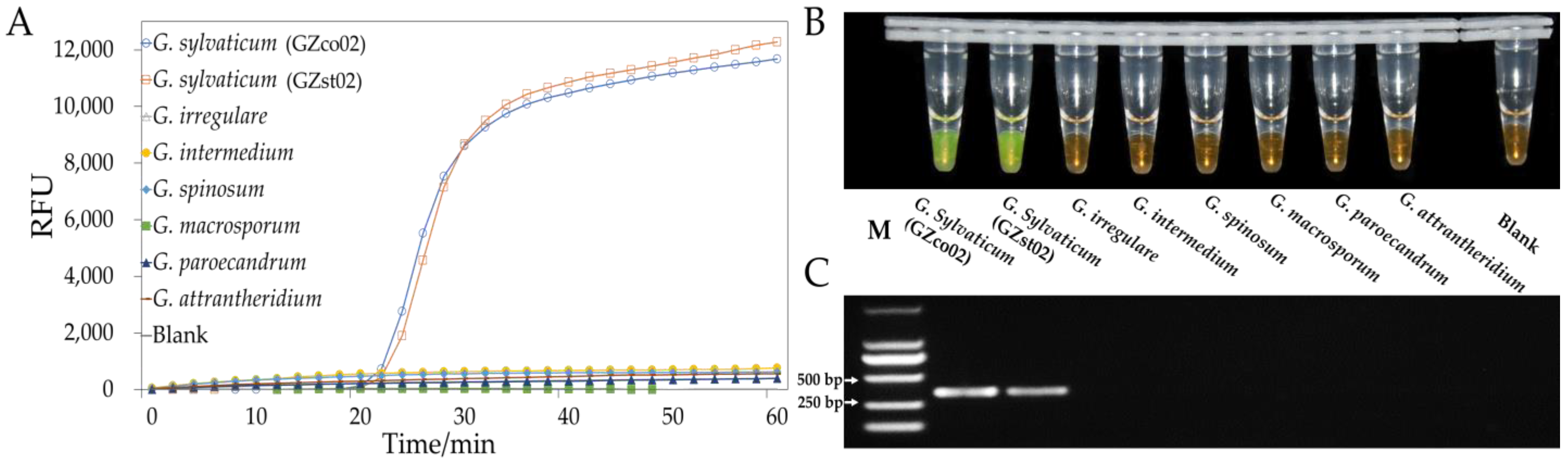
3.3. LAMP and PCR Primer Design and Specificity
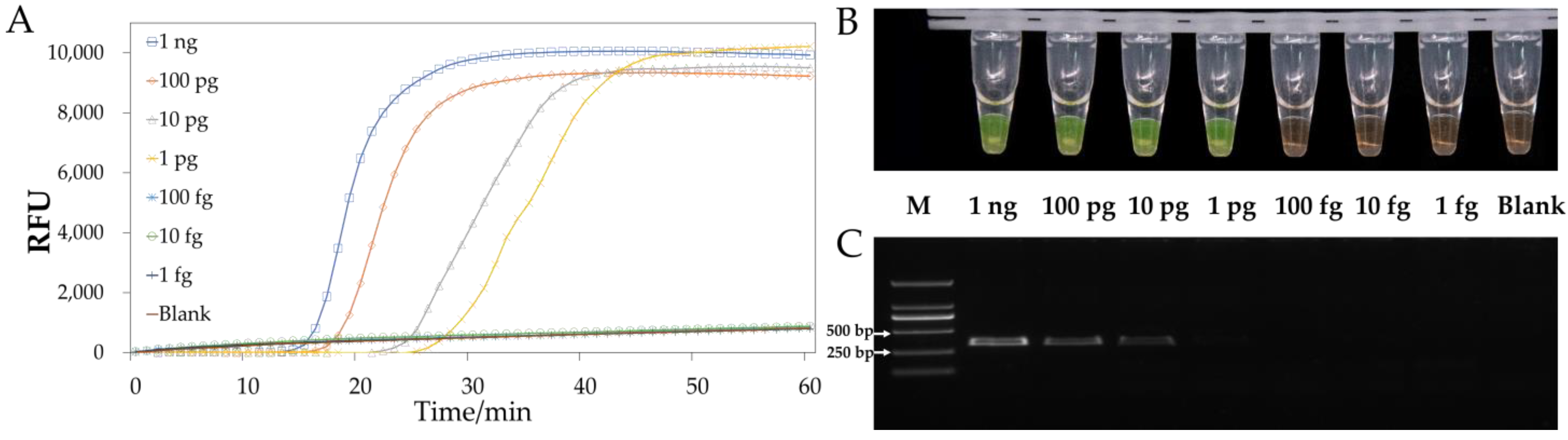
3.4. Sensitivity of LAMP or PCR In Vitro
3.5. Detection of the Pathogen in Field Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uzuhashi, S.; Kakishima, M.; Tojo, M. Phylogeny of the genus Pythium and description of new genera. Mycoscience 2010, 51, 337–365. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Dodge, A.; Dadej, K.; Rintoul, T.L.; Ponomareva, E.; Martin, F.N.; de Cock, A.W.A.M.; Lévesque, C.A.; Redhead, S.A.; Spies, C.F. Whole genome sequencing and phylogenomic analysis show support for the splitting of genus Pythium. Mycologia 2022, 114, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Abd-Elsalam, K.A.; Ingle, A.P.; Paralikar, P.; Ingle, P. The genus Pythium: An overview. In Pythium; CRC Press: Boca Raton, FL, USA, 2020; pp. 3–14. [Google Scholar]
- Parveen, T.; Sharma, K. Pythium diseases, control and management strategies: A review. Int. J. Plant Anim. Environ. Sci. 2015, 5, 244–257. [Google Scholar]
- Qian, C.; Qiang, H.; Zhang, G.; Li, M. Long-term changes of forest biomass and its driving factors in karst area, Guizhou, China. Int. J. Distrib. Sens. Netw. 2021, 17. [Google Scholar] [CrossRef]
- Ho, H.H.; Abd-Elsalam, K.A. Pathogenic and beneficial Pythium species in China: An updated review. In Pythium; CRC Press: Boca Raton, FL, USA, 2020; pp. 107–122. [Google Scholar]
- Campbell, W.A.; Hendrix, F.F., Jr. A new heterothallic Pythium from southern United States. Mycologia 1967, 59, 274–278. [Google Scholar] [CrossRef]
- Lin, F.; Wani, S.H.; Collins, P.J.; Wen, Z.; Li, W.; Zhang, N.; McCoy, A.G.; Bi, Y.; Tan, R.; Zhang, S.; et al. QTL mapping and GWAS for identification of loci conferring partial resistance to Pythium sylvaticum in soybean (Glycine max (L.) Merr). Mol. Breeding 2020, 40, 1–11. [Google Scholar] [CrossRef]
- Magnée, K.J.; Postma, J.; Groot, S.P.; Gort, G.; Lammerts van Bueren, E.T.; Scholten, O.E. Evaluation of damping-off tolerance in spinach cultivars in field soils and in a standardized lab assay with Pythium ultimum. Plant Health Prog. 2022, 23, 174–187. [Google Scholar] [CrossRef]
- Alejandro Rojas, J.; Jacobs, J.L.; Napieralski, S.; Karaj, B.; Bradley, C.A.; Chase, T.; Esker, P.D.; Giesler, L.J.; Jardine, D.J.; Malvick, D.K.; et al. Oomycete species associated with soybean seedlings in North America—Part I: Identification and pathogenicity characterization. Phytopathology 2017, 107, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Chen, S.; Yan, X.; Yuan, H.; Yang, D. First report of Pythium sylvaticum causing corn root rot in Northeastern China. Plant Dis. 2021, 105, 231. [Google Scholar] [CrossRef]
- Gilardi, G.; Matic, S.; Gullino, M.L.; Garibaldi, A. First report of root rot caused by Pythium sylvaticum on lettuce in Italy. Plant Dis. 2018, 102, 454–455. [Google Scholar] [CrossRef]
