
Autophagy-Related Gene 4 Participates in the Asexual Development, Stress Response and Virulence of Filamentous Insect Pathogenic Fungus Beauveria bassiana

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. Bioinformatic Analysis of BbAtg4
2.3. Targeted Gene Disruption and Complementation
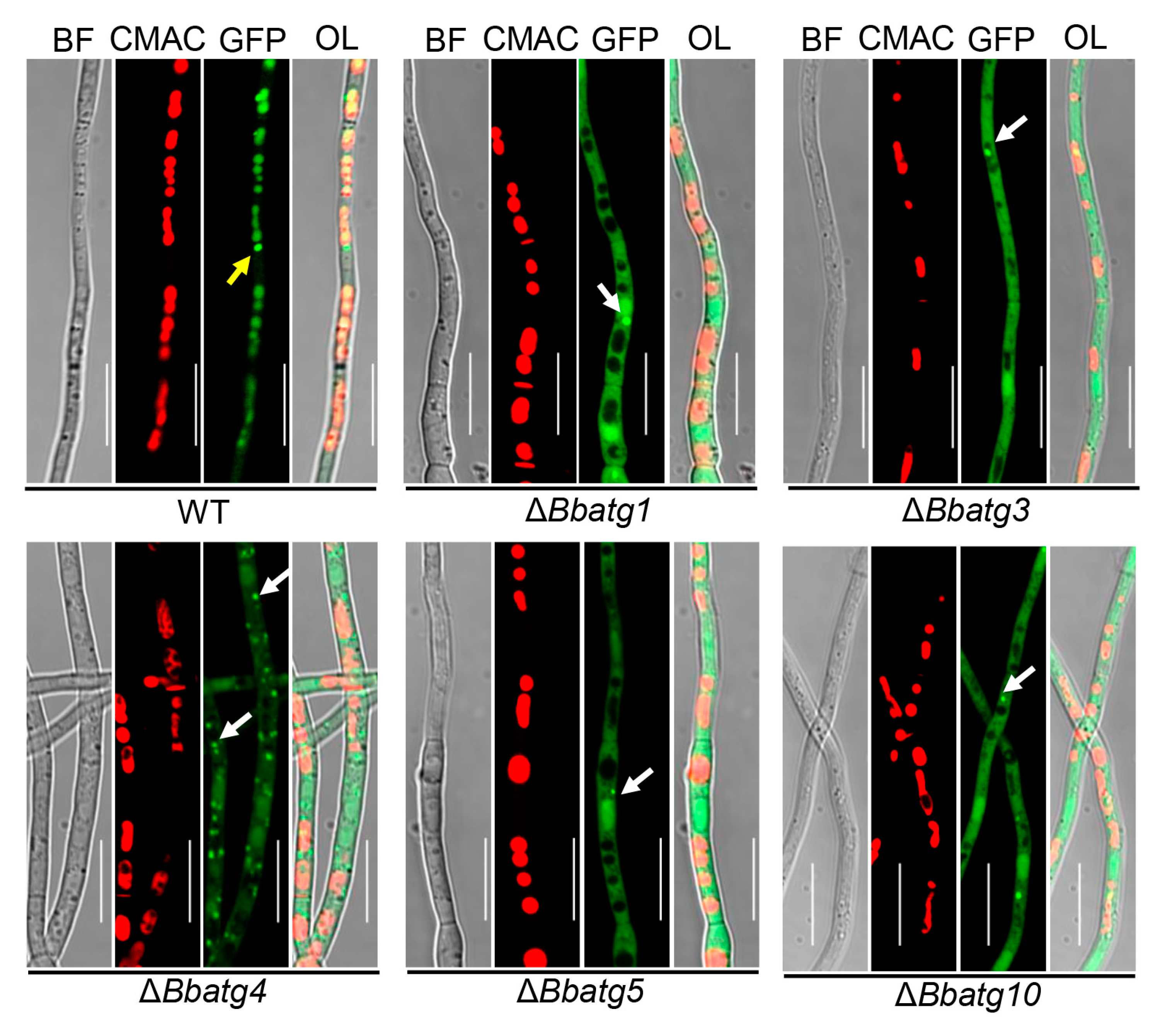
2.4. Visualizing Autophagic Flux in Fungal Strains
2.5. Phenotypic Assays
2.6. Insect Bioassay with Two Methods in Preparing Conidial Suspensioi5n
2.7. Statistical Analyses
3. Results
3.1. Characterization and Molecular Manipulation of BbAtg4
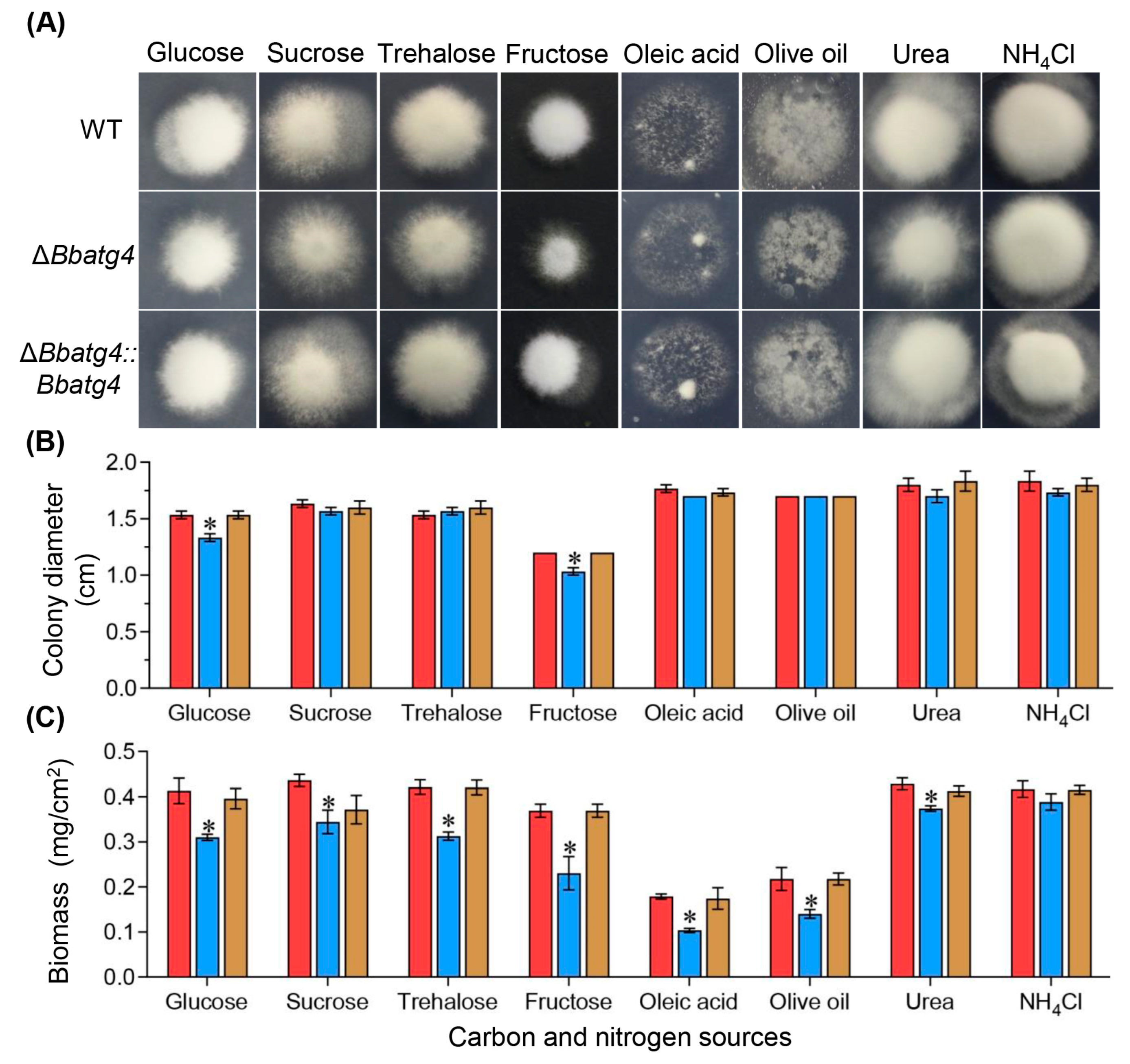
3.2. BbAtg4 Contributes to Vegetative Growth
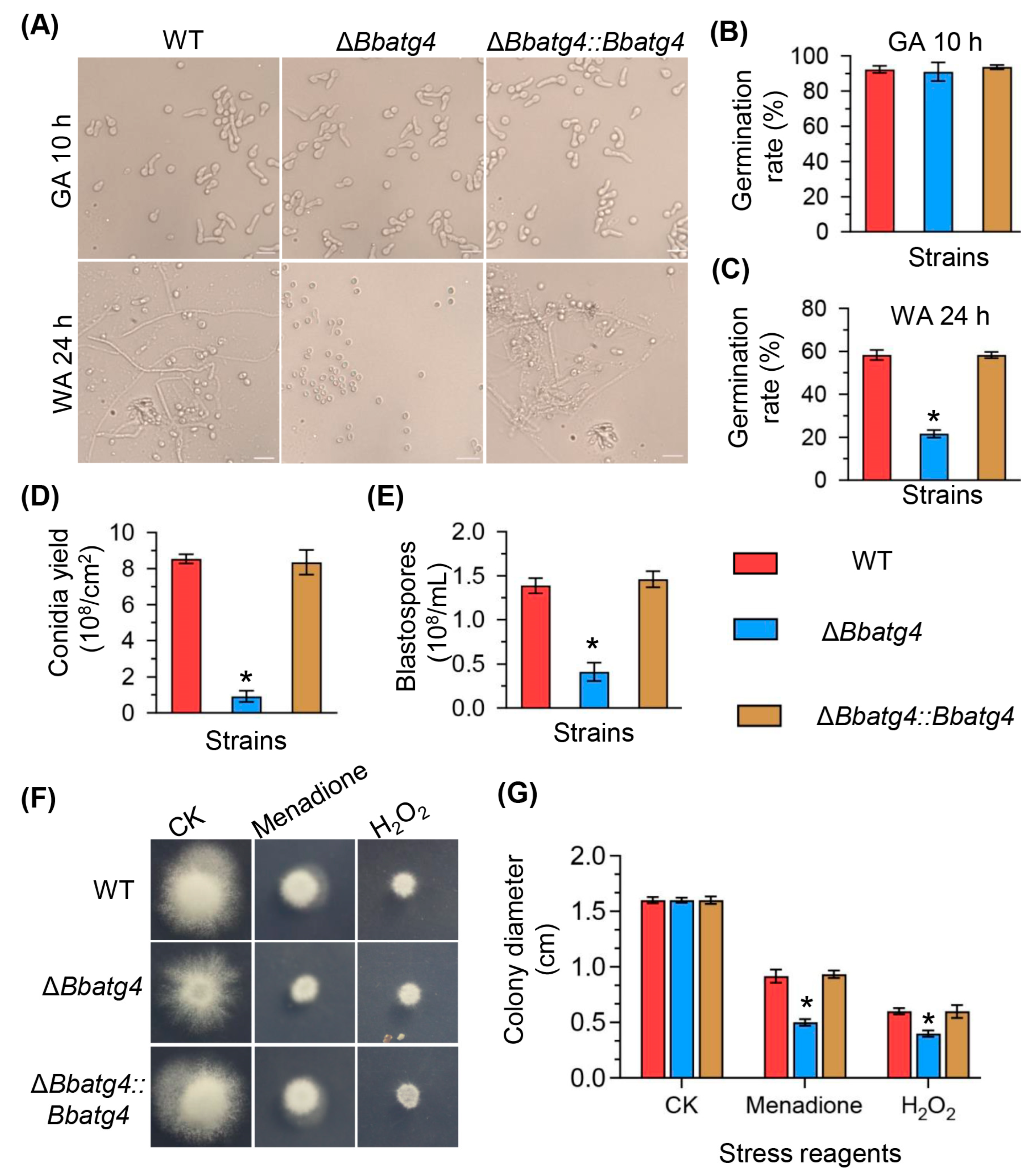
3.3. BbAtg4 Is Required for Conidial Germination under Nutrient-Limitation Condition
3.4. BbAtg4 Is Involved in Fungal Development
3.5. BbAtg4 Contributes to Fungal Resistance to Oxidative Stress
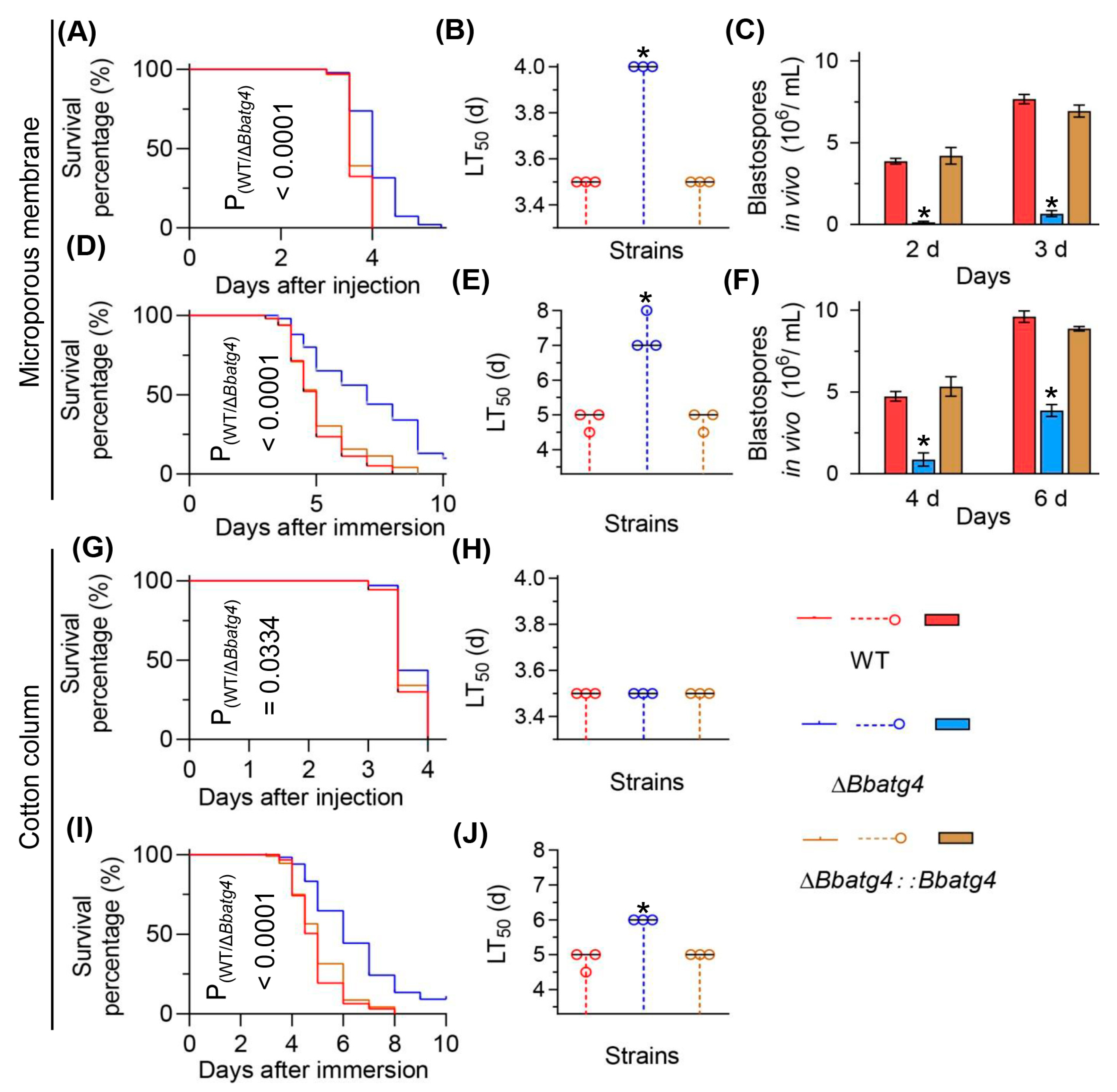
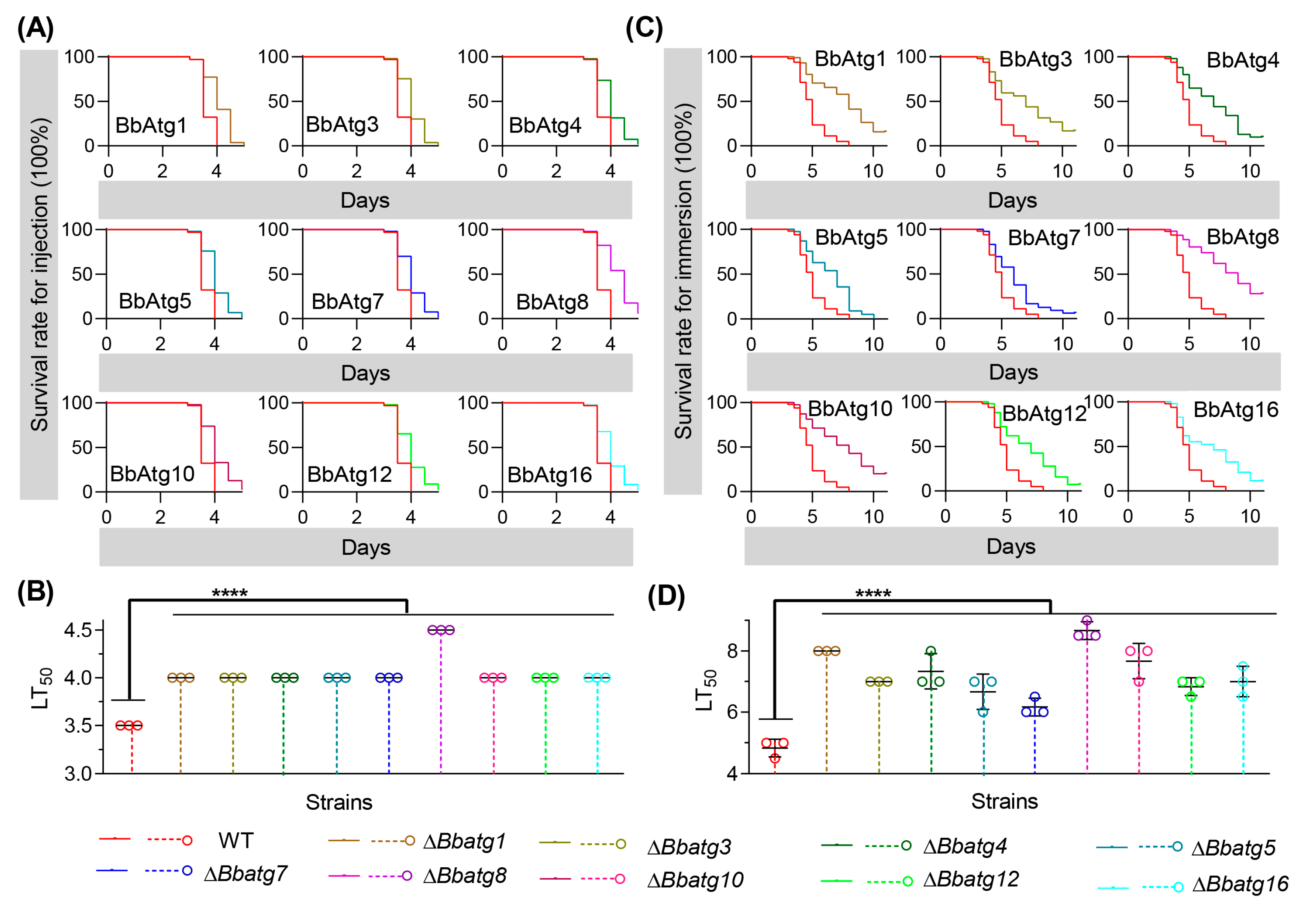
3.6. BbAtg4 Is Important to Fungal Virulence
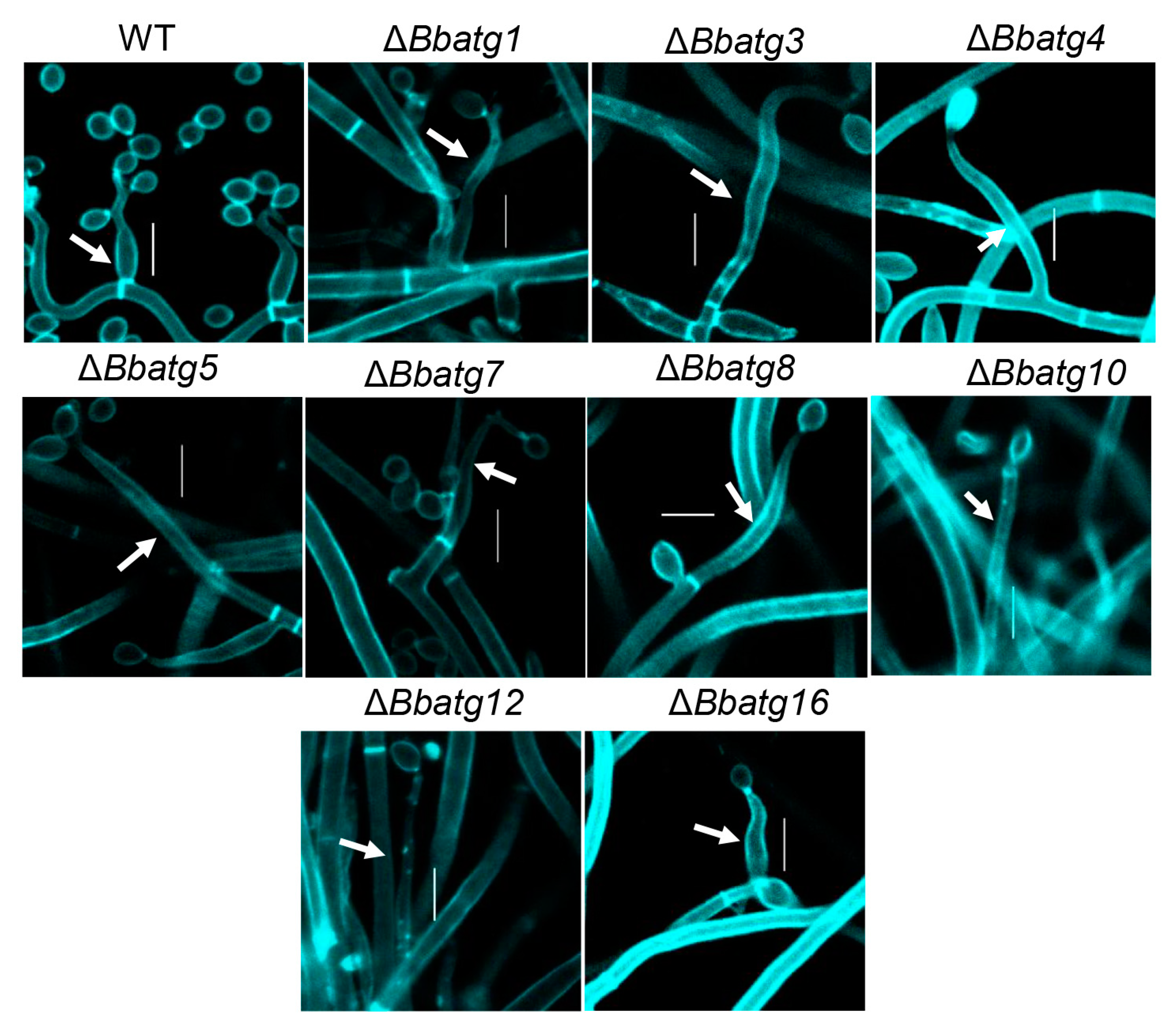
3.7. Autophagy Is Crucial for the Differentiation of Spore-Formation Structures
