

New Insights in Phenothiazinium-Mediated Photodynamic Inactivation of Candida Auris
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. auris Growth Conditions
2.2. Photosensitizers and Light Source
2.3. Cell Suspension Preparation and PDI
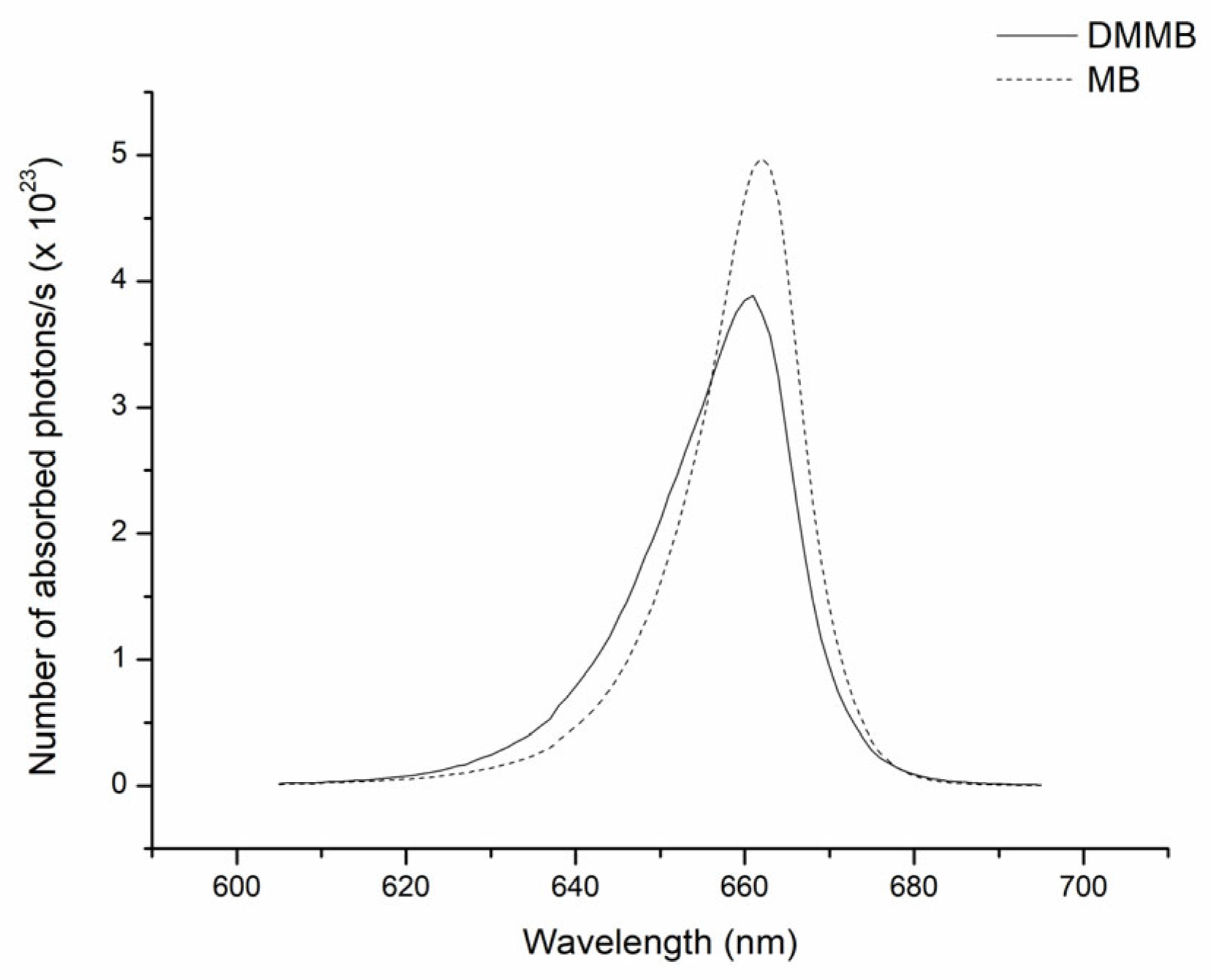
2.4. Number of Absorbed Photons
- c: velocity of light in vacuum (~3 × 108 m/s);
- h: Planck constant (6.62606957 × 10−34 J·s).
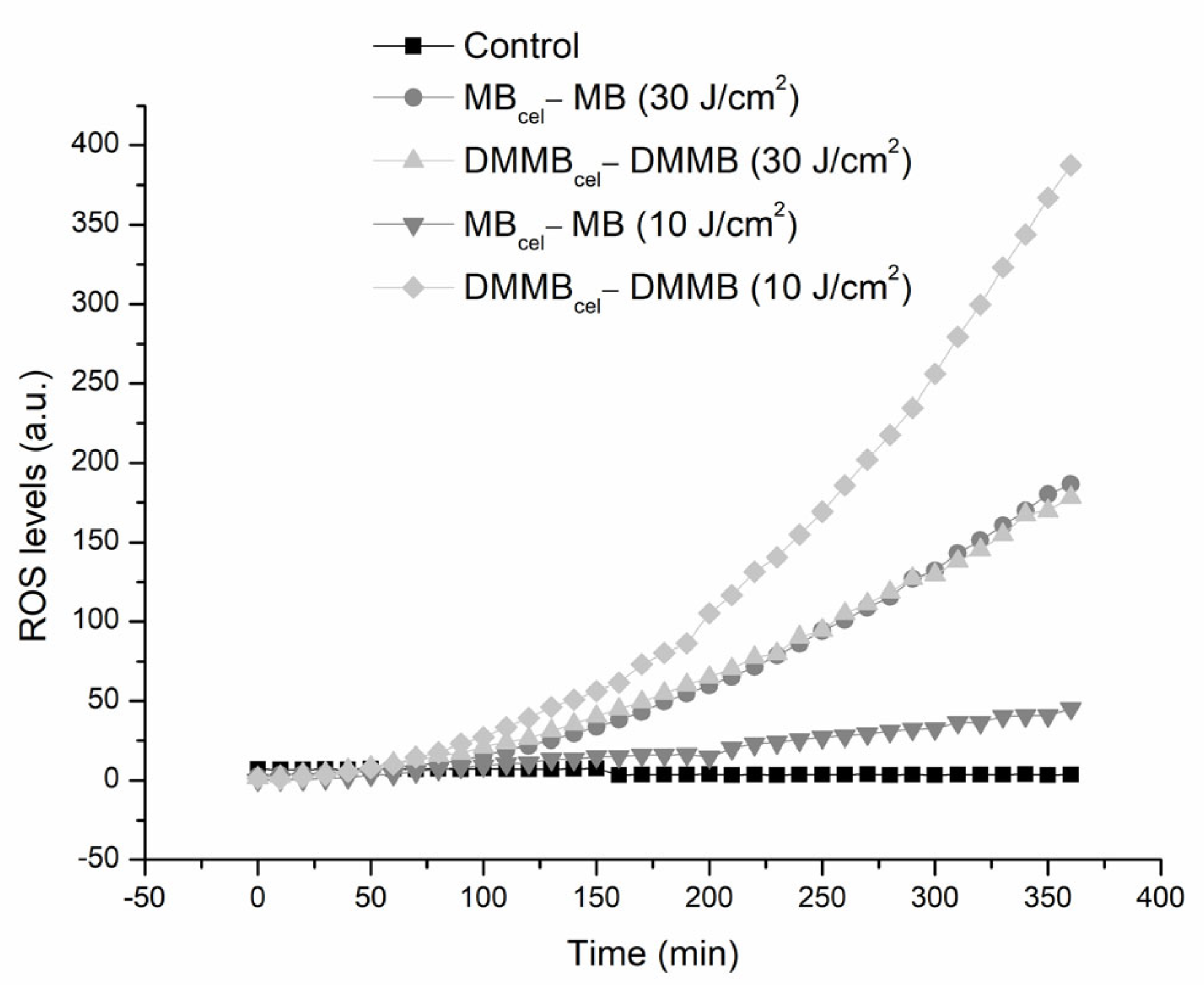
2.5. ROS Kinetics
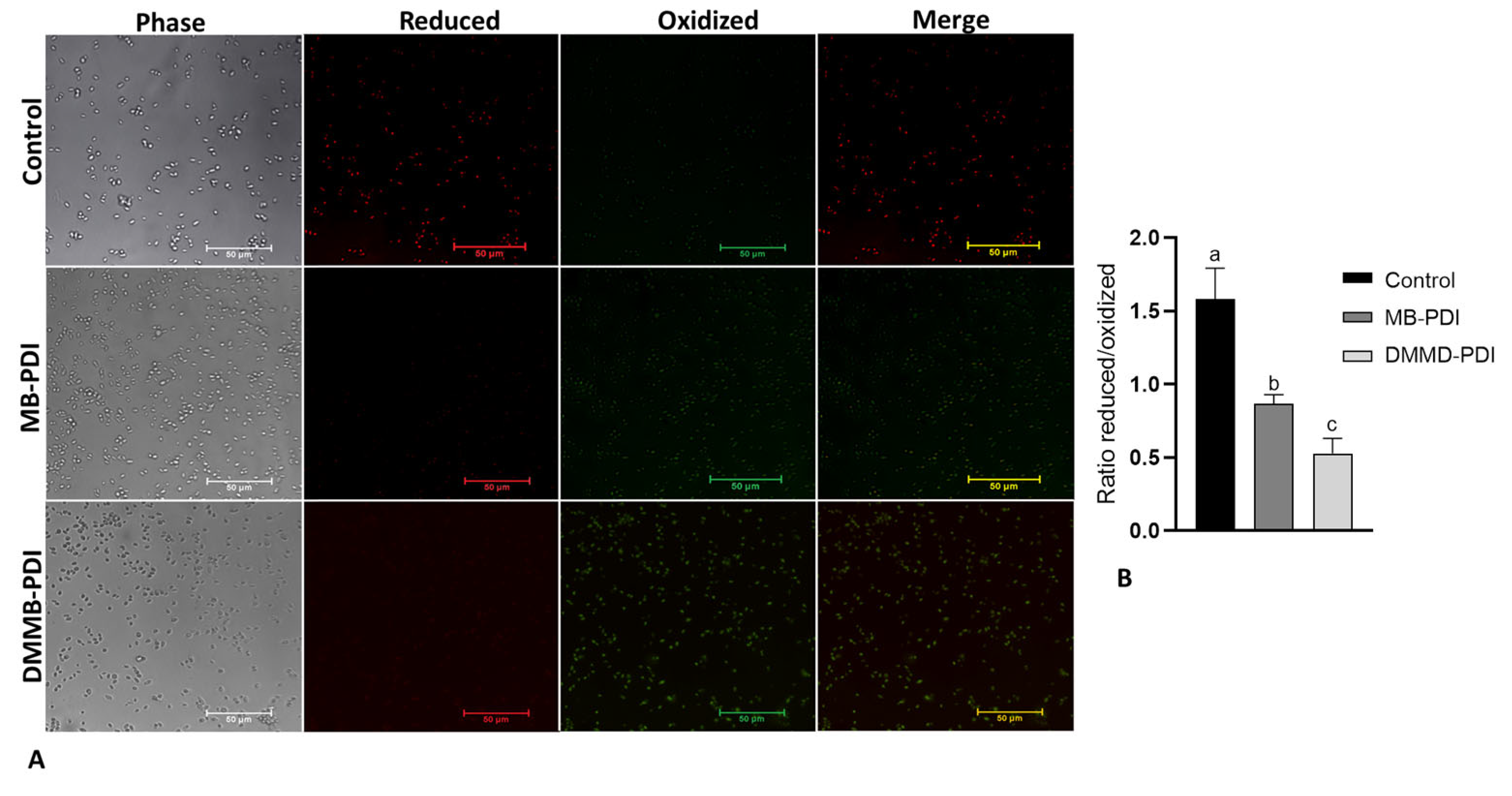
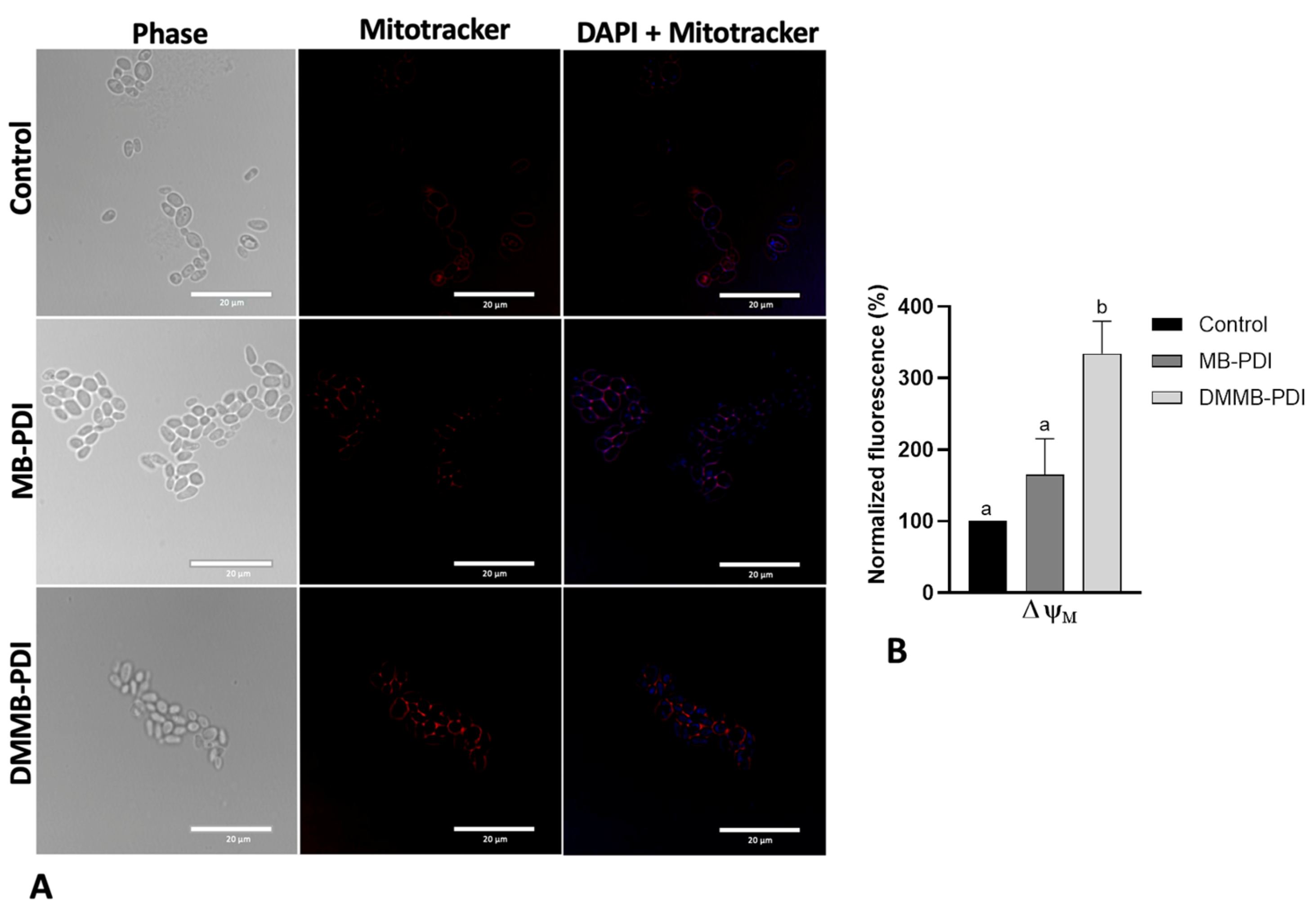
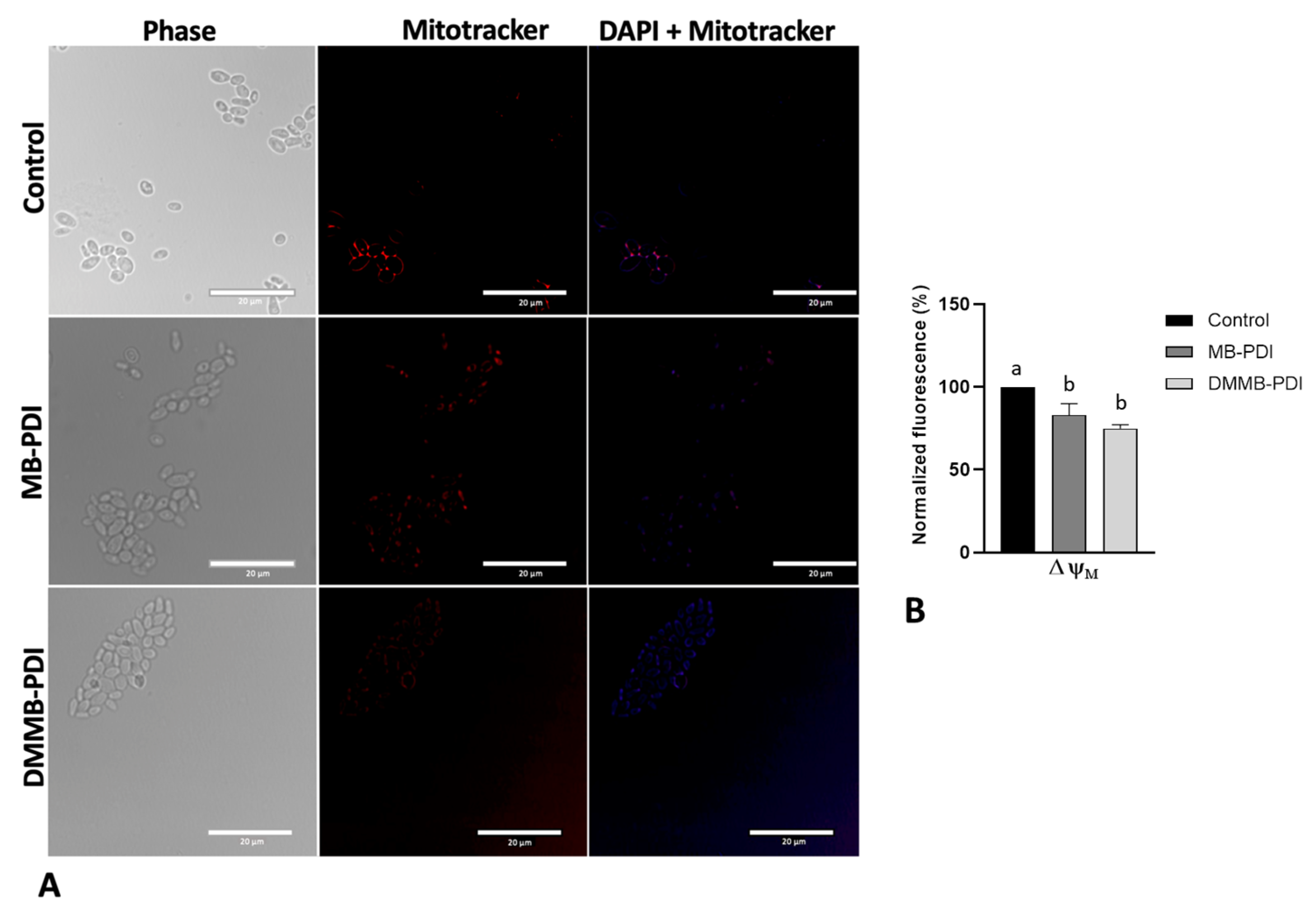
2.6. Peroxidation Lipid and Mitochondrial Membrane Potential (ΔΨm)
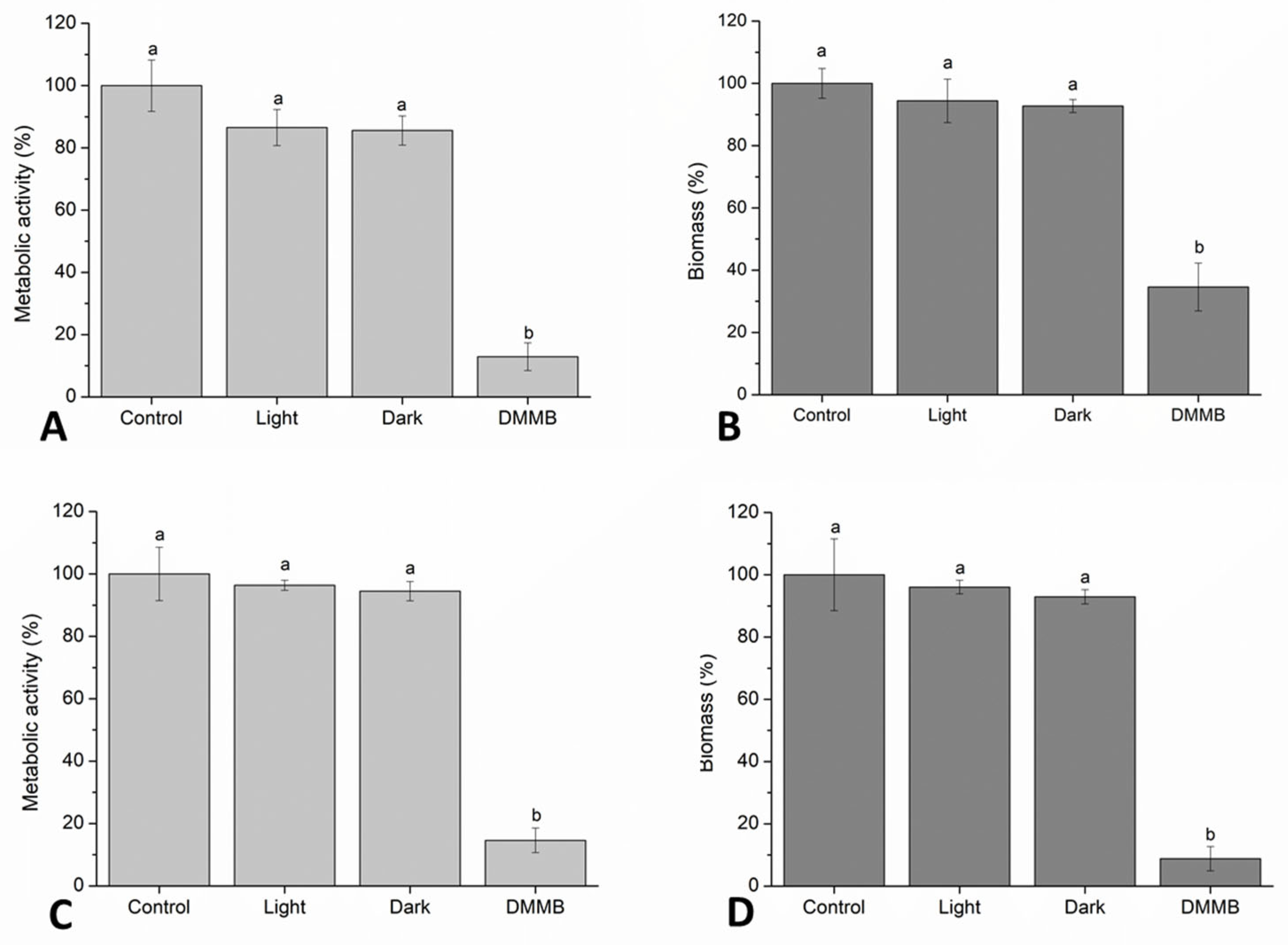
2.7. Biofilm Formation and PDI
2.8. Analysis of Metabolic Activity and Biomass in Biofilms
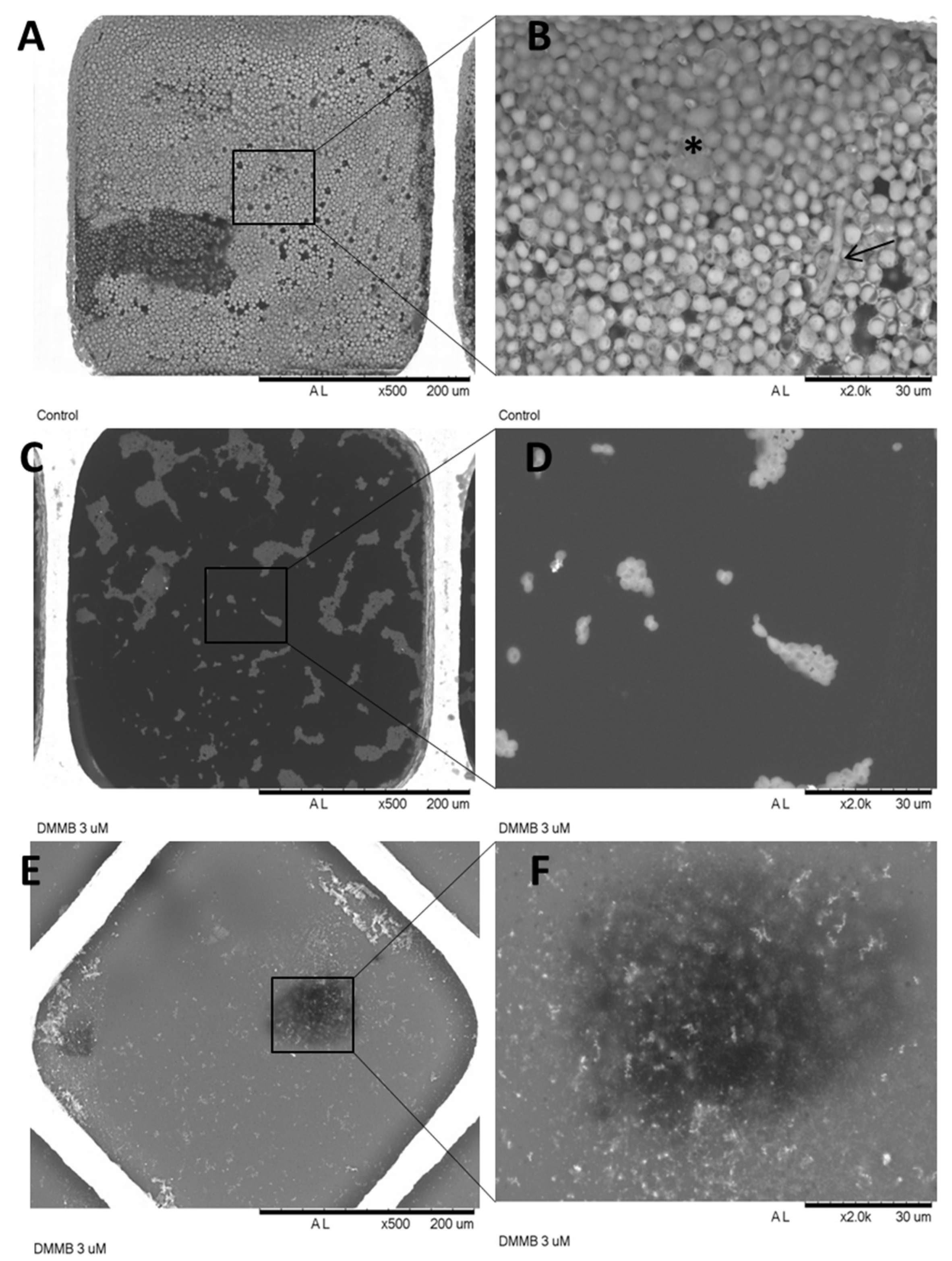
2.9. Structural Changes by SEM and OCT
2.10. Cytotoxicity on Mammalian Cells
2.11. Statistical Analysis
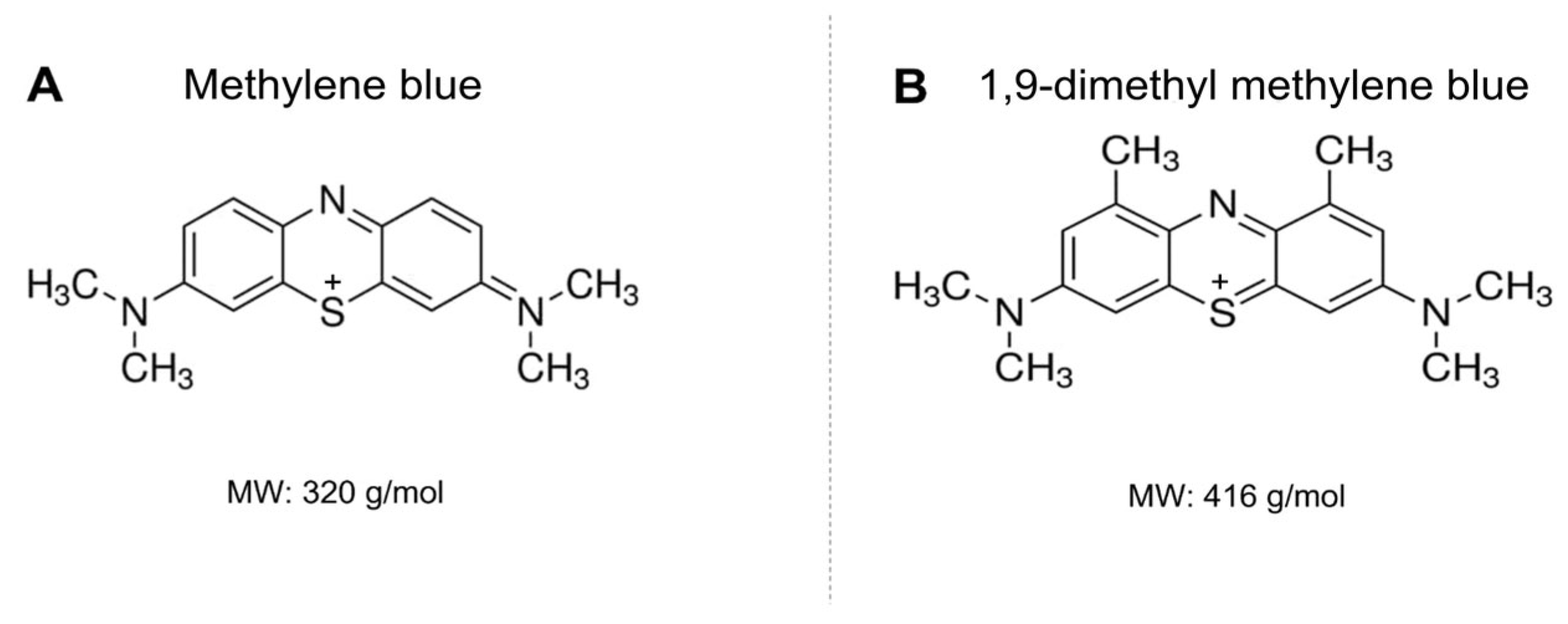
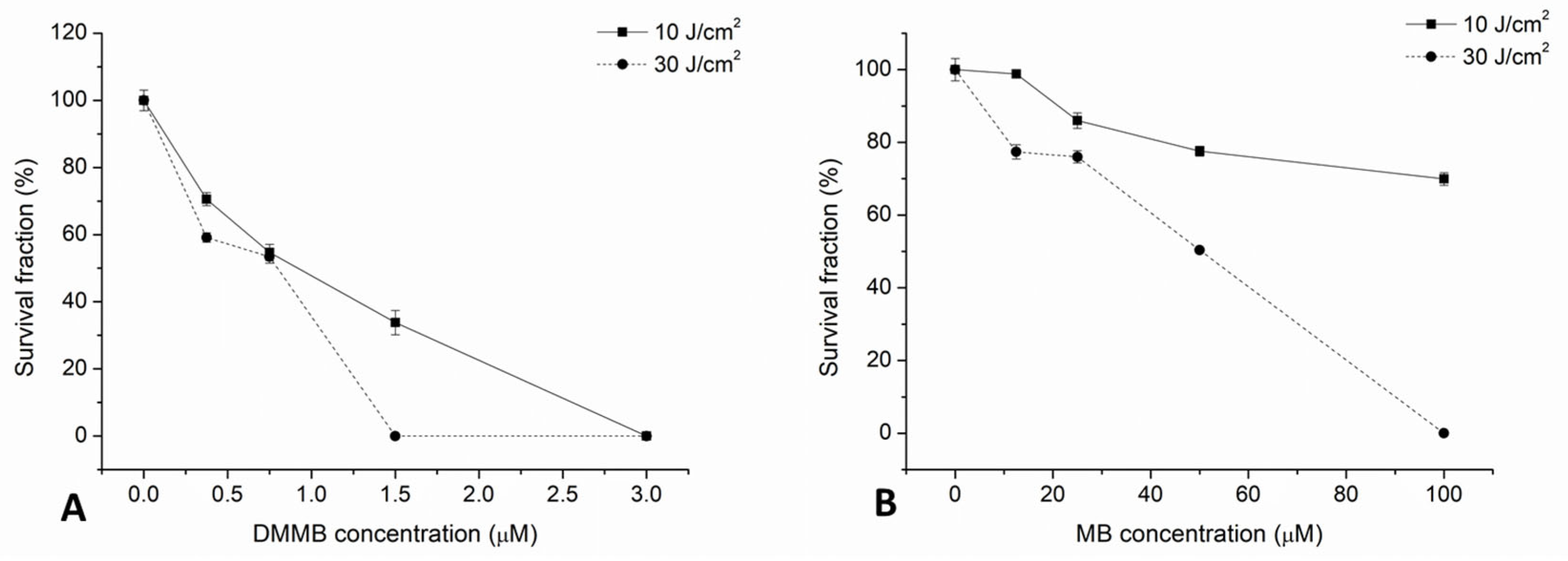
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Watkins, R.R.; Gowen, R.; Lionakis, M.S.; Ghannoum, M. Update on the Pathogenesis, Virulence, and Treatment of Candida auris. Pathog. Immun. 2022, 7, 46–65. [Google Scholar] [CrossRef]
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; WHO Technical Report Series; World Health Organization: Geneva, Switzerland, 2022; 48p. [Google Scholar]