- Ahonsi, M.O.; Agindotan, B.O.; Gray, M.E.; Bradley, C.A. First report of basal stem rot and foliar blight caused by Pythium sylvaticum on Miscanthus sinensis in Illinois. Plant Dis. 2011, 95, 616. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.D.; Platt, H.W.B.; Lévesque, C.A. First report of Pythium sylvaticum causing potato tuber rot. Am. J. Potato Res. 2005, 82, 173–177. [Google Scholar] [CrossRef]
- Kageyama, K. Molecular taxonomy and its application to ecological studies of Pythium species. J. Gen. Plant Pathol. 2014, 80, 314–326. [Google Scholar] [CrossRef]
- Van der Heyden, H.; Wallon, T.; Lévesque, C.A.; Carisse, O. Detection and quantification of Pythium tracheiphilum in soil by multiplex real-time qPCR. Plant Dis. 2019, 103, 475–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushford, C.A.; North, R.L.; Miller, G.L. Detection of Pythium spp. in Golf Course Irrigation Systems. Plant Dis. 2022, 106, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Binggan, L.; Bingxin, Z. Identification of Pythium sylvaticum and species-specific primers. Mycosystema 2004, 23, 356–365. [Google Scholar]
- Patel, R.; Mitra, B.; Vinchurkar, M.; Adami, A.; Patkar, R.; Giacomozzi, F.; Lorenzelli, L.; Baghini, M.S. Plant pathogenicity and associated/related detection systems. A review. Talanta 2022, 251, 123808. [Google Scholar] [CrossRef]
- Jeevalatha, A.; Zumaila, F.; Biju, C.N.; Punya, K.C. Duplex recombinase polymerase amplification assay for simultaneous detection of Pythium spp. and Ralstonia pseudosolanacearum from ginger rhizomes. Crop Protect. 2022, 161, 106057. [Google Scholar] [CrossRef]
- Feng, H.; Ye, W.; Liu, Z.; Wang, Y.; Chen, J.; Wang, Y.; Zheng, X. Development of LAMP Assays Using a Novel Target Gene for Specific Detection of Pythium terrestris, Pythium spinosum, and ‘Candidatus Pythium huanghuaiense’. Plant Dis. 2021, 105, 2888–2897. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Hieno, A.; Kusunoki, M.; Suga, H.; Kageyama, K. LAMP detection of four plant-pathogenic oomycetes and its application in lettuce fields. Plant Dis. 2019, 103, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zou, H.; Zhang, J.; Ding, H.; Li, C.; Chen, X.; Li, Y.; Feng, W.; Kageyama, K. High-efficiency and high-fidelity ssDNA circularisation via the pairing of five 3′-terminal bases to assist LR-LAMP for the genotyping of single-nucleotide polymorphisms. Analyst 2022, 147, 3993–3999. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Chen, R.; Chen, G.; Li, M.; Wang, J.; Zou, J.; Du, F.; Dong, J.; Cui, X.; Huang, X.; et al. One-step colorimetric genotyping of single nucleotide polymorphism using probe-enhanced loop-mediated isothermal amplification (PE-LAMP). Theranostics 2019, 9, 3723. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-mediated isothermal amplification (LAMP): A versatile technique for detection of micro-organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Ji, H.; Yu, Y.; Wang, X.; Cheng, Y.; Li, Z.; Chen, J.; Guo, L.; Xu, J.; Gao, C. Development of a loop-mediated isothermal amplification method for the rapid detection of phytopythium vexans. Front. Microbiol. 2021, 12, 720485. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tojo, M. Modifications of PARP medium using fluazinam, miconazole, and nystatin for detection of Pythium spp. in soil. Plant Dis. 2007, 91, 1591–1599. [Google Scholar] [CrossRef] [Green Version]
- Van der Plaats-Niterink, A.J. Monograph of the genus Pythium. In Studies in Mycology; Centraalbureau voor Schimmelcultures (Baarn): Baarn, The Netherlands, 1981; Volume 21. [Google Scholar]
- Baten, M.A.; Asano, T.; Motohashi, K.; Ishiguro, Y.; Rahman, M.Z.; Inaba, S.; Suga, H.; Kageyama, K. Phylogenetic relationships among Phytopythium species, and re-evaluation of Phytopythium fagopyri comb. nov., recovered from damped-off buckwheat seedlings in Japan. Mycol. Prog. 2014, 13, 1145–1156. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Robideau, G.P.; de Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Desaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef]
- Feng, W.; Ishiguro, Y.; Hotta, K.; Watanabe, H.; Suga, H.; Kageyama, K. Simple detection of Pythium irregulare using loop-mediated isothermal amplification assay. FEMS Microbiol. Lett. 2015, 362, fnv174. [Google Scholar] [CrossRef] [Green Version]
- Moein, S.; Mazzola, M.; Spies, C.F.J.; McLeod, A. Evaluating different approaches for the quantification of oomycete apple replant pathogens, and their relationship with seedling growth reductions. Eur. J. Plant Pathol. 2019, 154, 243–257. [Google Scholar] [CrossRef]
- Matthiesen, R.L.; Ahmad, A.A.; Robertson, A.E. Temperature affects aggressiveness and fungicide sensitivity of four Pythium spp. that cause soybean and corn damping off in Iowa. Plant Dis. 2016, 100, 583–591. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, U.; Dee, M.M.; Ownley, B.H.; Butler, D.M. First Report of Strawberry Black Root Rot Caused by Globisporangium sylvaticum in Tennessee, USA. Plant Dis. 2023. [CrossRef]
- Paul, B. Pythium terrestris, a new species isolated from France, its ITS region, taxonomy and its comparison with related species. FEMS Microbiol. Lett. 2002, 212, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jambhulkar, P.P.; Raja, M.; Javeria, S. Pythium spp. on vegetable crops: Research progress and major challenges. In Pythium; CRC Press: Boca Raton, FL, USA, 2020; pp. 136–161. [Google Scholar]
- Lévesque, C.A.; Beckenbach, K.; Baillie, D.L.; Rahe, J.E. Pathogenicity and DNA restriction fragment length polymorphisms of isolates of Pythium spp. from glyphosatetreated seedlings. Mycol. Res. 1993, 97, 307–312. [Google Scholar] [CrossRef]
