
Penicillium citrinum Provides Transkingdom Growth Benefits in Choy Sum (Brassica rapa var. parachinensis)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungus Culture Conditions and Molecular Identification
2.2. Plant Growth-Promotion Assays
2.3. Agrobacterium-Mediated Transformation of B9
2.4. Fungal Interaction/Colonization Assays in Choy Sum Roots
2.5. Plant Growth Assay under Low Pi Using P. citrinum Isolate(s)
2.6. Extraction and Purification Methods for Detection of Phytohormones in P. citrinum
2.7. Liquid Chromatography–Mass Spectrometry
2.8. Statistical Analysis
3. Results
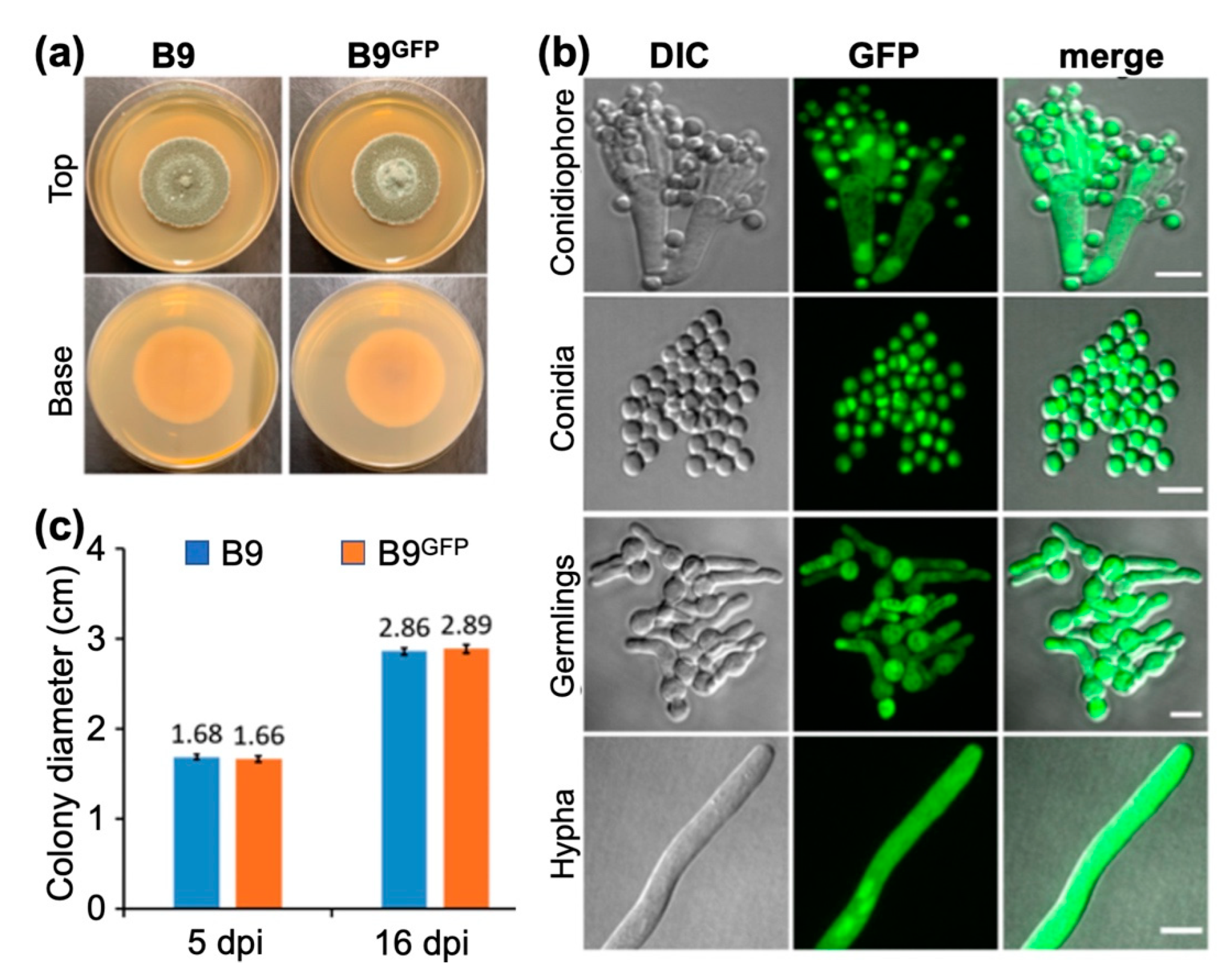
3.1. Morphological Characteristics and Identification of Fungal Isolates B9 and FLP7
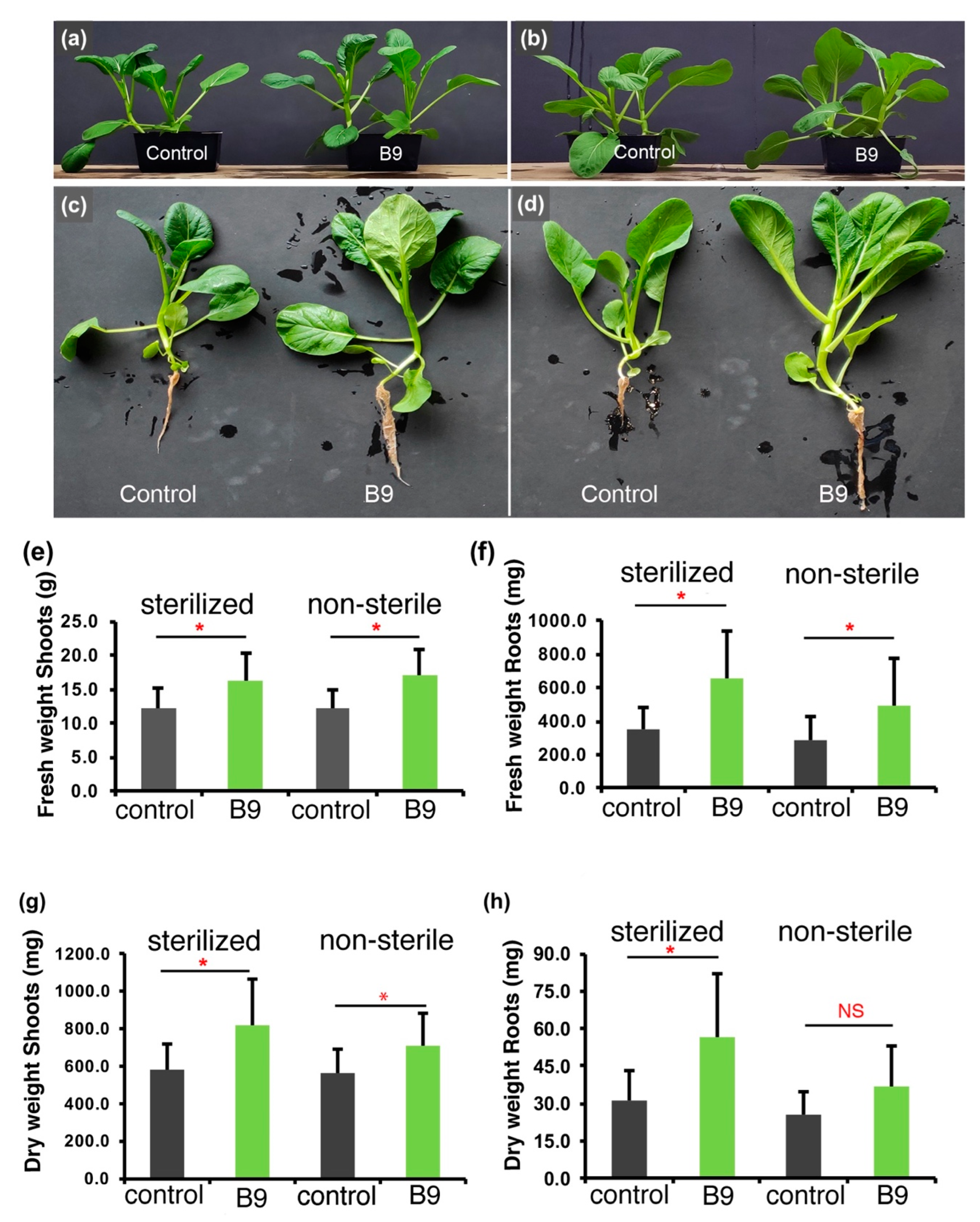
