
Genetic and Environmental Factors Influencing the Production of Select Fungal Colorants: Challenges and Opportunities in Industrial Applications


Abstract
:1. Introduction
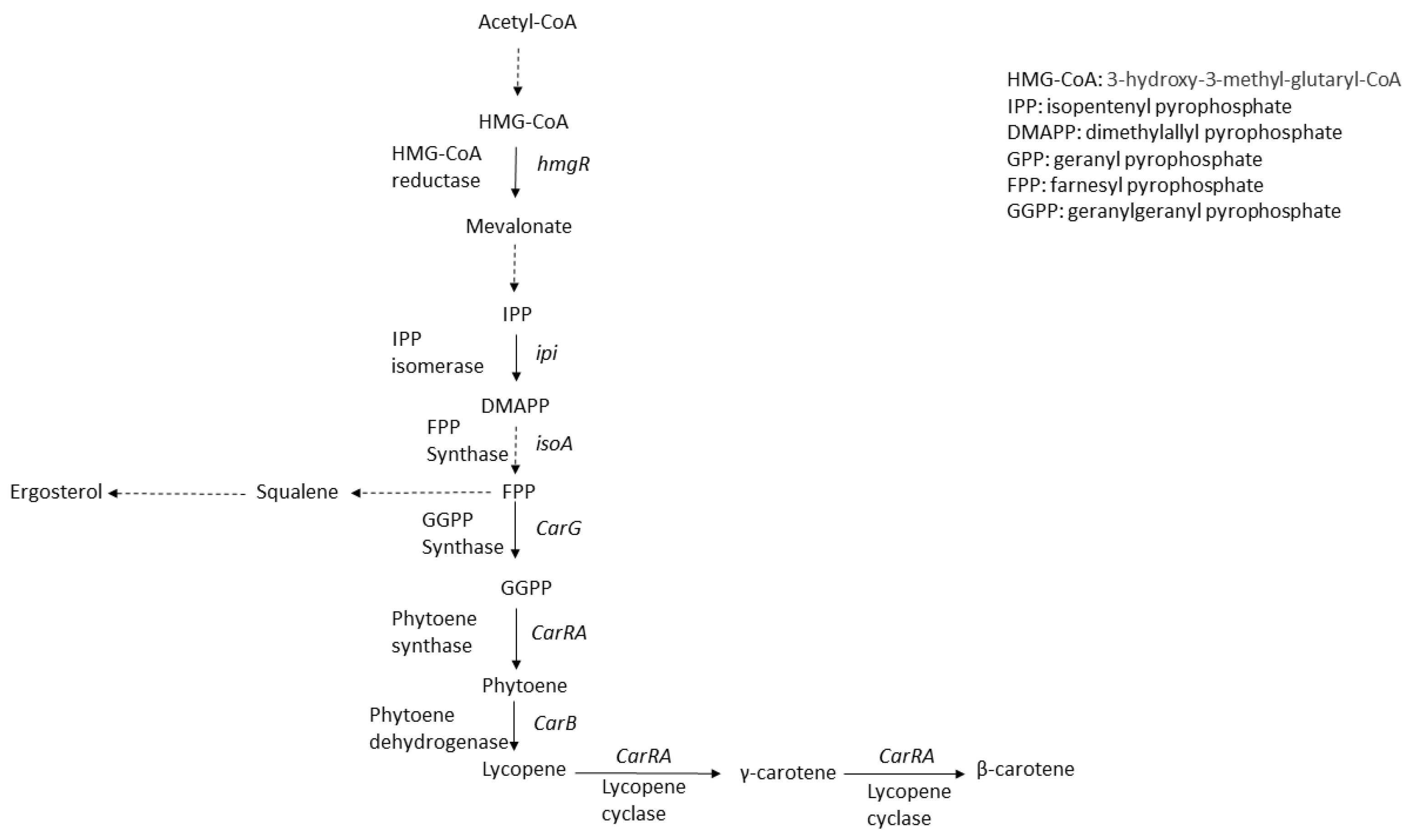
2. Carotenoids
2.1. Carotenoids and Carotenoid Synthesis Pathways
2.2. Production of Carotenoids
3. Melanins
3.1. Melanins and Melanin Synthesis Pathways
3.2. Production of Fungal Melanins
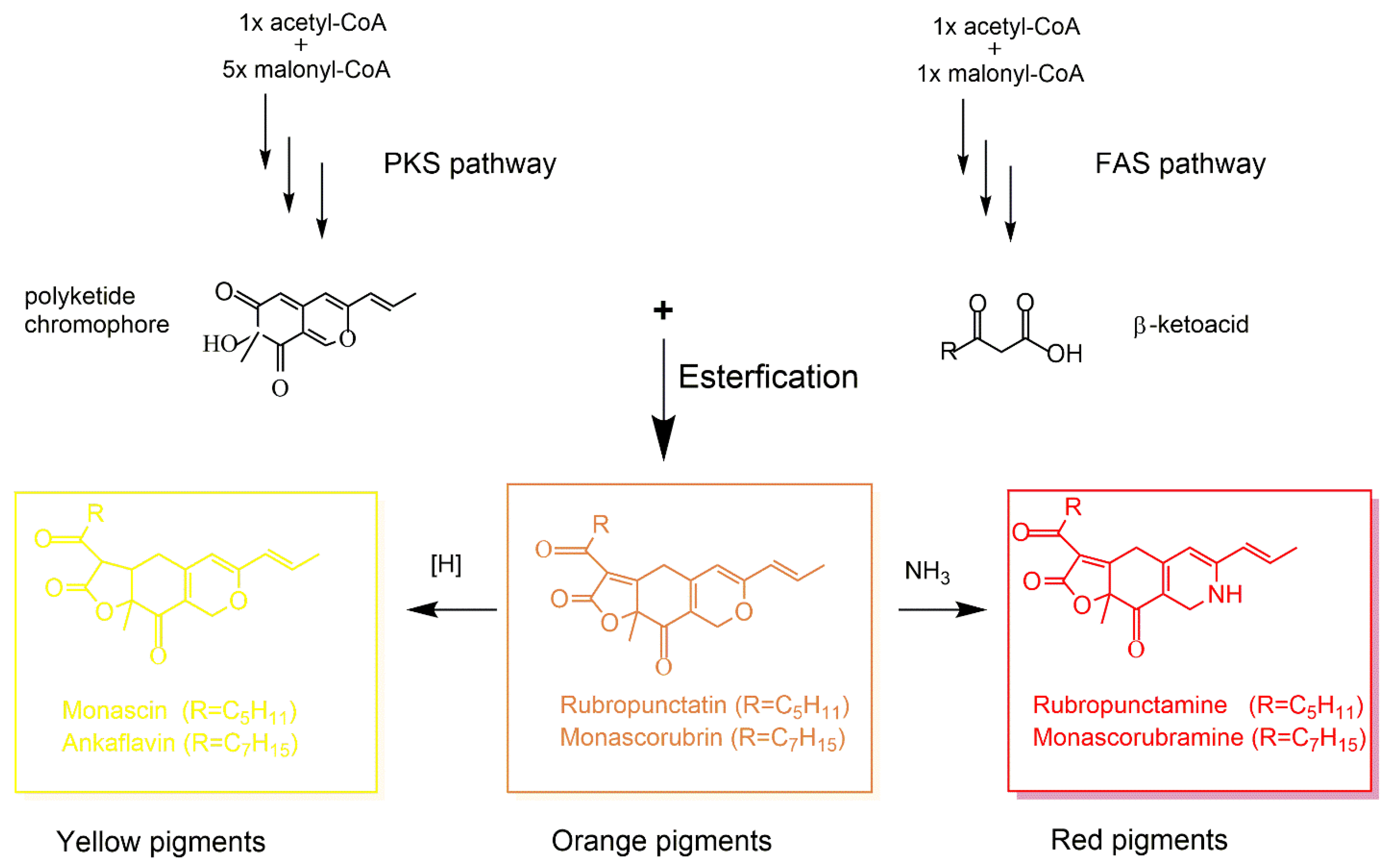
4. Fungal Polyketide-Derived Colorants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demain, A.L.; Vandamme, E.J.; Collins, J.; BuchhoIz, K. History of industrial biotechnology. In Industrial Biotechnology: Microorganisms; Wittmann, C., Liao, J.C., Eds.; Wiley-VCH: Weinheim, Germany, 2017; pp. 3–84. [Google Scholar]
- Chatragadda, R.; Dufossé, L. Ecological and biotechnological aspects of pigmented microbes: A way forward in development of food and pharmaceutical grade pigments. Microorganisms 2021, 9, 637. [Google Scholar] [CrossRef]
- Sen, T.; Barrow, C.J.; Deshmukh, S.K. Microbial pigments in the food industry—Challenges and the way forward. Front. Nutr. 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Oplatowska-Stachowiak, M.; Christopher, E.T. Food colours: Existing and emerging food safety concerns. Crit. Rev. Food Sci. Nutr. 2015, 57, 524–548. [Google Scholar] [CrossRef]
- Dikshit, R.; Tallapragada, P. Comparative study of natural and artificial flavoring agents and dyes. In Natural and Artificial Flavoring Agents and Food Dyes; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier: London, UK, 2018; Volume 7, pp. 83–111. [Google Scholar]
- McCann, D.; Barrett, A.; Cooper, A.; Crumpler, D.; Dalen, L.; Grimshaw, K.; Kitchin, E.; Lok, K.; Porteous, L.; Prince, E.; et al. Food additives and hyperactive behaviour in 3-year-old and 8/9-year-old children in the community: A randomised, double-blinded, placebo controlled trial. Lancet 2007, 370, 1560–1567. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, J.P. Fungal pigments and their roles associated with human health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef]
- Venil, C.K.; Velmurugan, P.; Dufossé, L.; Devi, P.R.; Ravi, A.V. Fungal pigments: Potential coloring compounds for wide ranging applications in textile dyeing. J. Fungi 2020, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Gürses, A.; Açıkyıldız, M.; Güneş, K.; Gürses, M.S. Dyes and pigments. In SpringerBriefs in Green Chemistry for Sustainability; Sharma, S.K., Ed.; Springer International Publishing AG: Cham, Switzerland, 2016; pp. 13–29. [Google Scholar]
- Urista, C.M.; Rodríguez, J.G.; Corona, A.A.; Cuenca, A.A.; Jurado, A.T. Pigments from fungi, an opportunity of production for diverse applications. Biologia 2016, 71, 1067–1079. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids and their biosynthesis in fungi. Molecules 2022, 27, 1431. [Google Scholar] [CrossRef]
- Nanou, K.; Roukas, T. Waste cooking oil: A new substrate for carotene production by Blakeslea trispora in submerged fermentation. Bioresour. Technol. 2016, 203, 198–203. [Google Scholar] [CrossRef]
- Luo, W.; Xue, C.; Zhao, Y.; Zhang, H.; Rao, Z.; Yu, X. Blakeslea trispora photoreceptors: Identification and functional analysis. Appl. Environ. Microbiol. 2020, 86, e02962-19. [Google Scholar] [CrossRef]