3.8. Re-Examine the Virulence of Autophagy-Null Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Cruz, Q.R.; Roussos, S.; Hernandez, D.; Rodriguez, R.; Castillo, F.; Aguilar, C.N. Challenges and opportunities of the biopesticides production by solid-state fermentation: Filamentous fungi as a model. Crit. Rev. Biotechnol. 2015, 35, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Maistrou, S.; Natsopoulou, M.E.; Jensen, A.B.; Meyling, N.V. Virulence traits within a community of the fungal entomopathogen Beauveria: Associations with abundance and distribution. Fungal Ecol. 2020, 48, 100992. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, J.; Xie, X.Q.; Keyhani, N.O.; Feng, M.G.; Ying, S.H. The autophagy gene BbATG5, involved in the formation of the autophagosome, contributes to cell differentiation and growth but is dispensable for pathogenesis in the entomopathogenic fungus Beauveria bassiana. Microbiology 2013, 159, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Wanchoo, A.; Lewis, M.W.; Keyhani, N.O. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria bassiana. Microbiology 2009, 155, 3121–3133. [Google Scholar] [CrossRef]
- Ding, J.L.; Hou, J.; Feng, M.G.; Ying, S.H. Transcriptomic analyses reveal comprehensive responses of insect hemocytes to mycopathogen Beauveria bassiana, and fungal virulence-related cell wall protein assists pathogen to evade host cellular defense. Virulence 2020, 11, 1352–1365. [Google Scholar] [CrossRef]
