
Disentangling the Taxonomy, Systematics, and Life History of the Spider-Parasitic Fungus Gibellula (Cordycipitaceae, Hypocreales)
 , , , and
, , , and 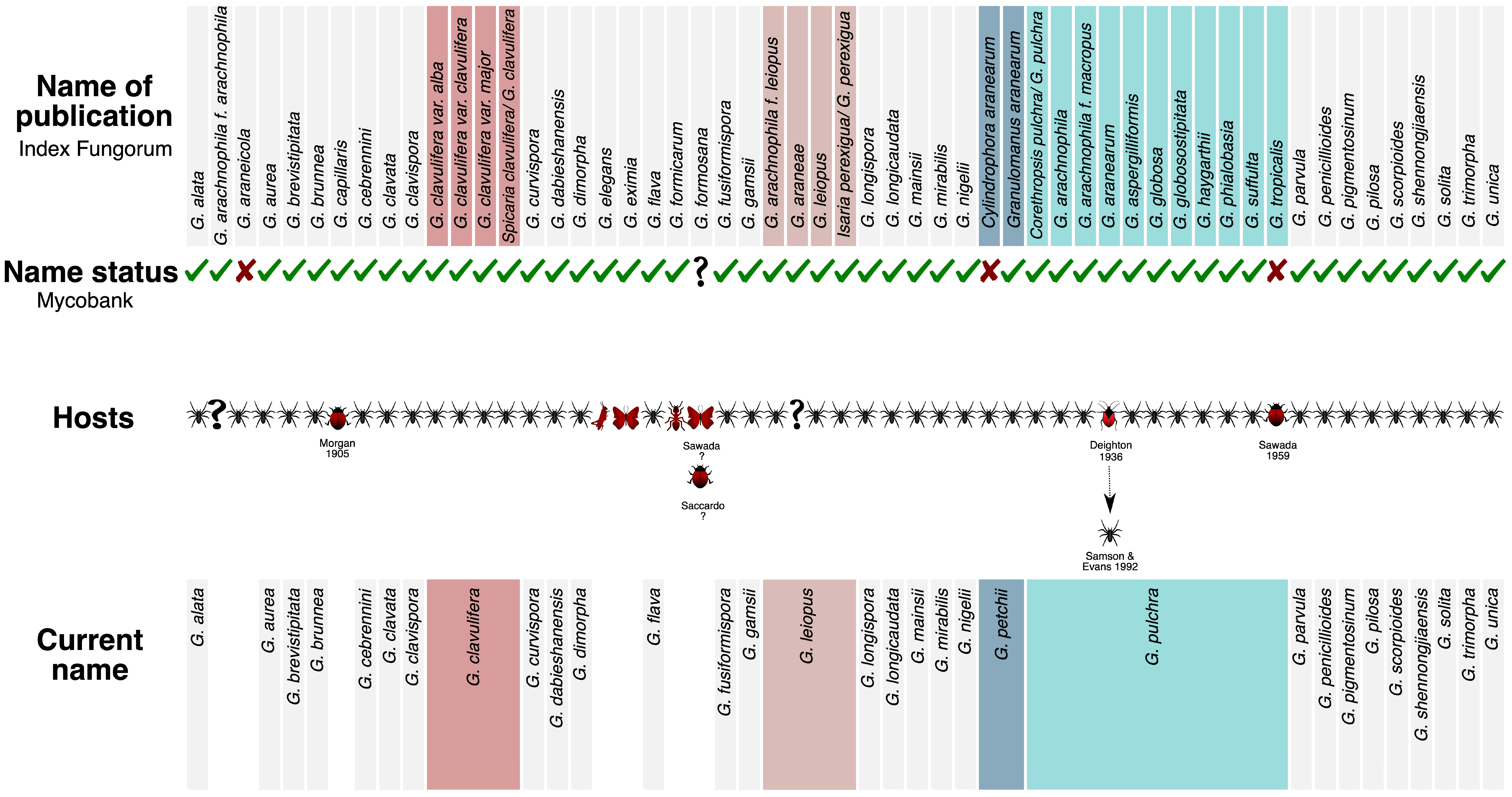{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Systematic Review Approach
2.1.1. Eligibility Criteria and Information Sources
2.1.2. Search Strategy, Study Selection, and Data Collection
2.1.3. Data Items and Summary Measures
2.2. Fieldwork Areas and Collection of Under-Represented Species
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Analyses of Morphological Traits
2.5. Multigene Phylogenetic Analyses
2.5.1. Obtaining Molecular Data
2.5.2. Phylogenetic Reconstruction
2.6. Global Distribution
2.7. Host–Parasite Interaction
3. Results
3.1. Systematic Review Approach
3.2. Indexation, Validity, and Currently Accepted Species
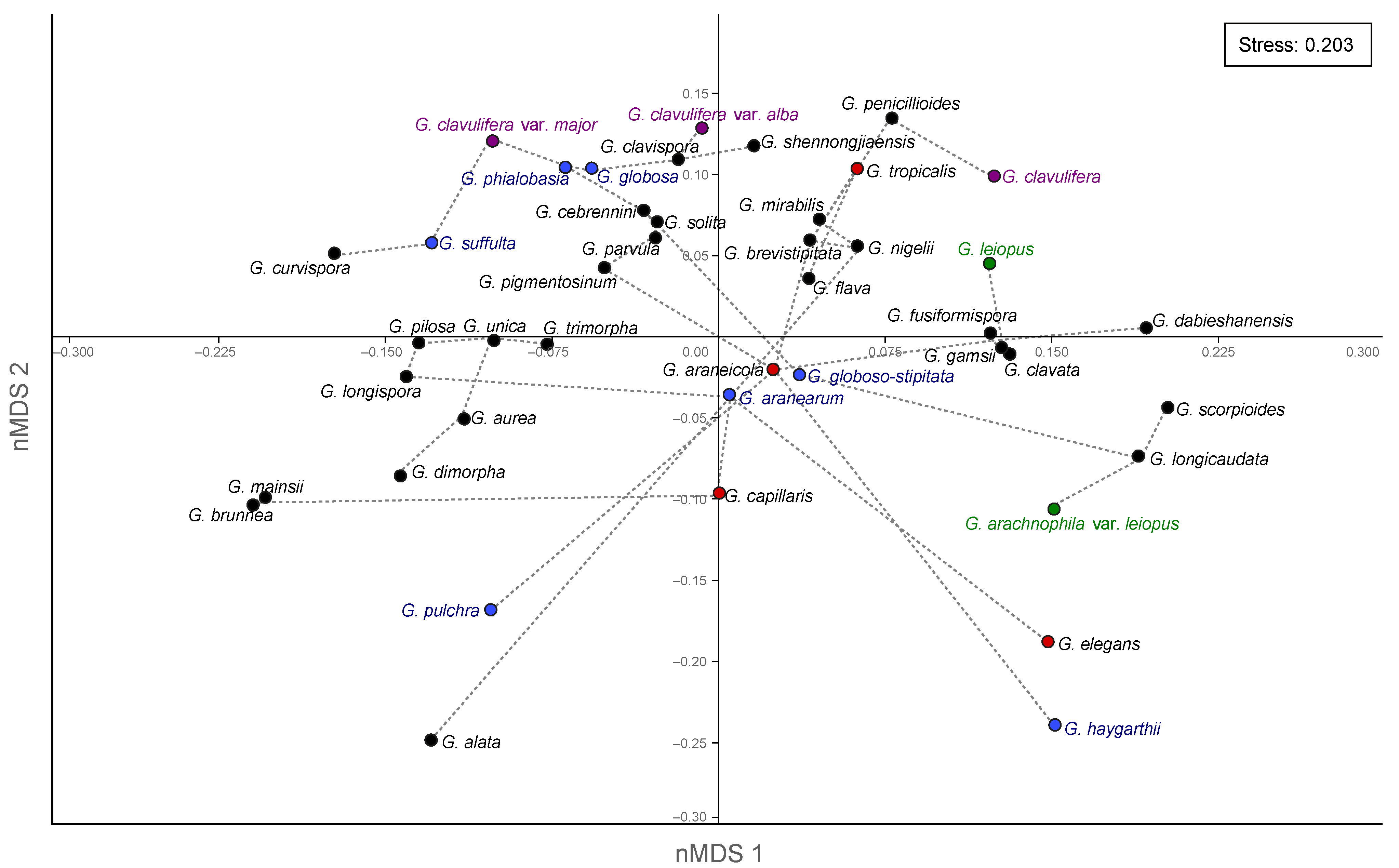
3.3. Similarities among Morphological Traits
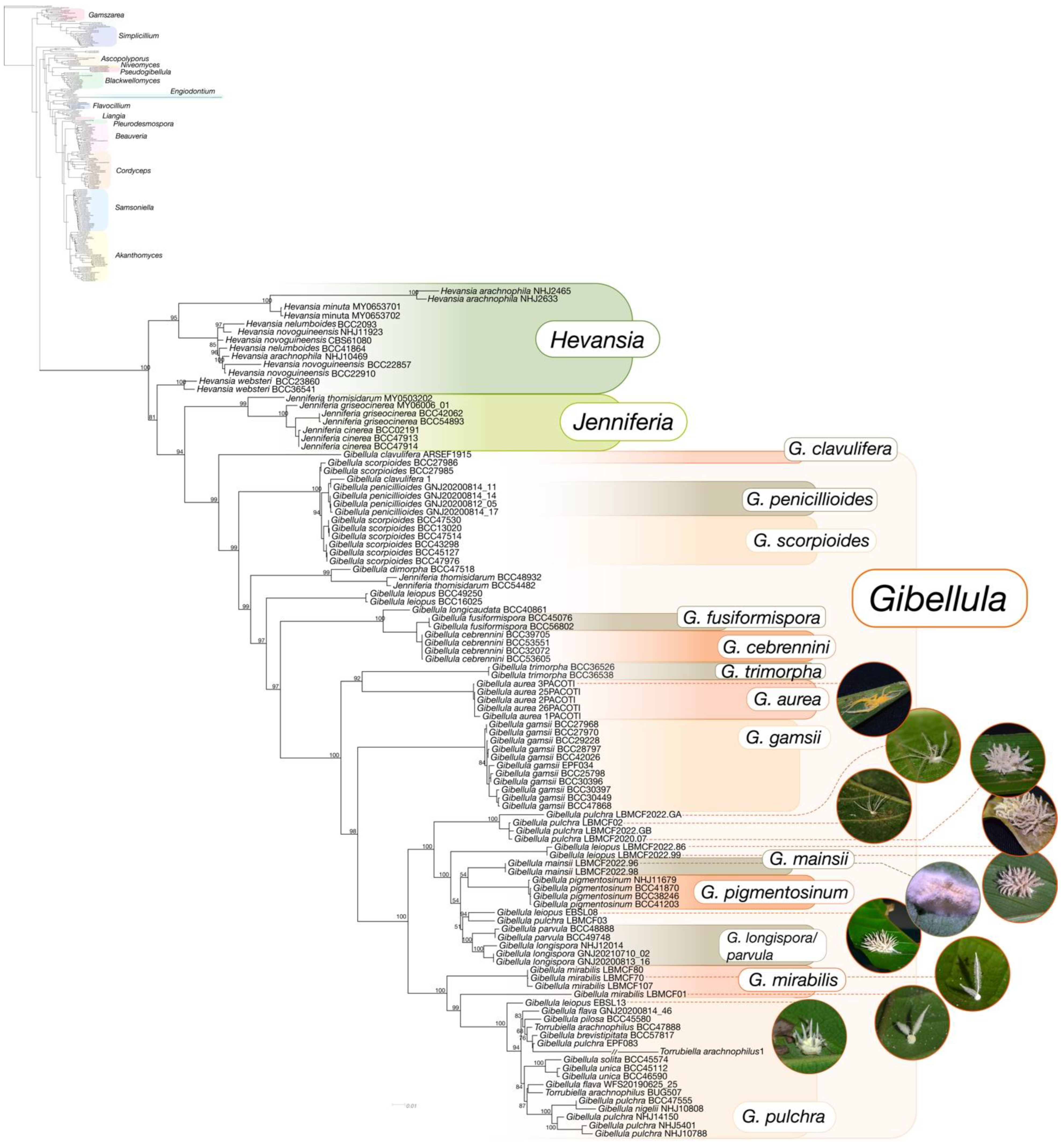
3.4. Multigene Phylogenetic Analyses
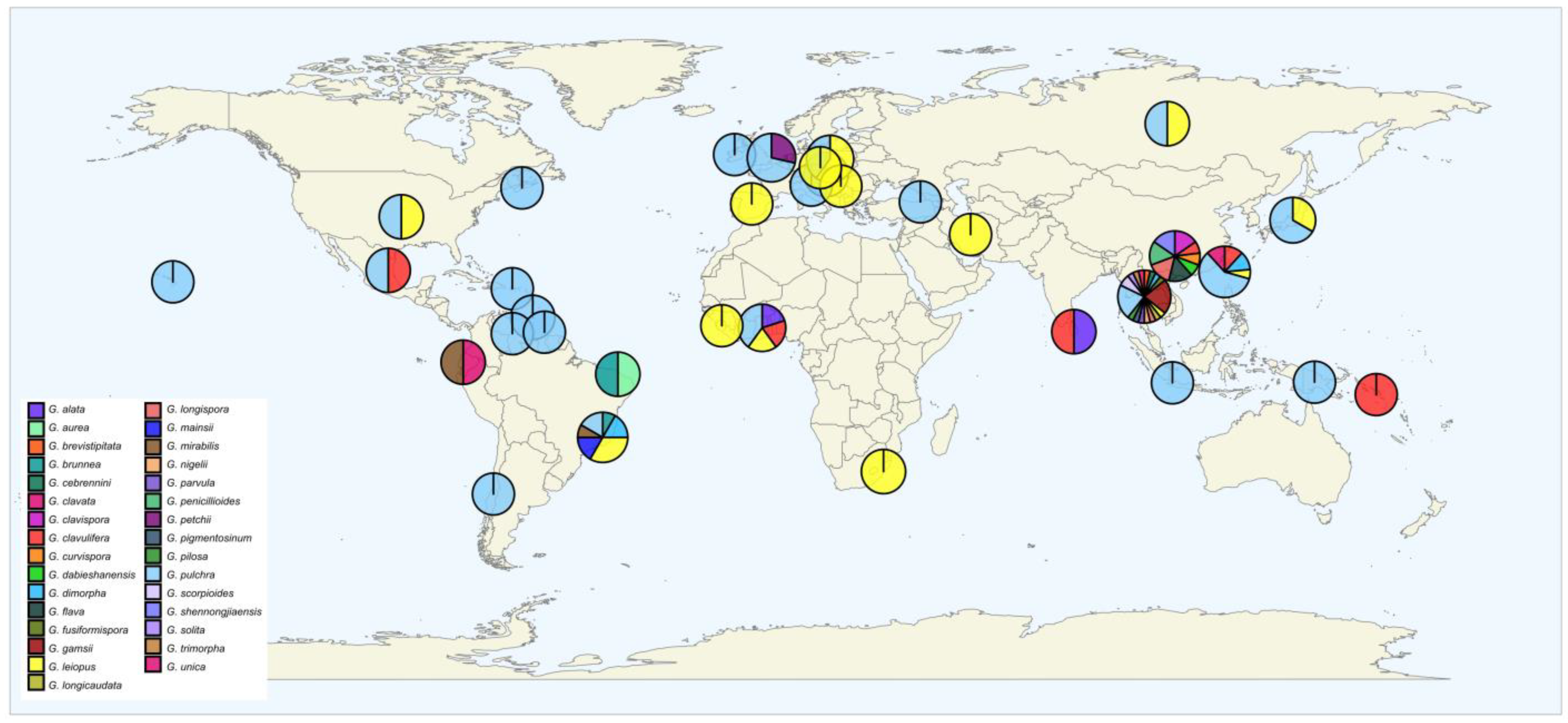
3.5. Global Distribution
3.6. Host–Parasite Interaction
4. Discussion
4.1. Currently Accepted Species and Morphological Traits
4.2. Multigene Phylogeny
4.3. Global Distribution
4.4. Host–Parasite Interaction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hywel-Jones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. The Life of a Dead Ant: The Expression of an Adaptive Extended Phenotype. Am. Nat. 2009, 174, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.P.; Araújo, J.P.M.; Loreto, R.G.; Quevillon, L.; de Bekker, C.; Evans, H.C. From so Simple a Beginning: The Evolution of Behavioral Manipulation by Fungi. Adv. Genet. 