- Fisher, M.C.; Denning, D.W. The WHO fungal priority pathogens list as a game-changer. Nat. Rev. Microbiol. 2023, 21, 211–212. [Google Scholar] [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10, e01397-19. [Google Scholar] [CrossRef] [Green Version]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial photodynamic therapy-what we know and what we don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [Green Version]
- Ziental, D.; Mlynarczyk, D.T.; Czarczynska-Goslinska, B.; Lewandowski, K.; Sobotta, L. Photosensitizers Mediated Photodynamic Inactivation against Fungi. Nanomater 2021, 11, 2883. [Google Scholar] [CrossRef]
- Freire, F.; de Barros, P.P.; Pereira, C.A.; Junqueira, J.C.; Jorge, A.O.C. Photodynamic inactivation in the expression of the Candida albicans genes ALS3, HWP1, BCR1, TEC1, CPH1, and EFG1 in biofilms. Lasers Med. Sci. 2018, 33, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Štefánek, M.; Černáková, L.; Dekkerová, J.; Bujdáková, H. Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes. J. Fungi 2022, 8, 1137. [Google Scholar] [CrossRef]
- Ziental, D.; Mlynarczyk, D.T.; Kolasinski, E.; Güzel, E.; Dlugaszewska, J.; Popenda, Ł.; Jurga, S.; Goslinski, T.; Sobotta, L. Zinc(II), Palladium(II), and Metal-Free Phthalocyanines Bearing Nipagin-Functionalized Substituents against Candida auris and Selected Multidrug-Resistant Microbes. Pharmaceutics 2022, 14, 1686. [Google Scholar] [CrossRef]
- Liu, X.; Fang, R.; Feng, R.; Li, Q.; Su, M.; Hou, C.; Zhuang, K.; Dai, Y.; Lei, N.; Jiang, Y.; et al. Cage-modified hypocrellin against multidrug-resistant Candida spp. with unprecedented activity in light-triggered combinational photodynamic therapy. Drug Resist. Updates 2022, 65, 100887. [Google Scholar] [CrossRef]