- Meyling, N.V.; Pell, J.K.; Eilenberg, J. Dispersal of Beauveria bassiana by the activity of nettle insects. J. Invertebr. Pathol. 2006, 93, 121–126. [Google Scholar] [CrossRef]
- Farré, J.C.; Subramani, S. Mechanistic insights into selective autophagy pathways: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2016, 17, 537–552. [Google Scholar] [CrossRef]
- Ying, S.H.; Liu, J.; Chu, X.L.; Xie, X.Q.; Feng, M.G. The autophagy-related genes BbATG1 and BbATG8 have different functions in differentiation, stress resistance and virulence of mycopathogen Beauveria bassiana. Sci. Rep. 2016, 6, 26376. [Google Scholar] [CrossRef]
- Meijer, W.H.; van der Klei, I.J.; Veenhuis, M.; Kiel, J.A.K.W. ATG genes involved in non-selective autophagy are conserved from yeast to man, but the selective Cvt and pexophagy pathways also require organism-specific genes. Autophagy 2007, 3, 106–116. [Google Scholar] [CrossRef]
- Nakatogawa, H. Mechanisms governing autophagosome biogenesis. Nat. Rev. Mol. Cell Biol. 2020, 21, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Kikuma, T.; Kitamoto, K. Analysis of autophagy in Aspergillus oryzae by disruption of Aoatg13, Aoatg4, and Aoatg15 genes. FEMS Microbiol. Lett. 2011, 316, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ren, W.; Li, F.; Chen, C.; Ma, Z. Involvement of the cysteine protease BcAtg4 in development and virulence of Botrytis cinerea. Curr. Genet. 