2016, 94, 437–469. [Google Scholar] [PubMed]
- Samson, R.A.; Evans, H.C.; Latge, J.-P. Atlas of Entomopathogenic Fungi; Springer: Berlin/Heidelberg, Germany, 1988; ISBN 978-3-662-05890-9. [Google Scholar]
- Poulin, R. Parasite Manipulation of Host Behavior: An Update and Frequently Asked Questions. In Advances in the Study of Behavior; Elsevier A cademic Press: Cambridge, MA, USA, 2010; Volume 41, pp. 151–186. ISBN 978-0-12-380892-9. [Google Scholar]
- Costa, P.P. Gibellula spp. Associadas a Aranhas Da Mata Do Paraíso, Viçosa-MG. Master’s Thesis, Universidade Federal de Viçosa, Minas Gerais, Brasil, 2014. [Google Scholar]
- Araújo, J.P.M.; Hughes, D.P. Diversity of Entomopathogenic Fungi: Which Groups Conquered the Insect Body? In Advances in Genetics; Elsevier Academic Press: Cambridge, MA, USA, 2016; Volume 94, pp. 1–39. ISBN 978-0-12-804694-4. [Google Scholar]
- Pontoppidan, M.-B.; Himaman, W.; Hywel-Jones, N.L.; Boomsma, J.J.; Hughes, D.P. Graveyards on the Move: The Spatio-Temporal Distribution of Dead Ophiocordyceps-Infected Ants. PLoS ONE 2009, 4, e4835. [Google Scholar] [CrossRef] [Green Version]
- Araújo, J.P.M.; Hughes, D.P. Zombie-Ant Fungi Emerged from Non-Manipulating, Beetle-Infecting Ancestors. Curr. Biol. 2019, 29, 3735–3738. [Google Scholar] [CrossRef]