- Bo, S.; Ni, X.; Guo, J.; Liu, Z.; Wang, X.; Sheng, Y.; Zhang, G.; Yang, J. Carotenoid biosynthesis: Genome-wide profiling, pathway identification in Rhodotorula glutinis X-20, and high-level production. Front. Nutr. 2022, 9, 918240. [Google Scholar] [CrossRef] [PubMed]
- Nutakor, C.; Kanwugu, O.N.; Kovaleva, E.G.; Glukhareva, T.V. Enhancing astaxanthin yield in Phaffia rhodozyma: Current trends and potential of phytohormones. Appl. Microbiol. Biotechnol. 2022, 106, 3531–3538. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Sáiz, M.; Paz, B.; de la Fuente, J.L.; Lo´pez-Nieto, M.J.; Cabri, W.; Barredo, J.L. Blakeslea trispora genes for carotene biosynthesis. Appl. Eniviron. Microbiol. 2004, 70, 5589–5594. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Schmidt-Dannert, C. Metabolic engineering towards biotechnological production of carotenoids in microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 1–11. [Google Scholar] [PubMed]
- Mussagy, C.U.; Winterburn, J.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Production and extraction of carotenoids produced by microorganisms. Appl. Microbiol. Biotechnol. 2019, 103, 1095–1114. [Google Scholar] [CrossRef]
- Sun, J.; Sun, X.X.; Tang, P.W.; Yuan, Q.P. Molecular cloning and functional expression of two key carotene synthetic genes derived from Blakeslea trispora into E. coli for increased β-carotene production. Biotechnol. Lett. 2012, 34, 2077–2082. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, C.; Bi, C.; Li, Q.; Zhang, X. Combinatory optimization of chromosomal integrated mevalonate pathway for β-carotene production in Escherichia coli. Microb. Cell Fact. 2016, 15, 202. [Google Scholar] [CrossRef]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of β-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef]
- Liu, L.; Qu, Y.L.; Dong, G.R.; Wang, J.; Hu, C.Y.; Meng, Y.H. Elevated β-carotene production using codon-adapted CarRA &B and metabolic balance in engineered Yarrowia lipolytica. Front. Microbiol. 2021, 12, 627150. [Google Scholar]
- Zhang, Y.; Navarro, E.; Cánovas-Márquez, J.T.; Almagro, L.; Chen, H.; Chen, Y.Q.; Zhang, H.; Torres-Martínez, S.; Chen, W.; Garre, V. A new regulatory mechanism controlling carotenogenesis in the fungus Mucor circinelloides as a target to generate β-carotene over-producing strains by genetic engineering. Microb. Cell Fact. 2016, 15, 99. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Reitzel, K.; Nielsen, U.G.; Dupuy-Galet, B.X.; Leivers, S.; Horn, S.J.; Shapaval, V. Calcium affects polyphosphate and lipid accumulation in Mucoromycota fungi. J. Fungi 2021, 7, 300. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Limon, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef]
- Echavarri-Erasun, C.; Johnson, E.A. Stimulation of astaxanthin formation in the yeast Xanthophyllomyces dendrorhous by the fungus Epicoccum nigrum. FEMS Yeast Res. 2004, 4, 511–519. [Google Scholar] [CrossRef]
- Ge, X.; Li, R.; Zhang, X.; Zhao, J.; Zhang, Y.; Xin, Q. Transcriptome sequencing and global analysis of blue light-responsive genes provide clues for high carotenoid yields in Blakeslea trispora. Int. Microbiol. 2022, 25, 325–338. [Google Scholar] [CrossRef]
- Bindl, E.; Lang, W.; Rau, W. Light dependent carotenoid synthesis. 6. Time course of synthesis of various carotenoids in Fusarium aquaeductuum after various inductive treatments. Planta 1970, 94, 156–174. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Schrott, E.L. Photoinduction of carotenoid biosynthesis in Gibberella fujikuroi. FEMS Microbiol. Lett. 1990, 66, 295–298. [Google Scholar] [CrossRef]