- Mertins, O.; Bacellar, I.O.; Thalmann, F.; Marques, C.M.; Baptista, M.S.; Itri, R. Physical damage on giant vesicles membrane as a result of methylene blue photoirradiation. Biophys. J. 2014, 106, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Campos Chaves Lamarque, G.; Cusicanqui Méndez, D.A.; Arruda Matos, A.; José Dionísio, T.; Andrade Moreira Machado, M.A.; Magalhães, A.C.; Cardoso Oliveira, R.; Cruvinel, T. Cytotoxic effect and apoptosis pathways activated by methylene blue-mediated photodynamic therapy in fibroblasts. Photodiagnos. Photodyn. Ther. 2020, 29, 101654. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, L.; Gu, X.; Yu, D.; Li, S. Amphiphilic di-cationic methylene blue for improving antibacterial photodynamic efficiency through high accumulation and low aggregation on bacterial cell surfaces. RSC Adv. 2022, 13, 239–250. [Google Scholar] [CrossRef]
- Gabrielli, D.; Belisle, E.; Severino, D.; Kowaltowski, A.J.; Baptista, M.S. Binding, aggregation and photochemical properties of methylene blue in mitochondrial suspensions. Photochem. Photobiol. 2004, 79, 227–232. [Google Scholar] [CrossRef]
- Wainwright, M.; Phoenix, D.A.; Rice, L.; Burrow, S.M.; Waring, J. Increased cytotoxicity and phototoxicity in the methylene blue series via chromophore methylation. J. Photochem. Photobiol. B 1997, 40, 233–239. [Google Scholar] [CrossRef]
- Paz-Cristobal, M.P.; Royo, D.; Rezusta, A.; Andrés-Ciriano, E.; Alejandre, M.C.; Meis, J.F.; Revillo, M.J.; Aspiroz, C.; Nonell, S.; Gilaberte, Y. Photodynamic fungicidal efficacy of hypericin and dimethyl methylene blue against azole-resistant Candida albicans strains. Mycoses 2014, 57, 35–42. [Google