- Araújo, J.P.M.; Evans, H.C.; Kepler, R.; Hughes, D.P. Zombie-Ant Fungi across Continents: 15 New Species and New Combinations within Ophiocordyceps. I. Myrmecophilous Hirsutelloid Species. Stud. Mycol. 2018, 90, 119–160. [Google Scholar] [CrossRef]
- Somavilla, A.; Barbosa, B.C.; Prezoto, F.; Oliveira, M.L. Infection and Behavior Manipulation of Social Wasps (Vespidae: Polistinae) by Ophiocordyceps humbertii in Neotropical Forests: New Records of Wasp-Zombification by a Fungus. Stud. Neotrop. Fauna Environ. 2020, 55, 23–28. [Google Scholar] [CrossRef]
- Samson, R.A.; Evans, H.C. Notes on Entomogenous Fungi From Ghana: II. The Genus Akanthomyces. Acta Bot. Neerl. 1974, 23, 28–35. [Google Scholar] [CrossRef]
- Sánchez-Peña, S.R. Some Insect and Spider Pathogenic Fungi from Mexico with Data on Their Host Ranges. Fla. Entomol. 1990, 73, 517. [Google Scholar] [CrossRef]
- Cooley, J.R.; Marshall, D.C.; Hill, K.B.R. A Specialized Fungal Parasite (Massospora cicadina) Hijacks the Sexual Signals of Periodical Cicadas (Hemiptera: Cicadidae: Magicicada). Sci. Rep. 2018, 8, 1432. [Google Scholar] [CrossRef] [PubMed]
- Arruda, I.D.P.; Villanueva-Bonilla, G.A.; Faustino, M.L.; Moura-Sobczak, J.C.M.S.; Sobczak, J.F. Behavioral Manipulation of the Spider Macrophyes pacoti (Araneae: Anyphaenidae) by the Araneopathogenic Fungus Gibellula sp. (Hypocreales: Cordycipitaceae). Can. J. Zool. 2021, 99, 401–408. [Google Scholar] [CrossRef]