- Rodríguez-Ortiz, R.; Michielse, C.; Rep, M.; Limón, M.C.; Avalos, J. Genetic basis of carotenoid overproduction in Fusarium oxysporum. Fungal Genet. Biol. 2012, 49, 684–696. [Google Scholar] [CrossRef]
- Rodríguez-Ortiz, R.; Limón, M.C.; Avalos, J. Functional analysis of the carS gene of Fusarium fujikuroi. Mol. Genet. Genomics 2013, 288, 157–173. [Google Scholar] [CrossRef]
- Castrillo, M.; Avalos, J. The flavoproteins CryD and VvdA cooperate with the white collar protein WcoA in the control of photocarotenogenesis in Fusarium fujikuroi. PLoS ONE 2015, 10, e0119785. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Zhan, W.; Li, Y.; Wang, X. Temperature influences β-carotene production in recombinant Saccharomyces cerevisiae expressing carotenogenic genes from Phaffia rhodozyma. World J. Microbiol. Biotechnol. 2014, 30, 125–133. [Google Scholar] [CrossRef]
- Kalra, R.; Conlan, X.A.; Goel, M. Fungi as a potential source of pigments: Harnessing filamentous fungi. Front. Chem. 2020, 8, 369. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Park, H.M.; Kim, J.E.; Lee, S.H.; Choi, M.S.; Kim, J.Y.; Oh, D.K.; Keasling, J.D.; Kim, S.W. Increased β-carotene production in recombinant Escherichia coli harboring an engineered isoprenoid precursor pathway with mevalonate addition. Biotechnol. Prog. 2007, 23, 599–605. [Google Scholar] [CrossRef]
- Alper, H.; Miyaoku, K.; Stephanopoulos, G. Construction of lycopene-overproducing E. coli strains by combining systematic and combinatorial gene knockout targets. Nat. Biotechnol. 2005, 23, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Yamane, Y.; Higashida, K.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Astaxanthin production by Phaffia rhodozyma enhanced in fed-batch culture with glucose and ethanol feeding. Biotechnol. Lett. 1997, 19, 1109–1111. [Google Scholar] [CrossRef]
- Choudhari, S.M.; Ananthanarayan, L.; Singhal, R.S. Use of metabolic stimulators and inhibitors for enhanced production of β-carotene and lycopene by Blakeslea trispora NRRL 2895 and 2896. Biores. Technol. 2008, 99, 3166–3173. [Google Scholar] [CrossRef] [PubMed]
- Naz, T.; Nazir, Y.; Nosheen, S.; Ullah, S.; Halim, H.; Fazili, A.B.A.; Li, S.; Mustafa, K.; Mohamed, H.; Yang, W.; et al. Redirecting metabolic flux towards the mevalonate pathway for enhanced β-carotene production in M. circinelloides CBS 277.49. BioMed Res. Intl. 2020, 2020, 8890269. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Y.; Yang, Q.; Zhao, J.; Feng, L.; Wang, M. SR5AL serves as a key regulatory gene in lycopene biosynthesis by Blakeslea trispora. Microb. Cell Fact. 2022, 21, 126. [Google Scholar] [CrossRef]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial melanin: Recent advances in biosynthesis, extraction, characterization, and applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef]
- Pralea, I.-E.; Moldovan, R.-C.; Petrache, A.-M.; Ilieș, M.; Hegheș, S.-C.; Ielciu, I.; Nicoară, R.; Moldovan, M.; Ene, M.; Radu, M.; et al. From extraction to advanced analytical methods: The challenges of melanin analysis. Int. J. Mol. Sci. 2019, 20, 3943. [Google Scholar] [CrossRef]