- Feng, H.; Chen, J.; Yu, Z.; Li, K.; Li, Z.; Li, Y.; Wang, Y.; Zheng, X. Pathogenicity and fungicide sensitivity of Pythium and Phytopythium spp. associated with soybean in the Huang-Huai region of China. Plant Pathol. 2020, 69, 1083–1092. [Google Scholar] [CrossRef]
- Bao, Y.; Jiang, Y.; Xiong, E.; Tian, T.; Zhang, Z.; Lv, J.; Li, Y.; Zhou, X. CUT-LAMP: Contamination-free loop-mediated isothermal amplification based on the CRISPR/Cas9 cleavage. ACS Sens. 2020, 5, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liang, M.; Lin, T.; Zhao, Q.; Wang, H.; Yang, S.; Guo, Q.; Wang, X.; Guo, H.; Cui, L.; et al. A LAMP-based toolbox developed for detecting the major pathogens affecting the production and quality of the Chinese medicinal crop Aconitum carmichaelii. Plant Dis. 2023, 107, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, K.; Zhang, H.; Yang, D.; Deng, Y.; Wang, Y.; Qi, Z. Development of a LAMP method for detecting F129L mutant in azoxystrobin-resistant Pyricularia oryzae. Fungal Biol. 2022, 126, 47–53. [Google Scholar] [CrossRef]
- Takahashi, R.; Fukuta, S.; Kuroyanagi, S.; Miyake, N.; Nagai, H.; Kageyama, K.; Ishiguro, Y. Development and application of a loop-mediated isothermal amplification assay for rapid detection of Pythium helicoides. FEMS Microbiol. Lett. 2014, 355, 28–35. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T.J.M.C.P. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Feng, W.; Otsubo, K.; Hieno, A.; Suga, H.; Kageyama, K. A simple loop-mediated isothermal amplification assay to detect Phytophthora colocasiae in infected taro plants. J. Gen. Plant Pathol. 2019, 85, 337–346. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, R.; Xu, C.; Liu, C.; Zheng, Y.; Zhang, X.; Liu, S.; Li, Y. Characterisation of Pythium aristosporum Oomycete—A Novel Pathogen Causing Rice Seedling Blight in China. J. Fungi 2022, 8, 890. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.S.; Zhang, B.; Ma, L.G.; Ma, G.P.; Qin, S.J.; Li, C.S.; Xu, Z.T.; Qi, K.; Zhang, Y.L. First report of Pythium aphanidermatum causing root rot of head lettuce in China. Plant Dis. 2021, 105, 3767. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, B.; Sun, X.; Qi, X.; Zhao, C.; Chang, X.; Khaskheli, M.I.; Gong, G. Symptoms and pathogens diversity of Corn Fusarium sheath rot in Sichuan Province, China. Sci. Rep. 2021, 11, 2835. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Isolates | Locality | GenBank Accession No. | |
|---|---|---|---|---|
| ITS | COI | |||
| Globisporangium abappressorium | CBS 110198 | USA | HQ643408 | HQ708455 |
| G. cryptoirregulare | CBS 118731 | USA | HQ643515 | HQ708561 |
| G. cylindrosporum | CBS 218.94 | Germany | HQ643516 | HQ708562 |
| G. debaryanum | CBS 752.96 | UK | HQ643519 | HQ708565 |