2019, 65, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Wang, C.; Yang, N.; Que, Y.; Talbot, N.J.; Wang, Z. Genome-wide functional analysis reveals that autophagy is necessary for growth, sporulation, deoxynivalenol production and virulence in Fusarium graminearum. Sci. Rep. 2017, 7, 11062. [Google Scholar] [CrossRef]
- Kershaw, M.J.; Talbot, N.J. Genome-wide functional analysis reveals that infection-associated fungal autophagy is necessary for rice blast disease. Proc. Natl. Acad. Sci. USA 2009, 106, 15967–15972. [Google Scholar] [CrossRef]
- Duan, Z.; Chen, Y.; Huang, W.; Shang, Y.; Chen, P.; Wang, C. Linkage of autophagy to fungal development, lipid storage and virulence in Metarhizium robertsii. Autophagy 2013, 9, 538–549. [Google Scholar] [CrossRef]
- Ding, J.L.; Lin, H.Y.; Feng, M.G.; Ying, S.H. Mbp1, a component of the MluI cell cycle box-binding complex, contributes to morphological transition and virulence in the filamentous entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2020, 22, 584–597. [Google Scholar] [CrossRef]
- Wang, J.J.; Peng, Y.J.; Ding, J.L.; Feng, M.G.; Ying, S.H. Mitochondrial fission is necessary for mitophagy, development and virulence of the insect pathogenic fungus Beauveria bassiana. J. Appl. Microbiol. 2020, 129, 411–421. [Google Scholar] [CrossRef]