Scholar] [CrossRef]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Spettel, K.; Kriz, R.; Wu, C.; Achter, L.; Schmid, S.; Galazka, S.; Selitsch, B.; Camp, I.; Makristathis, A.; Lagler, H.; et al. Candida auris in Austria—What Is New and What Is Different. J. Fungi 2023, 9, 129. [Google Scholar] [CrossRef]
- Gonzales, F.P.; Da Silva, S.H.; Roberts, D.W.; Braga, G.U.L. Photodynamic inactivation of conidia of the fungi Metarhizium anisopliae and Aspergillus nidulans with methylene blue and toluidine blue. Photochem. Photobiol. 2010, 86, 653–661. [Google Scholar] [CrossRef]
- Prates, R.A.; Fuchs, B.B.; Mizuno, K.; Naqvi, Q.; Kato, I.T.; Ribeiro, M.S.; Mylonakis, E.; Tegos, G.P.; Hamblin, M.R. Effect of Virulence Factors on the Photodynamic Inactivation of Cryptococcus neoformans. PLoS ONE 2013, 8, e54387. [Google Scholar] [CrossRef] [Green Version]
- de Senna, A.M.; Vieira, M.M.F.; Machado-de-Sena, R.M.; Bertolin, A.O.; Núñez, S.C.; Ribeiro, M.S. Photodynamic inactivation of Candida ssp. on denture stomatitis. A clinical trial involving palatal mucosa and prosthesis disinfection. Photodiagnos. Photodyn. Ther. 2018, 22, 212–216. [Google Scholar] [CrossRef]
- Alberdi, E.; Gomez, C. Efficiency of methylene blue-mediated photodynamic therapy vs intense pulsed light in the treatment of onychomycosis in the toenails. Photodermatol. Photoimmunol. Photomed. 2019, 35, 69–77. [Google Scholar] [CrossRef] [PubMed]