| G. emineosum | BR 479 | UK | GQ244427 | GQ244423 |
| G. irregulare | CBS 250.28 | Netherlands | HQ643596 | HQ708640 |
| G. lucens | CBS 113342 | UK | HQ643681 | HQ708725 |
| G. mamillatum | CBS 251.28 | Netherlands | HQ643687 | HQ708731 |
| G. paroecandrum | CBS 157.64 | Australia | HQ643731 | HQ708772 |
| G. recalcitrans | CBS 122440 | Spain | DQ357833 | EF426549 |
| G. spiculum | CBS 122645 | France | HQ643790 | HQ708831 |
| G. spinosum | CBS 122663 | India | HQ643791 | HQ708832 |
| G. sylvaticum | CBS 453.67 | USA | HQ643845 | HQ708886 |
| G. sylvaticum | BR647 | Netherlands | HQ643847 | HQ708888 |
| G. terrestre | CBS 112352 | France | HQ643857 | HQ708898 |
| G. terrestre | BR922 | USA | HQ643856 | HQ708897 |
| G. viniferum | CBS 119168 | France | HQ643956 | HQ708997 |
| G. sylvaticum | GZsh01 | China | OQ654058 | OQ694389 |
| G. sylvaticum | GZph14 | China | OQ654059 | OQ694390 |
| G. sylvaticum | GZco02 | China | OQ654060 | OQ694391 |
| G. sylvaticum | GZst02 | China | OQ654061 | OQ694392 |
| G. sylvaticum | GZso05 | China | OQ654062 | OQ694393 |
| G. sylvaticum | GZle07 | China | OQ654063 | OQ694394 |
| G. sylvaticum | GZbr07 | China | OQ654064 | OQ694395 |
| G. sylvaticum | GZpu58 | China | OQ654065 | OQ694396 |
| G. sylvaticum | GZcu17 | China | OQ654066 | OQ694397 |
| G. sylvaticum | GZgi09 | China | OQ654067 | OQ694398 |
| G. sylvaticum | GZeg44 | China | OQ654068 | OQ694399 |
| Elongisporangium dimorphum | CBS 40672 | USA | HQ643525 | HQ708571 |
| E. prolatum | CBS 84568 | USA | HQ643754 | HQ708795 |
| Species | Clade | Isolates a | Origin | Detection | |
|---|---|---|---|---|---|
| LAMP | PCR | ||||
| Pythium aphanidermatum | A | GZHca2 | Capsicum annuum L. | − | − |
| P. giumdeliense | A | GZHs21 | Soil | − | − |
| P. aristosporum | B | GZWco5 | Corns (Zea mays L.) | − | − |
| P. aquatile | B | GZal1 | Allium tuberosum Rottler ex Sprengle | − | − |
| P. deliense | B | GZAbr6 | Brassica rapa var. glabra Regel | − | − |
| P. dissotocum | B | GZbr24 | Brassica rapa var. glabra Regel | − | − |
| P. inflatum | B | GZHs55 | Soil | − | − |
| P. torulosum | B | GZHs12 | Soil | − | − |
| P. oligandrum | D | GZHs172 | Soil | − | − |
| Globisporangium hypogynum | E | GZbr2 | Brassica napus L. | − | − |
| G. middletonii | E | GZHs43 | Water | − | − |
| G. attrantheridium | F | GZLra1 | Raphanus sativus L. | − | − |
| G. intermedium | F | GZbr1 | Brassica rapa var. chinensis (Linnaeus) Kitamura | − | − |
| G. irregulare | F | GZvi11 | Vigna unguiculata (Linn.) Walp. | − | − |
| G. irregulare | F | GZLca2 | Capsicum annuum L. | − | − |
| G. macrosporum | F | GZHZgl3 | Glycine max (Linn.) Merr. | − | − |
| G. paroecandrum | F | GZHco1 | Coriandrum sativum L. | − | − |
| G. spinosum | F | GZbc1 | Brassica chinensis L. | − | − |
| G. spinosum | F | GZvi1 | Vicia faba L. | − | − |