- Mendes-Pereira, T.; Machado De Araújo, J.P.; Carvalho Mendes, F.; Oliveira Fonseca, E.; Erica Da Rocha Alves, J.; Fernando Sobczak, J.; Góes-Neto, A. Gibellula aurea sp. nov. (Ascomycota, Cordycipitaceae): A New Golden Spider-Devouring Fungus from a Brazilian Atlantic Rainforest. Phytotaxa 2022, 573, 85–102. [Google Scholar] [CrossRef]
- Evans, H.C. Fungal Pathogens of Spiders. In Spider Ecophysiology; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 107–121. ISBN 978-3-642-33988-2. [Google Scholar]
- Evans, H.C. Natural Control of Arthropods, with Special Reference to Ants (Formicidae), by Fungi in the Tropical High Forest of Ghana. J. Appl. Ecol. 1974, 11, 37–49. [Google Scholar] [CrossRef]
- Kuephadungphan, W.; Tasanathai, K.; Petcharad, B.; Khonsanit, A.; Stadler, M.; Luangsa-ard, J.J. Phylogeny- and Morphology-Based Recognition of New Species in the Spider-Parasitic Genus Gibellula (Hypocreales, Cordycipitaceae) from Thailand. MycoKeys 2020, 72, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Mains, E.B. The Genus Gibellula on Spiders in North America. Mycologia 1950, 42, 306–321. [Google Scholar] [CrossRef]
- Samson, R.A.; Evans, H.C. New Species of Gibellula on Spiders (Araneida) from South America. Mycologia 1992, 84, 300–314. [Google Scholar] [CrossRef]
- Vieira, C.; Japyassú, H.F.; Santos, A.J.; Gonzaga, M.O. Teias e Forrageamento. In Ecologia e Comportamento de Aranhas; Gonzaga, M.O., Santos, A.J., Japyassú, H.F., Eds.; Interciência: Rio de Janeiro, Brazil, 2007; pp. 45–65. [Google Scholar]