- Lu, Y.; Ye, M.; Song, S.; Li, L.; Shaikh, F.; Li, J. Isolation, purification, and antiaging activity of melanin from Lachnum singerianum. Appl. Biochem. Biotechnol. 2014, 174, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Schmaler-Ripcke, J.; Sugareva, V.; Gebhardt, P.; Winkler, R.; Kniemeyer, O.; Heinekamp, T.; Brakhage, A.A. Production of pyomelanin, a second type of melanin, via the tyrosine degradation pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 2009, 75, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Vasanthakumar, A.; DeAraujo, A.; Mazurek, J.; Schilling, M.; Mitchell, R. Pyomelanin production in Penicillium chrysogenum is stimulated by L-tyrosine. Microbiology 2015, 161, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, K.; Streibel, M.; Jahn, B.; Gerhard Haase, G.; Brakhage, A.A. Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal Genet. Biol. 2003, 38, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Valiante, V.; Baldin, C.; Hortschansky, P.; Jain, R.; Thywißen, A.; Straßburger, M.; Shelest, E.; Heinekamp, T.; Brakhage, A.A. The Aspergillus fumigatus conidial melanin production is regulated by the bifunctional bHLH DevR and MADS-box RlmA transcription factors. Mol. Microbiol. 2016, 102, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Xu, X.; Lowry, D.; Jackson, J.C.; Roberson, R.W.; Lin, X. Subcellular compartmentalization and trafficking of the biosynthetic machinery for fungal melanin. Cell Rep. 2016, 14, 2511–2518. [Google Scholar] [CrossRef]
- Strycker, B.D.; Han, Z.; Bahari, A.; Pham, T.; Lin, X.; Shaw, B.D.; Sokolov, A.V.; Scully, M.O. Raman characterization of fungal DHN and DOPA melanin biosynthesis pathways. J. Fungi 2021, 7, 841. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Chow, S.; Tsé, K.K.; McClelland, E.; Casadevall, A. The effect of L-DOPA on Cryptococcus neoformans growth and gene expression. Virulence 2011, 2, 329–336. [Google Scholar] [CrossRef]
- Ribera, J.; Panzarasa, G.; Stobbe, A.; Osypova, A.; Rupper, P.; Klose, D.; Schwarze, F.W.M.R. Scalable biosynthesis of melanin by the Basidiomycete Armillaria cepistipes. J. Agric. Food Chem. 2019, 67, 132–139. [Google Scholar] [CrossRef]
- Ye, M.; Guo, G.; Lu, Y.; Song, S.; Wang, H.; Yang, L. Purification, structure and anti-radiation activity of melanin from Lachnum YM404. Int. J. Biol. Macromol. 2014, 63, 170–176. [Google Scholar] [CrossRef]
- Arun, G.; Eyini, M.; Gunasekaran, P. Characterization and biological activities of extracellular melanin produced by Schizophyllum commune (Fries). Indian J. Exp. Biol. 2015, 63, 380–387. [Google Scholar]
- Hu, H.L.; Dai, D.H.; Huang, G.R.; Zhang, Z.D. Isolation and characterization of extracellular melanin produced by Chroogomphus rutilus D447. Am. J. Food Technol. 2015, 10, 68–77. [Google Scholar] [CrossRef]
- Oh, J.-J.; Kim, Y.J.; Kim, J.Y.; Kwon, S.L.; Lee, C.; Lee, M.-E.; Kim, J.W.; Kim, G.-H. Genomic analysis and assessment of melanin synthesis in Amorphotheca resinae KUC3009. J. Fungi 2021, 7, 289. [Google Scholar] [CrossRef] [PubMed]
- Tudor, D.; Robinson, S.C.; Cooper, P.A. The influence of moisture content variation on fungal pigment formation in spalted wood. AMB Express 2012, 2, 69. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; de Menezes, G.C.A.; e Silva, T.R.; Bicas, J.L.; Oliveira, V.M.; Rosa, L.H. Antarctic fungi as producers of pigments. In Fungi of Antarctica; Rosa, L.H., Ed.; Springer Nature: Cham, Switzerland, 2019; pp. 305–318. [Google Scholar]