3.2. P. citrinum Improves Choy Sum Growth under Nutrient Rich Conditions
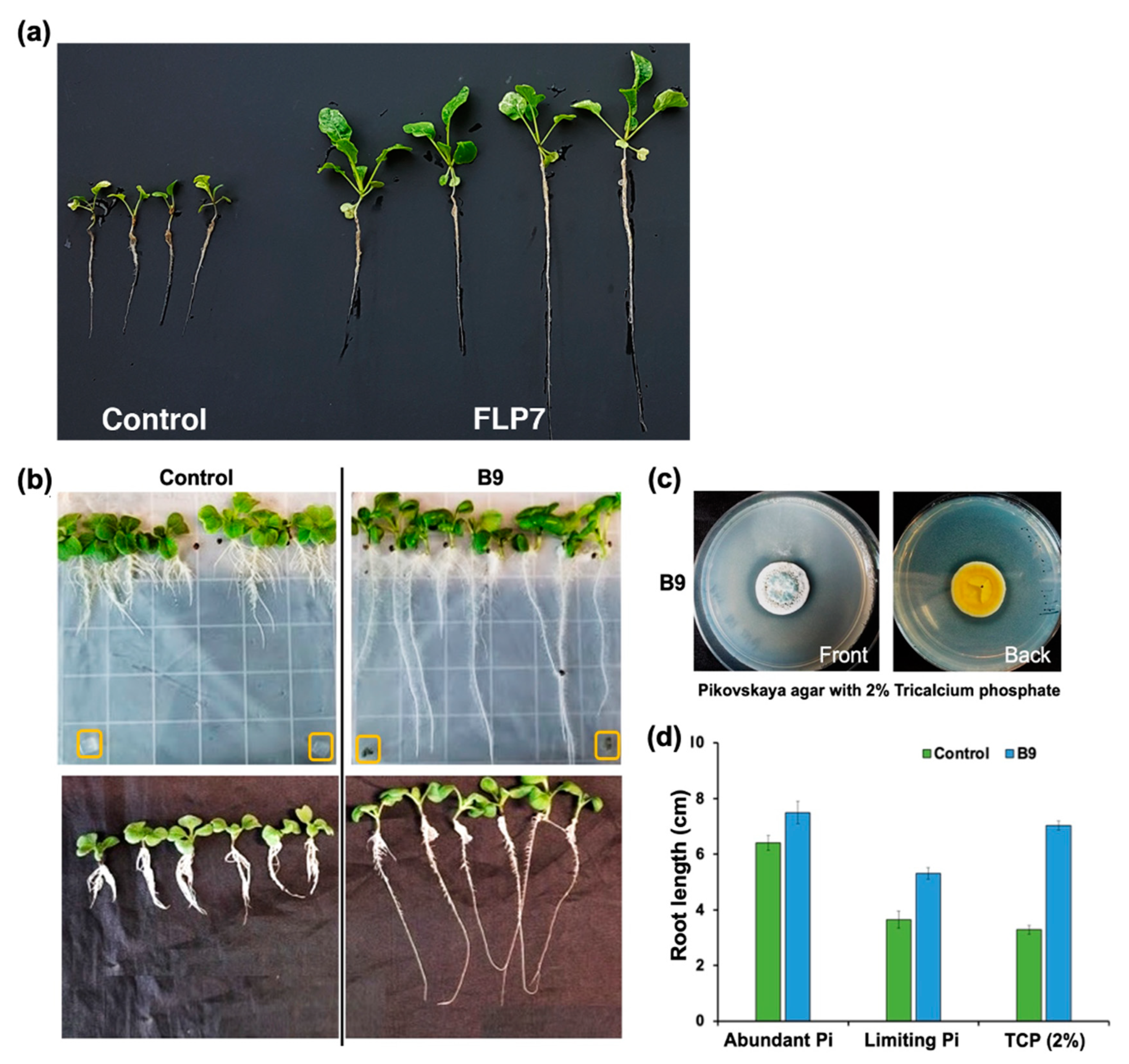
3.3. P. citrinum Improves Choy Sum Growth under Pi-Limiting Conditions
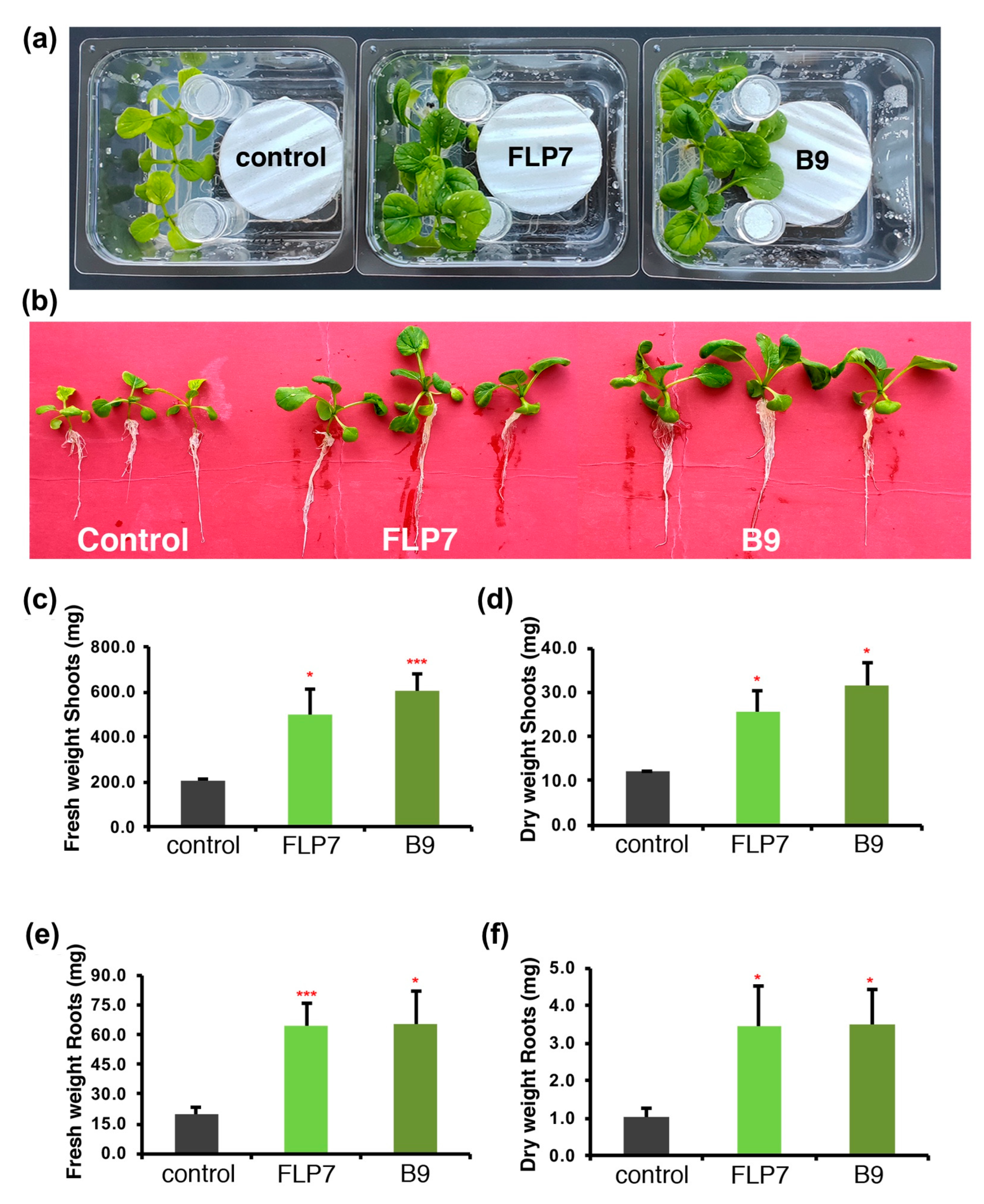
3.4. P. citrinum Isolates Enhance Choy Sum Growth via Volatile Secondary Metabolites
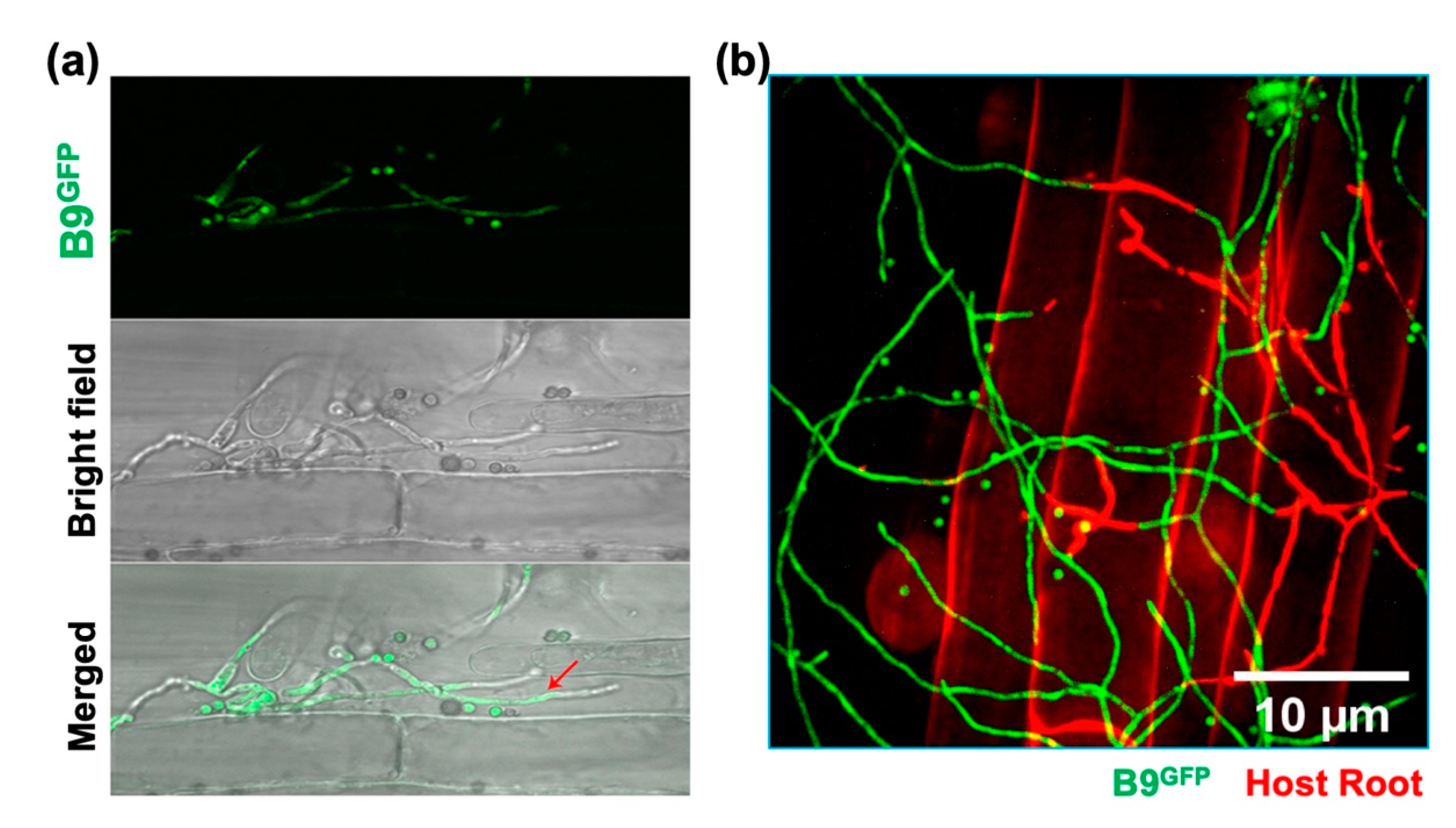
3.5. Analysing the Colonization of Plant Roots by P. citrinum Isolate B9
3.6. P. citrinum Produces the Phytohormones Gibberellin and Cytokinin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poveda, J.; Zabalgogeazcoa, I.; Soengas, P.; Rodriguez, V.M.; Cartea, M.E.; Abilleira, R.; Velasco, P. Brassica oleracea var. acephala (kale) improvement by biological activity of root endophytic fungi. Sci. Rep. 2020, 10, 20224. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.K.; Goenadie, V.; Lee, H.W.; Liang, X.; Loh, C.S.; Ong, C.N.; Tan, H.T.W. Growth and glucosinolate profiles of a common Asian green leafy vegetable, Brassica rapa subsp. chinensis var. parachinensis (choy sum), under LED lighting. Sci. Hortic. 2020, 261, 108922. [Google Scholar] [CrossRef]
- Gupta, S.K. Biotechnology of Crucifers, 1st ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Zou, L.; Tan, W.K.; Du, Y.; Lee, H.W.; Liang, X.; Lei, J.; Striegel, L.; Weber, N.; Rychlik, M.; Ong, C.N. Nutritional metabolites in Brassica rapa subsp. chinensis var. parachinensis (choy sum) at three different growth stages: Microgreen, seedling and adult plant. Food Chem. 2021, 357, 129535. [Google Scholar] [CrossRef]
- Huang, J.J.; D’Souza, C.; Zhou, W. Light-Time-Biomass Response Model for Predicting the Growth of Choy Sum (Brassica rapa var. parachinensis) in Soil-Based LED-Constructed Indoor Plant Factory for Efficient Seedling Production. Front. Plant Sci. 2021, 12, 623682. [Google Scholar] [CrossRef]
- Poveda, J.; Díaz-González, S.; Díaz-Urbano, M.; Velasco, P.; Sacristán, S. Fungal endophytes of Brassicaceae: Molecular interactions and crop benefits. Front. Plant Sci. 2022, 13, 932288. [Google Scholar] [CrossRef]
- Poveda, J.; Rodríguez, V.M.; Díaz-Urbano, M.; Sklenář, F.; Saati-Santamaría, Z.; Menéndez, E.; Velasco, P. Endophytic fungi from kale (Brassica oleracea var. acephala) modify roots-glucosinolate