- Guo, N.; Qian, Y.; Zhang, Q.; Chen, X.; Zeng, G.; Zhang, X.; Mi, W.; Xu, C.; St Leger, R.J.; Fang, W. Alternative transcription startsite selection in Mr-OPY2 controls lifestyle transitions in the fungus Metarhizium robertsii. Nat. Commun. 2017, 8, 1565. [Google Scholar] [CrossRef]
- Ding, J.L.; Li, X.H.; Lei, J.H.; Feng, M.G.; Ying, S.H. Succinate dehydrogenase subunit C contributes to mycelial growth and development, stress response, and virulence in the insect parasitic fungus Beauveria bassiana. Microbiol. Spectr. 2022, 10, e0289122. [Google Scholar] [CrossRef]
- Ding, J.L.; Lin, H.Y.; Hou, J.; Feng, M.G.; Ying, S.H. The entomopathogenic fungus Beauveria bassiana employs autophagy as a persistence and recovery mechanism during conidial dormancy. mBio 2023, 14, e03049-22. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Hou, J.; Zhang, H.; Lei, J.H.; Lin, H.Y.; Ding, J.L.; Feng, M.G.; Ying, S.H. Systematic contributions of CFEM domain-containing proteins to iron acquisition are essential for interspecies interaction of the filamentous pathogenic fungus Beauveria bassiana. Environ. Microbiol. 2022, 24, 3693–3704. [Google Scholar] [CrossRef]