- Sung, G.-H.; Hywel-Jones, N.L.; Sung, J.-M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic Classification of Cordyceps and the Clavicipitaceous Fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [Green Version]
- Kobayasi, Y.; Shimizu, D. Some Species of Cordyceps and Its Allies on Spiders. Kew Bull. 1977, 31, 557–566. [Google Scholar] [CrossRef]
- Tzean, S.S.; Hsieh, L.S.; Wu, W.J. The Genus Gibellula on Spiders from Taiwan. Mycologia 1997, 89, 309–318. [Google Scholar] [CrossRef]
- Johnson, D.; Sung, G.-H.; Hywel-Jones, N.L.; Luangsa-Ard, J.J.; Bischoff, J.F.; Kepler, R.M.; Spatafora, J.W. Systematics and Evolution of the Genus Torrubiella (Hypocreales, Ascomycota). Mycol. Res. 2009, 113, 279–289. [Google Scholar] [CrossRef]
- Turland, N.; Wiersema, J.; Barrie, F.; Greuter, W.; Hawksworth, D.; Herendeen, P.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants; Regnum Vegetabile; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume 159, ISBN 978-3-946583-16-5. [Google Scholar]
- Kepler, R.M.; Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.-H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A Phylogenetically-Based Nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavara, F. Ulteriore Contribuzione Alla Micologia Lombarda. In Atti dell’Istituto Botanico della Università e Laboratorio Crittogamico di Pavia; Università di Pavia: Pavia, Italy, 1894; Volume 3, pp. 313–350. [Google Scholar]
- Index Fungorum Index Fungorum. Available online: https://www.indexfungorum.org (accessed on 30 November 2022).