- Sajjad, W.; Din, G.; Rafiq, M.; Iqbal, A.; Khan, S.; Zada, S.; Ali, B.; Kang, S. Pigment production by cold-adapted bacteria and fungi: Colorful tale of cryosphere with wide range applications. Extremophiles 2020, 24, 447–473. [Google Scholar] [CrossRef]
- Selbmann, L.; Isola, D.; Zucconi, L.; Onofri, S. Resistance to UV-B induced DNA damage in extreme-tolerant cryptoendolithic Antarctic fungi: Detection by PCR assays. Fungal Biol. 2011, 115, 937–944. [Google Scholar] [CrossRef]
- Rosa, L.H.; Vaz, A.B.M.; Caligiorne, R.B.; Campolina, S.; Rosa, C.A. Endophytic fungi associated with the Antarctic grass Deschampsia antarctica Desv. (Poaceae). Polar Biol. 2009, 32, 161–167. [Google Scholar] [CrossRef]
- Pacelli, C.; Cassaro, A.; Maturilli, A.; Timperio, A.M.; Gevi, F.; Cavalazzi, B.; Stefan, M.; Ghica, D.; Onofri, S. Multidisciplinary characterization of melanin pigments from the black fungus Cryomyces antarcticus. Appl. Microbiol. Biotechnol. 2020, 104, 6385–6395. [Google Scholar] [CrossRef]
- de Menezes, G.C.A.; Godinho, V.M.; Porto, B.A.; Gonçalves, V.N.; Rosa, L.H. Antarctomyces pellizariae sp. nov., a new, endemic, blue, snow resident psychrophilic ascomycete fungus from Antarctica. Extremophiles 2017, 21, 259–269. [Google Scholar] [CrossRef]
- Gamze, N.M. Natural melanin synthesized by Aureobasidium pullulans using food wastes and its characterization. Appl. Food Biotechnol. 2021, 8, 307–318. [Google Scholar]
- Rana, B.; Bhattacharyya, M.; Patni, B.; Arya, M.; Joshi, G.K. The realm of microbial pigments in the food color market. Front. Sustain. Food Syst. 2021, 5, 603892. [Google Scholar] [CrossRef]
- Saini, A.S.; Melo, J.S. Biosorption of uranium by melanin: Kinetic, equilibrium and thermodynamic studies. Bioresour. Technol. 2013, 149, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Coelho, E.; Reis, T.A.; Cotrim, M.; Mullan, T.K.; Correa, B. Resistant fungi isolated from contaminated uranium mine in Brazil shows a high capacity to uptake uranium from water. Chemosphere 2020, 248, 126068. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Kim, J.Y.; Kim, Y.J.; Kim, S.; Kim, G. Utilization of extracellular fungal melanin as an eco-friendly biosorbent for treatment of metal-contaminated effluents. Chemosphere 2021, 272, 129884. [Google Scholar] [CrossRef] [PubMed]
- Manirethan, V.; Raval, K.; Rajan, R.; Thaira, H.; Balakrishnan, R.M. Kinetic and thermodynamic studies on the adsorption of heavy metals from aqueous solution by melanin nanopigment obtained from marine source: Pseudomonas stutzeri. J. Environ. Manag. 2018, 214, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Tran-Ly, A.N.; Ribera, J.; Schwarze, F.W.M.; Brunelli, M.; Giuseppino, F. Fungal melanin-based electrospun membranes for heavy metal detoxification of water. Sustain. Mater. Technol. 2020, 23, e00146. [Google Scholar] [CrossRef]
- Song, S.; Yang, L.; Ye, M.; Chen, X.; Shi, F.; Shaikh, F. Antioxidant activity of a Lachnum YM226 melanin–iron complex and its influence on cytokine production in mice with iron deficiency anemia. Food Funct. 2016, 7, 1508. [Google Scholar] [CrossRef]
- Schweitzer, A.D.; Revskaya, E.; Chu, P.; Pazo, V.; Friedman, M.; Nosanchuk, J.D.; Cahill, S.; Frases, S.; Casadevall, A.; Dadachova, E. Melanin-covered nanoparticles for protection of bone marrow during radiation therapy of cancer. Int. J. Rad. Oncol. Biol. Phys. 2010, 78, 1494–1502. [Google Scholar] [CrossRef]