| G. sylvaticum | F | GZsh01 | Shallots (Allium schoenoprasum L.) | + | + |
| G. sylvaticum | F | GZph14 | Green beans (Phaseolus vulgaris L.) | + | + |
| G. sylvaticum | F | GZco02 | Corns (Zea mays L.) | + | + |
| G. sylvaticum | F | GZst02 | Strawberry (Fragaria× ananassa Duch.) | + | + |
| G. sylvaticum | F | GZso05 | Soybean (Glycine max (L.) Merr.) | + | + |
| G. sylvaticum | F | GZle07 | Lettuces (Lactuca sativa var. ramosa Hort.) | + | + |
| G. sylvaticum | F | GZbr07 | Brassica napus L. | + | + |
| G. sylvaticum | F | GZpu58 | Pumpkins (Cucurbita moschata Duchesne ex Poir.) | + | + |
| G. sylvaticum | F | GZcu17 | Cucumbers (Cucumis sativus L.) | + | + |
| G. sylvaticum | F | GZgi09 | Ginger (Zingiber officinale Roscoe) | + | + |
| G. sylvaticum | F | GZeg44 | Eggplants (Solanum melongena L.) | + | + |
| G. parvum | G | GZal2 | Allium schoenoprasum L. | − | − |
| G. heterothallicum | I | Gzla21 | Lactuca | − | − |
| G. ultimum | I | GZph1 | Phaseolus vulgaris Linn. | − | − |
| G. nodosum | J | GZHs15 | Soil | − | − |
| Phytopythium helicoides | K | GZw1 | Water | − | − |
| Phy. vexans | K | GZHs24 | Soil | − | − |
| Phytophthora nicotianae | 1 | GZst31 | Strawberry (Fragaria× ananassa Duch.) | − | − |
| Ph. cactorum | 1 | GZst21 | Strawberry (Fragaria× ananassa Duch.) | − | − |
| Ph. capsic | 2 | GZsm21 | Solanum melongena L. | − | − |
| Colletotrichum siamense | 1-0 | Camellia sinensis (L.) O. Ktze. | − | − | |
| Alternaria tenuissima | V832 | Solanum tuberosum L. | − | − | |
| Fusarium oxysporum | 1-7 | Capsicum annuum L. | − | − | |
| Species | Primer Set | Primers | Sequences (5′-3′) | Region Amplified |
|---|---|---|---|---|
| G. sylvaticum | L-Psy (LAMP) | F3 | TGCTTATTGGGTGTCTGTTC | rDNA-ITS |
| FIP | AGCCGCCCACTACTAACAA~TCGCCTTGAGGTGTACTGG | |||
| B3 | TCTTGTCTGATATCAGGTCCA | |||
| BIP | ACTTGTGCAATTGGCAGAA~CAGGATCAAACCCGGAGTAC | |||
| F-loop | AACCAGTTCAATCCCACAGC | |||
| P-Psy(PCR) | For | TTCAAACCCCATACCTAACTT | rDNA-ITS | |
| Rev | CGCAAGTTGTGCATAAACAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Sun, X.; Ao, N.; Zou, H.; Shao, H.; Kageyama, K.; Feng, W. Host Range and Loop-Mediated Isothermal Amplification Detection of Globisporangium sylvaticum from Guizhou, China. J. Fungi 2023, 9, 752. https://doi.org/10.3390/jof9070752
Zhang J, Sun X, Ao N, Zou H, Shao H, Kageyama K, Feng W. Host Range and Loop-Mediated Isothermal Amplification Detection of Globisporangium sylvaticum from Guizhou, China. Journal of Fungi. 2023; 9(7):752. https://doi.org/10.3390/jof9070752
Chicago/Turabian StyleZhang, Jing, Xiaonan Sun, Ningjing Ao, Huayan Zou, Huijuan Shao, Koji Kageyama, and Wenzhuo Feng. 2023. "Host Range and Loop-Mediated Isothermal Amplification Detection of Globisporangium sylvaticum from Guizhou, China" Journal of Fungi 9, no. 7: 752. https://doi.org/10.3390/jof9070752