- Mycobank MYCOBANK Database—Fungal Databases, Nomenclature & Species Banks. Available online: https://www.mycobank.org/ (accessed on 30 November 2022).
- Shrestha, B.; Kubátová, A.; Tanaka, E.; Oh, J.; Yoon, D.-H.; Sung, J.-M.; Sung, G.-H. Spider-Pathogenic Fungi within Hypocreales (Ascomycota): Their Current Nomenclature, Diversity, and Distribution. Mycol. Prog. 2019, 18, 983–1003. [Google Scholar] [CrossRef]
- Petch, T. A List of the Entomogenous Fungi of Great Britain. Trans. Br. Mycol. Soc. 1932, 17, 170–178. [Google Scholar] [CrossRef]
- Chen, W.-H.; Han, Y.-F.; Liang, Z.-Q.; Zou, X. Morphological Traits, DELTA System, and Molecular Analysis for Gibellula Clavispora Sp. Nov. from China. Mycotaxon 2016, 131, 111–121. [Google Scholar] [CrossRef]
- Han, Y.-F.; Chen, W.H.; Zou, X.; Liang, Z.-Q. Gibellula curvispora, a New Species of Gibellula. Mycosystem 2013, 32, 777–780. [Google Scholar]
- Kuephadungphan, W.; Macabeo, A.P.G.; Luangsa-ard, J.J.; Tasanathai, K.; Thanakitpipattana, D.; Phongpaichit, S.; Yuyama, K.; Stadler, M. Studies on the Biologically Active Secondary Metabolites of the New Spider Parasitic Fungus Gibellula gamsii. Mycol. Prog. 2019, 18, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Chen, W.H.; Han, Y.-F.; Liang, Z.-Q. A New Species of the Genus Gibellula. Mycosystem 2016, 35, 1161–1168. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Centro de Coleções Taxonômicas O Centro de Biodiversidade Da UFMG. Available online: https://www2.icb.ufmg.br/cct/ (accessed on 10 January 2022).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Sun, B.L. Ancient and Recent Patterns of Geographic Speciation in the Oyster Mushroom Pleurotus Revealed by Phylogenetic Analysis of Ribosomal DNA Sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 4599–4603. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.-H.; Hyten, A.S.; Spatafora, J.W. Multigene Phylogeny Reveals New Lineage for Stachybotrys Chartarum, the Indoor Air Fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Murata, N.; Aoki, T.; Kusaba, M.; Tosa, Y.; Chuma, I. Various Species of Pyricularia Constitute a Robust Clade Distinct from Magnaporthe Salvinii and Its Relatives in Magnaporthaceae. J. Gen. Plant Pathol. 2014, 80, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geneious Prime Geneious Prime ® 2022.1.1 2022. Available online: https://www.geneious.com (accessed on 30 November 2022).
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.M.; Sondhi, S.; Ziesel, A.; Paliwal, S.; Fiumera, H.L. Mitochondrial-Nuclear Coadaptation Revealed through MtDNA Replacements in Saccharomyces cerevisiae. BMC Evol. Biol. 2020, 20, 128. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian Evolutionary Analysis by Sampling Trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Rambaut, A. FigTree: Tree Figure Drawing Tool Version 1.4. 3. 2009 2017. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 30 November 2022).
- Inkscape Inkscape: Draw Freely 2021. Available online: https://inkscape.org/pt-br/ (accessed on 30 November 2022).