- Wang, F.; Sethiya, P.; Hu, X.; Guo, S.; Chen, Y.; Li, A.; Tan, K.; Wong, K.H. Transcription in fungal conidia before dormancy produces phenotypically variable conidia that maximize survival in different environments. Nat. Microbiol. 2021, 6, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Ding, J.L.; Peng, Y.J.; Feng, M.G.; Ying, S.H. Proteomic and phosphoryproteomic investigations reveal that autophagy-related protein 1, a protein kinase for autophagy initiation, synchronously deploys phosphoregulation on the ubiquitin-like conjugation system in the mycopathogen Beauveria bassiana. mSystems 2022, 22, e0146321. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Wang, J.J.; Feng, M.G.; Ying, S.H. Autophagy-related gene ATG7 participates in the asexual development, stress response and virulence of filamentous insect pathogenic fungus Beauveria bassiana. Curr. Genet. 2019, 65, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, J.J.; Lin, H.Y.; Feng, M.G.; Ying, S.H. Roles of autophagy-related genes in conidiogenesis and blastospore formation, virulence, and stress response of Beauveria bassiana. Fungal Biol. 2020, 124, 1052–1057. [Google Scholar] [CrossRef]
- Ding, J.L.; Zhang, H.; Feng, M.G.; Ying, S.H. Divergent physiological functions of four Atg22-like proteins in conidial germination, development, and virulence of the entomopathogenic fungus Beauveria bassiana. J. Fungi 2023, 9, 262. [Google Scholar] [CrossRef]