- Ju, K.-Y.; Lee, Y.; Lee, S.; Park, S.B.; Lee, J.K. Bioinspired polymerization of dopamine to generate melanin-like nanoparticles having an excellent free-radical-scavenging property. Biomacromolecules 2011, 12, 625–632. [Google Scholar] [CrossRef]
- Araújo, M.; Viveiros, R.; Correia, T.R.; Correia, I.J.; Bonifácio, V.D.B.; Casimiro, T.; Aguiar-Ricardo, A. Natural melanin: A potential pH-responsive drug release device. Int. J. Pharm. 2014, 469, 140–145. [Google Scholar] [CrossRef]
- Chyizhanska, N.; Beregova, T. Effect of melanin isolated from Antarctic yeasts on preservation of pig livestock after ablactation. Ukrainian Antarctic J. 2009, 8, 382–385. [Google Scholar] [CrossRef]
- Dufossé, L.; Caro, Y.; Fouillaud, M. Fungal Pigments; MDPI: Basel, Switzerland, 2018; ISBN 978-3-03842-787-2. [Google Scholar]
- Mund, N.K.; Čellárová, E. Recent advances in the identification of biosynthetic genes and gene clusters of the polyketide-derived pathways for anthraquinone biosynthesis and biotechnological applications. Biotechnol. Adv. 2023, 63, 108104. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J.P. Production of fungal pigments: Molecular processes and their applications. J. Fungi 2023, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, Y.; Chen, M.; Fan, P.; Li, G.; Wang, C. Ammonium nitrate regulated the color characteristic changes of pigments in Monascus purpureus M9. AMB Expr. 2021, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Muangsin, N.; Wisetsakdakorn, W.; Chaichit, N.; Sihanonth, P.; Petsom, A.; Sangvanich, P. Austrocortinin: Crystal structure of a natural anthraquinone pigment from fungi. Dyes Pigments 2008, 77, 653–656. [Google Scholar] [CrossRef]
- Cui, Y.; Xia, C.-J.; Su, Y.-H.; Zhang, B.-Q.; Zhang, T.-T.; Liu, Y.; Hu, Z.-Y.; Tang, X.-J. Switching characteristics of anthraquinone molecular devices based on graphene electrodes. Acta Physica Sinica 2021, 70, 038501. [Google Scholar] [CrossRef]
- Giesbers, G.; Van Schenck, J.; Quinn, A.; Van Court, R.; Vega Gutierrez, S.M.; Robinson, S.C.; Ostroverkhova, O. Xylindein: Naturally produced fungal compound for sustainable (Opto)electronics. ACS Omega 2019, 4, 13309–13318. [Google Scholar] [CrossRef]
- Giesbers, G.; Krueger, T.; Schenck, J.V.; Court, R.V.; Moore, J.; Fang, C.; Robinson, S.C.; Ostroverkhova, O. Fungi-derived xylindein: Effect of purity on optical and electronic properties. MRS Adv. 2019, 4, 1769–1777. [Google Scholar] [CrossRef]
- Van Court, R.C.; Giesbers, G.; Ostroverkhova, O.; Robinson, S.C. Optimizing xylindein from Chlorociboria spp. for (Opto)electronic applications. Processes 2020, 8, 1477. [Google Scholar] [CrossRef]
- Khalid, S.; Keller, N.P. Chemical signals driving bacterial–fungal interactions. Environ. Microbiol. 2021, 23, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Spraker, J.E.; Wiemann, P.; Baccile, J.A.; Venkatesh, N.; Schumacher, J.; Schroeder, F.C. Conserved responses in a war of small molecules between a plant-pathogenic bacterium and fungi. MBio 2018, 9, e00820. [Google Scholar] [CrossRef] [PubMed]
- Moussa, M.; Ebrahim, W.; Bonus, M.; Gohlke, H.; Mandi, A.; Kurtan, T.; Hartmann, R.; Kalscheuer, R.; Lin, W.; Liu, Z.; et al. Co-culture of the fungus Fusarium tricinctum with Streptomyces lividans induces production of cryptic naphthoquinone dimers. RSC Adv. 2019, 9, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Akimenko, V.K. Naphthoquinone metabolites of the fungi. Phytochemistry 1998, 47, 935–959. [Google Scholar] [CrossRef] [PubMed]