- de Lapena, S.A.B.; Terra-Garcia, M.; Ward, R.A.d.C.; Rossoni, R.D.; Melo, V.M.M.; Junqueira, J.C. Enhancing effect of chitosan on methylene blue-mediated photodynamic therapy against C. albicans: A study in planktonic growth, biofilms, and persister cells. Photodiagnos. Photodyn. Ther. 2022, 38, 102837. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Laguna, V.; Barrena-López, Y.; Gilaberte, Y.; Rezusta, A. In Vitro Effect of Photodynamic Therapy with Different Lights and Combined or Uncombined with Chlorhexidine on Candida spp. Pharmaceutics 2021, 13, 1176. [Google Scholar] [CrossRef]
- Nunes, I.P.F.; Crugeira, P.J.L.; Sampaio, F.J.P.; de Oliveira, S.C.P.S.; Azevedo, J.M.; Santos, C.L.O.; Soares, L.G.P.; Samuel, I.D.W.; Persheyev, S.; de Ameida, P.F.; et al. Evaluation of dual application of photodynamic therapy—PDT in Candida albicans. Photodiagnos. Photodyn. Ther. 2023, 42, 103327. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Pummer, A.; Regensburger, J.; Hiller, K.A.; Späth, A.; Tabenski, L.; Buchalla, W.; Maisch, T. The impact of absorbed photons on antimicrobial photodynamic efficacy. Front. Microbiol. 2015, 6, 706. [Google Scholar] [CrossRef] [Green Version]
- Bacellar, I.O.L.; Pavani, C.; Sales, E.M.; Itri, R.; Wainwright, M.; Baptista, M.S. Membrane damage efficiency of phenothiazinium photosensitizers. Photochem. Photobiol. 2014, 90, 801–813. [Google Scholar] [CrossRef]
- Belozerskaya, T.A.; Gessler, N.N. Reactive oxygen species and the strategy of antioxidant defense in fungi: A review. Appl. Biochem. Microbiol. 2007, 43, 506–515. [Google Scholar] [CrossRef]
- Dantas, A.D.S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative Stress Responses in the Human Fungal Pathogen, Candida albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [Green Version]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids and Their Biosynthesis in Fungi. Molecules 2022, 27, 1431. [Google Scholar] [CrossRef]