- Deng, Y.Z.; Qu, Z.; Naqvi, N.I. Role of macroautophagy in nutrient homeostasis during fungal development and pathogenesis. Cells 2012, 1, 449–463. [Google Scholar] [CrossRef]
- Mizushima, N. The ATG conjugation systems in autophagy. Curr. Opin. Cell Biol. 2020, 63, 1–10. [Google Scholar] [CrossRef]
- Pollack, J.K.; Harris, S.D.; Marten, M.R. Autophagy in filamentous fungi. Fungal Genet. Biol. 2009, 46, 1–8. [Google Scholar] [CrossRef]
- Ying, S.H.; Feng, M.G. Insight into vital role of autophagy in sustaining biological control potential of fungal pathogens against pest insects and nematodes. Virulence 2019, 10, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.J.; Codogno, P. AMP-activated protein kinase and autophagy. Autophagy 2007, 3, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.L.; Peng, Y.J.; Chu, X.L.; Feng, M.G.; Ying, S.H. Autophagy-related gene BbATG11 is indispensable for pexophagy and mitophagy, and contributes to stress response, conidiation and virulence in the insect mycopathogen Beauveria bassiana. Environ. Microbiol. 2018, 20, 3309–3324. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K.; Debnath, J. Beyond self-eating: The control of non-autophagic functions and signaling pathways by autophagy-related proteins. J. Cell Biol. 2017, 217, 813–822. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Injection Infection | Topical Infection | ||
|---|---|---|---|---|