- Dufossé, L. Microbial production of food grade pigments. Food Technol. Biotechnol. 2006, 44, 313–321. [Google Scholar]
- Yang, Y.; Liu, B.; Du, X.; Li, P.; Liang, B.; Cheng, X.; Du, L.; Huang, D.; Wang, L.; Wang, S. Complete genome sequence and transcriptomics analyses reveal pigment biosynthesis and regulatory mechanisms in an industrial strain, Monascus purpureus YY-1. Sci. Rep. 2015, 5, 8331. [Google Scholar] [CrossRef]
- Hajjaj, H.; Klaebe, A.; Goma, G.; Blanc, P.J.; Barbier, E.; Francois, J. Medium-chain fatty acids affect citrinin production in the filamentous fungus Monascus ruber. Appl. Environ. Microbiol. 2000, 66, 1120–1125. [Google Scholar] [CrossRef]
- Shi, K.; Song, D.; Chen, G.; Pistolozzi, M.; Wu, Z.; Quan, L. Controlling composition and color characteristics of Monascus pigments by pH and nitrogen sources in submerged fermentation. J. Biosci. Bioeng. 2015, 120, 145–154. [Google Scholar] [CrossRef]
- Mapari, S.A.S.; Meyer, A.S.; Thrane, U.; Frisvad, J.C. Identification of potentially safe promising fungal cell factories for the production of polyketide natural food colorants using chemotaxonomic rationale. Microb. Cell Fact. 2009, 8, 24. [Google Scholar] [CrossRef]
- Pisareva, E.; Savov, V.; Kujumdzieva, A. Pigments and citrinin biosynthesis by fungi belonging to genus Monascus. Z. Naturforsch. 2005, 60C, 116–120. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Xu, Y.; Li, Y.; Tan, W. Construction of a replacement vector to disrupt pksCT gene for the mycotoxin citrinin biosynthesis in Monascus aurantiacus and maintain food red pigment production. Asia Pac. J. Clin. Nutr. 2007, 16, 137–142. [Google Scholar]
- Shimizu, T.; Kinoshita, H.; Nihira, T. Identification and in vivo functional analysis by gene disruption of ctnA, an activator gene involved in citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2007, 73, 5097–5103. [Google Scholar] [CrossRef]
- Wan, Y.; Zhang, J.; Han, H.; Li, L.; Liu, Y.; Gao, M. Citrinin-producing capacity of Monascus purpureus in response to low-frequency magnetic fields. Process Biochem. 2017, 53, 25–29. [Google Scholar] [CrossRef]
- Koli, S.H.; Suryawanshi, R.K.; Patil, C.D.; Pati, S.V. Fluconazole treatment enhances extracellular release of red pigments in the fungus Monascus purpureus. FEMS Microbiol. Lett. 2017, 364, fnx058. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liu, X.; Wang, Y.; Xie, J.; Gao, H.; Li, X.; Huang, Z. Metabolomics analysis based on UHPLC-Q-TOF-MS/MS reveals effects of genistein on reducing mycotoxin citrinin production by Monascus aurantiacus Li AS3.4384. LWT-Food Sci. Technol. 2020, 130, 109613. [Google Scholar] [CrossRef]
- Liang, B.; Du, X.-J.; Li, P.; Sun, C.-C.; Wang, S. Investigation of citrinin and pigment biosynthesis mechanisms in Monascus purpureus by transcriptomic analysis. Front. Microbiol. 2018, 9, 1374. [Google Scholar] [CrossRef]
- Zhen, Z.; Xiong, X.; Liu, Y.; Zhang, J.; Wang, S.; Li, L.; Gao, M. NaCl inhibits citrinin and stimulates Monascus pigments and monacolin K production. Toxins 2019, 11, 118. [Google Scholar] [CrossRef]
- Chen, M.H.; Johns, M.R. Effect of pH and nitrogen source on pigment production by Monascus purpureus. Appl. Microbiol. Biotechnol. 1993, 40, 132–138. [Google Scholar] [CrossRef]