- Shimada, H.; Kondo, K.; Fraser, P.D.; Miura, Y.; Saito, T.; Misawa, N. Increased Carotenoid Production by the Food Yeast Candida utilis through Metabolic Engineering of the Isoprenoid Pathway. Appl. Environ. Microbiol. 1998, 64, 2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacellar, I.O.L.; Baptista, M.S. Mechanisms of Photosensitized Lipid Oxidation and Membrane Permeabilization. ACS Omega 2019, 4, 21636–21646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, W.K.; Santos, N.F.; Rocha, C.D.S.; Bacellar, I.O.L.; Tsubone, T.M.; Viotto, A.C.; Matsukuma, A.Y.; Abrantes, A.B.d.P.; Siani, P.; Dias, L.G.; et al. Parallel damage in mitochondria and lysosomes is an efficient way to photoinduce cell death. Autophagy 2019, 15, 259–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.Y.; Sun, L.; Chen, X.P.; Zhang, D.S. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Ferreira, S.D.R.M.; Tedesco, A.C.; Sousa, G.; Zângaro, R.A.; Silva, N.S.; Pacheco, M.T.T.; Pacheco-Soares, C. Analysis of mitochondria, endoplasmic reticulum and actin filaments after PDT with AlPcS4. Lasers Med. Sci. 2004, 18, 207–212. [Google Scholar] [CrossRef]
- Andrade, C.G.; Figueiredo, R.C.B.Q.; Ribeiro, K.R.C.; Souza, L.I.O.; Sarmento-Neto, J.F.; Rebouças, J.S.; Santos, B.S.; Ribeiro, M.S.; Carvalho, L.B.; Fontes, A. Photodynamic effect of zinc porphyrin on the promastigote and amastigote forms of: Leishmania braziliensis. Photochem. Photobiol. Sci. 2018, 17, 482–490. [Google Scholar] [CrossRef]