| χ2 Value | p Value | χ2 Value | p Value | |
| ΔBbatg1 | 57.38 | <0.0001 | 61.33 | <0.0001 |
| ΔBbatg3 | 49.93 | <0.0001 | 29.54 | <0.0001 |
| ΔBbatg4 | 44.54 | <0.0001 | 44.02 | <0.0001 |
| ΔBbatg5 | 49.62 | <0.0001 | 29.24 | <0.0001 |
| ΔBbatg7 | 40.10 | <0.0001 | 18.75 | <0.0001 |
| ΔBbatg8 | 74.79 | <0.0001 | 99.63 | <0.0001 |
| ΔBbatg10 | 47.54 | <0.0001 | 60.45 | <0.0001 |
| ΔBbatg12 | 31.18 | <0.0001 | 36.15 | <0.0001 |
| ΔBbatg16 | 37.12 | <0.0001 | 23.89 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.-L.; Wei, K.; Feng, M.-G.; Ying, S.-H. Autophagy-Related Gene 4 Participates in the Asexual Development, Stress Response and Virulence of Filamentous Insect Pathogenic Fungus Beauveria bassiana. J. Fungi 2023, 9, 543. https://doi.org/10.3390/jof9050543
Ding J-L, Wei K, Feng M-G, Ying S-H. Autophagy-Related Gene 4 Participates in the Asexual Development, Stress Response and Virulence of Filamentous Insect Pathogenic Fungus Beauveria bassiana. Journal of Fungi. 2023; 9(5):543. https://doi.org/10.3390/jof9050543
Chicago/Turabian StyleDing, Jin-Li, Kang Wei, Ming-Guang Feng, and Sheng-Hua Ying. 2023. "Autophagy-Related Gene 4 Participates in the Asexual Development, Stress Response and Virulence of Filamentous Insect Pathogenic Fungus Beauveria bassiana" Journal of Fungi 9, no. 5: 543. https://doi.org/10.3390/jof9050543