- Google Earth Google Earth Pro 2022. Available online: https://www.google.com.br/earth/ (accessed on 30 November 2022).
- Montana State University Convert Geographic Units. Available online: http://rcn.montana.edu/Resources/Converter.aspx (accessed on 30 November 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Use R! Springer International Publishing: New York, NY, USA, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Samson, R.A.; Evans, H.C. Notes on Entomogenous Fungi from Ghana: I. The Genera Gibellula and Pseudogibellula. Acta Bot. Neerl. 1973, 22, 522–528. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Bałazy, S. Gibellula leiopus (Vuillemin in Maublanc) Mains—The Fungus Pathogenic for Spiders—Gibellula leiopus (Vuillemin in Maublanc) Mains. Acta Mycol. 2014, 6, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Humber, R.A.; Rombach, M.C. Torrubiella ratticaudata sp. nov. (Pyrenomycetes: Clavicipitales) and Other Fungi from Spiders on the Solomon Islands. Mycologia 1987, 79, 375–382. [Google Scholar] [CrossRef]
- Morgan, A.P. The Genus Gibellula Cavara. J. Mycol. 1905, 11, 49–50. [Google Scholar] [CrossRef]
- Hennings, P. Organ Für Kryptogamenkunde Und Phytopathologie Melist Repertorium Für Literature. Hedwigia 1902, 4, 148–149. [Google Scholar]
- von Höhnel, F. Eumycetes et Myxomycetes: Bearbeitet von Franz v. Höhnel. In Ergebnisse der Botanischen Expedition der Kaiserlichen Akademie der Wissenschaften nach Süd-Brasilien 1901. Denkschriften der Akademie der Wissenschaften.Math.Natw.Kl. 1907, 83, 1–45. [Google Scholar]
- Sawada, K. Descriptive Catalogue of Taiwan (Formosan) Fungi. Part IX. Spec. Publ Coll. Agric. Taiwan Univ 1959, 8, 228–229. [Google Scholar]
- Deighton, F.C. List of Fungi Collected in Sierra Leone. Bull. Misc. Inf. (R. Gard. Kew) 1936, 1936, 424. [Google Scholar] [CrossRef]
- Petch, T. Notes on Entomogenous Fungi. Trans. Br. Mycol. Soc. 1944, 27, 81–93. [Google Scholar] [CrossRef]
- Höfer, H.; Brescovit, A.D. Species and Guild Structure of a Neotropical Spider Assemblage (Araneae) from Reserva Ducke, Amazonas, Brazil. Andrias 2001, 15, 99–119. [Google Scholar]
- Nascimento, F.F.; dos Reis, M.; Yang, Z. A Biologist’s Guide to Bayesian Phylogenetic Analysis. Nat. Ecol. Evol. 2017, 1, 1446–1454. [Google Scholar] [CrossRef] [Green Version]
- Elias, S.G.; Salvador-Montoya, C.A.; Costa-Rezende, D.H.; Guterres, D.C.; Fernandes, M.; Olkoski, D.; Klabunde, G.H.F.; Drechsler-Santos, E.R. Studies on the Biogeography of Phellinotus piptadeniae (Hymenochaetales, Basidiomycota): Expanding the Knowledge on Its Distribution and Clarifying Hosts Relationships. Fungal. Ecol. 2020, 45, 100912. [Google Scholar] [CrossRef]
- Ferraz, K.M.P.B.; Oliveira, B.G.; Attias, N.; Desbiez, A.L.J. Species Distribution Model Reveals Only Highly Fragmented Suitable Patches Remaining for Giant Armadillo in the Brazilian Cerrado. Perspect. Ecol. Conserv. 2021, 19, 43–52. [Google Scholar] [CrossRef]
- Soberón, J. Grinnellian and Eltonian Niches and Geographic Distributions of Species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar] [CrossRef]
- Hao, T.; Guillera-Arroita, G.; May, T.W.; Lahoz-Monfort, J.J.; Elith, J. Using Species Distribution Models For Fungi. Fungal. Biol. Rev. 2020, 34, 74–88. [Google Scholar] [CrossRef]
- Wheeler, W.C.; Coddington, J.A.; Crowley, L.M.; Dimitrov, D.; Goloboff, P.A.; Griswold, C.E.; Hormiga, G.; Prendini, L.; Ramírez, M.J.; Sierwald, P.; et al. The Spider Tree of Life: Phylogeny of Araneae Based on Target-gene Analyses from an Extensive Taxon Sampling. Cladistics 2016, 33, 574–616. [Google Scholar] [CrossRef]