- Vianello, A.; Casolo, V.; Petrussa, E.; Peresson, C.; Patui, S.; Bertolini, A.; Passamonti, S.; Braidot, E.; Zancani, M. The mitochondrial permeability transition pore (PTP)-an example of multiple molecular exaptation? Biochim. Biophys. Acta 2012, 1817, 2072–2086. [Google Scholar] [CrossRef] [Green Version]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef]
- Horton, M.V.; Nett, J.E. Candida auris infection and biofilm formation: Going beyond the surface. Curr. Clin. Microbiol. Rep. 2020, 7, 51–56. [Google Scholar] [CrossRef]
- Pezzotti, G.; Kobara, M.; Nakaya, T.; Imamura, H.; Fujii, T.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; et al. Raman Metabolomics of Candida auris Clades: Profiling and Barcode Identification. Int. J. Mol. Sci. 2022, 23, 11736. [Google Scholar] [CrossRef] [PubMed]
- Oremefetse, D.; Aijaz, A.; Sanelisiwe, D.; Mrudula, P. Survival of Candida auris on environmental surface material and low-level resistance to disinfectant. J. Hosp. Infect. 2023, 137, 17–23. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candida Strain | Form | PS (Concentration) (µM) | PIT (min) | Light Source | λmax (nm) | Irradiance (mW/cm2) | Light Dose (J/cm2) | Exposure Time (s) | Outcome |
|---|---|---|---|---|---|---|---|---|---|
| C. albicans (ATCC 18804 and Ca70 fluconazole-resistant) | Planktonic | MB (300) | 15 | LED | 660 | 42 | 30 | 714 | ~99% log reduction regardless of the resistance pattern |
| Biofilm 48 h | MB (600) | ~99.9% reduction regardless of the resistance pattern [23] | |||||||
| C. albicans ATCC 10231, C. parapsilosis ATCC 22019 and C. krusei ATCC 6258 | Planktonic | MB (1000) * | 0 | LED | 625 | 7 | 18 | 2571.6 | 99.9999% reduction regardless of the strain. PS concentration for C. parapsilosis was 250 μM [24] |