- Marcoleta, A.; Niklitschek, M.; Wozniak, A.; Lozano, C.; Alcaíno, J.; Baeza, M.; Cifuentes, V. Glucose and ethanol-dependent transcriptional regulation of the astaxanthin biosynthesis pathway in Xanthophyllomyces dendrorhous. BMC Microbiol. 2011, 11, 190. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Magalon, H.; Dufossé, L.; Fouillaud, M. Production of pigments from the tropical marine-derived fungi Talaromyces albobiverticillius: New resources for natural red-colored metabolites. J. Food Composit. Anal. 2018, 70, 35–48. [Google Scholar] [CrossRef]
- Egorova, A.S.; Gessler, N.N.; Belozerskaya, T.A. Melanin pigments in the fungus Paecilomyces lilacinus (thom) samson. Dokl Biochem. Biophys. 2011, 437, 84. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Assessing global fungal threats to humans. mLife 2022, 1, 223–240. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Class of Carotenoids | Species | Biosynthesis-Related Genes | References |
|---|---|---|---|
| β-Carotene | Blakeslea trispora | carRA, carB, carG | [12,13] |
| Xanthophyllomyces dendrorhous | crtE, crtI, crtYB | [11] | |
| Rhodotorula glutinis | crtI, crtYB | [14] | |
| Astaxanthin | Phaffia rhodozyma (syn. Xanthophyllomyces dendrorhous) | crtYB, crtS (also called Asy), crtI, crtE | [15] |
| Lycopene | Blakeslea trispora | carB, carRA | [16] |
| Type of Melanin Biosynthesis Pathway | Species | Biosynthesis -Related Genes | References |
|---|---|---|---|
| DHN–melanin | Aspergillus fumigatus | pksP/alb1, ayg1, arp1, arp2, abr1, abr2 | [47] |
| Aspergillus niger | alb1, ayg1, abr1, abr2 | [49] | |
| Aspergillus nidulans | ωA, yA | [50] | |
| Penicillium marneffei | alb1, ayg1, arp1, arp2, abr1, abr2 | [49] | |
| DOPA–melanin | Cryptococcus neoformans | Lac1, Lac2 | [51] |
| Pyomelanin | Aspergillus fumigatus | HppD, HmgA | [45] |
| Penicillium chrysogenum | HppD, HmgA | [46] |
| Class of Colorants/Paradigm | Fungal Species | Biosynthesis-Related Genes | References |
|---|---|---|---|
| Anthraquinones/endocrocin | Aspergillus fumigatus | encA, encB, encC | [77] |
| Naphthoquinone/bikaverin | Fusarium fujikuroi | Bik1, Bik2, Bik3, Bik4, Bik5, Bik6 | [78] |
| Azaphilone/Monascus pigments | Monascus purpureus | MppD, MpPKS5, MpFasB2, MpFasA2, MppF, MppA, MppB, MppE, Mpp7, MppC, MppG, MppR1, MppR2 | [79] |
| Categories of Colorants | Environmental Factors | References |
|---|---|---|
| Carotenoids | Light, temperature, oxygen, carbon source (i.e., glucose, ethanol), nitrogen, Ca++ availability | [24] |
| Melanins | UV irradiation, cold, desiccation, heavy-metal stress | [58,59] |
| Polyketide-derived pigments | Biotic stresses (i.e., other organisms), low-frequency magnetic field, NaCl, pH, NH4NO3 | [79,102,103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; Zhang, T.; Xu, J. Genetic and Environmental Factors Influencing the Production of Select Fungal Colorants: Challenges and Opportunities in Industrial Applications. J. Fungi 2023, 9, 585. https://doi.org/10.3390/jof9050585
Lin L, Zhang T, Xu J. Genetic and Environmental Factors Influencing the Production of Select Fungal Colorants: Challenges and Opportunities in Industrial Applications. Journal of Fungi. 2023; 9(5):585. https://doi.org/10.3390/jof9050585
Chicago/Turabian StyleLin, Lan, Tong Zhang, and Jianping Xu. 2023. "Genetic and Environmental Factors Influencing the Production of Select Fungal Colorants: Challenges and Opportunities in Industrial Applications" Journal of Fungi 9, no. 5: 585. https://doi.org/10.3390/jof9050585