- van der Bijl, P.A. A Fungus—Gibellula haygarthii, Sp. n.— On a Spider of the Family Lycosidae. Trans. R. Soc. S. Afr. 1922, 10, 149–150. [Google Scholar] [CrossRef]
- Ruszkiewicz-Michalska, M.; Tkaczuk, C.; Dynowska, M.; Sucharzewska, E.; Szkodzik, J.; Wrzosek, M. Preliminary Studies of Fungi in the Biebrza National Park (NE Poland). I. Micromycetes. Acta Mycol. 2013, 47, 213–234. [Google Scholar] [CrossRef]
- Kepler, R.M.; Sung, G.; Harada, Y.; Tanaka, K.; Tanaka, E.; Hosoya, T.; Bischoff, J.F.; Spatafora, J.W. Host Jumping onto Close Relatives and across Kingdoms by Tyrannicordyceps (Clavicipitaceae) Gen. Nov. and Ustilaginoidea (Clavicipitaceae). Am. J. Bot. 2012, 99, 552–561. [Google Scholar] [CrossRef]
- Nikoh, N.; Fukatsu, T. Interkingdom Host Jumping Underground: Phylogenetic Analysis of Entomoparasitic Fungi of the Genus Cordyceps. Mol. Biol. Evol. 2000, 17, 629–638. [Google Scholar] [CrossRef] [PubMed]
- de Menezes, T.A.; Aburjaile, F.F.; Quintanilha-Peixoto, G.; Tomé, L.M.R.; Fonseca, P.L.C.; Mendes-Pereira, T.; Araújo, D.S.; Melo, T.S.; Kato, R.B.; Delabie, J.H.C.; et al. Unraveling the Secrets of a Double-Life Fungus by Genomics: Ophiocordyceps australis CCMB661 Displays Molecular Machinery for Both Parasitic and Endophytic Lifestyles. JoF 2023, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Loreto, R.G.; Elliot, S.L.; Freitas, M.L.R.; Pereira, T.M.; Hughes, D.P. Long-Term Disease Dynamics for a Specialized Parasite of Ant Societies: A Field Study. PLoS ONE 2014, 9, e103516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Małagocka, J.; Grell, M.N.; Lange, L.; Eilenberg, J.; Jensen, A.B. Transcriptome of an Entomophthoralean Fungus (Pandora formicae) Shows Molecular Machinery Adjusted for Successful Host Exploitation and Transmission. J. Invertebr. Pathol. 2015, 128, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Krasnoff, S.B.; Watson, D.W.; Gibson, D.M.; Kwan, E.C. Behavioral Effects of the Entomopathogenic Fungus, Entomophthora muscae on Its Host Musca Domestica: Postural Changes in Dying Hosts and Gated Pattern of Mortality. J. Insect Physiol. 1995, 41, 895–903. [Google Scholar] [CrossRef]
- Maitland, D.P. A Parasitic Fungus Infecting Yellow Dungflies Manipulates Host Perching Behaviour. Proc. R. Soc. Lond. B 1994, 258, 187–193. [Google Scholar] [CrossRef]
- Carruthers, R.I.; Ramos, M.E.; Larkin, T.S.; Hostetter, D.L.; Soper, R.S. The Entomophaga grylli (Fresenius) Batko Species Complex: Its Biology, Ecology, and Use For Biological Control of Pest Grasshoppers. Mem. Entomol. Soc. Can. 1997, 129, 329–353. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes-Pereira, T.; de Araújo, J.P.M.; Kloss, T.G.; Costa-Rezende, D.H.; de Carvalho, D.S.; Góes-Neto, A. Disentangling the Taxonomy, Systematics, and Life History of the Spider-Parasitic Fungus Gibellula (Cordycipitaceae, Hypocreales). J. Fungi 2023, 9, 457. https://doi.org/10.3390/jof9040457
Mendes-Pereira T, de Araújo JPM, Kloss TG, Costa-Rezende DH, de Carvalho DS, Góes-Neto A. Disentangling the Taxonomy, Systematics, and Life History of the Spider-Parasitic Fungus Gibellula (Cordycipitaceae, Hypocreales). Journal of Fungi. 2023; 9(4):457. https://doi.org/10.3390/jof9040457
Chicago/Turabian StyleMendes-Pereira, Thairine, João Paulo Machado de Araújo, Thiago Gechel Kloss, Diogo Henrique Costa-Rezende, Daniel Santana de Carvalho, and Aristóteles Góes-Neto. 2023. "Disentangling the Taxonomy, Systematics, and Life History of the Spider-Parasitic Fungus Gibellula (Cordycipitaceae, Hypocreales)" Journal of Fungi 9, no. 4: 457. https://doi.org/10.3390/jof9040457