| White lamp | 420–700 | 90 | 205 | ||||||
| C. auris (H261 and fluconazole-resistant) | Biofilm 24 h | MB (250) | 15 | Laser | 660 | 190 | 58 | 300 | >90% reduction [8] |
| C. albicans (ATCC 10231 and AZN9635, 456325H, AMO7/0267 azole-resistants) | Planktonic | DMMB (0.6 to 2.5) | NI | LED | 639.8 | 19 | 18 and 37 | NI | >99.9% reduction regardless of the resistance pattern [16] |
| C. albicans (ATCC 90028) | Planktonic | DMMB (2.4) * | 5 | LED | 630 | 41.2 | 20 | 870 | 99.9% reduction [25] |
| C. auris (CBS 10913) | Planktonic | MB (100 μM) DMMB (1.5 μM) | 5 | LED | 662 | 50 | 30 | 596 | 100% reduction |
| Biofilm 24 h | DMMB (3 μM) | 10 | 198 | >85% reduction (this work) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.R.; Cabral, F.V.; Silva, C.R.; Silva, D.F.T.; Freitas, A.Z.; Fontes, A.; Ribeiro, M.S. New Insights in Phenothiazinium-Mediated Photodynamic Inactivation of Candida Auris. J. Fungi 2023, 9, 717. https://doi.org/10.3390/jof9070717
Silva AR, Cabral FV, Silva CR, Silva DFT, Freitas AZ, Fontes A, Ribeiro MS. New Insights in Phenothiazinium-Mediated Photodynamic Inactivation of Candida Auris. Journal of Fungi. 2023; 9(7):717. https://doi.org/10.3390/jof9070717
Chicago/Turabian StyleSilva, Abdênego R., Fernanda V. Cabral, Camila R. Silva, Daniela F. T. Silva, Anderson Z. Freitas, Adriana Fontes, and Martha S. Ribeiro. 2023. "New Insights in Phenothiazinium-Mediated Photodynamic Inactivation of Candida Auris" Journal of Fungi 9, no. 7: 717. https://doi.org/10.3390/jof9070717