profile and promote plant growth in cultivated Brassica species. First description of Pyrenophora gallaeciana. Front. Microbiol. 2022, 13, 981507. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, K.; Kobae, Y.; Toju, H. Beneficial associations between Brassicaceae plants and fungal endophytes under nutrient-limiting conditions: Evolutionary origins and host-symbiont molecular mechanisms. Curr. Opin. Plant Biol. 2018, 44, 145–154. [Google Scholar] [CrossRef]
- Jiang, L.; Seo, J.; Peng, Y.; Jeon, D.; Park, S.J.; Kim, C.Y.; Kim, P.I.; Kim, C.H.; Lee, J.H.; Lee, J. Genome insights into the plant growth-promoting bacterium Saccharibacillus brassicae ATSA2(T). AMB Express 2023, 13, 9. [Google Scholar] [CrossRef]
- Jia, R.; Chen, J.; Hu, L.; Liu, X.; Xiao, K.; Wang, Y. Alcaligenes faecalis Juj3 alleviates Plasmodiophora brassicae stress to cabbage via promoting growth and inducing resistance. Front. Sustain. Food Syst. 2022, 6, 942409. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moenne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dye, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Mabood, F.; Zhou, X.; Smith, D.L. Microbial signaling and plant growth promotion. Can. J. Plant Sci. 2014, 94, 1051–1063. [Google Scholar] [CrossRef]
- Hyakumachi, M. Plant-Growth-Promoting Fungi from Turfgrass Rhizosphere with Potential for Disease Suppression. Soil Microorg. 1994, 44, 53–68. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Miyazawa, M.; Hyakumachi, M. Plant growth-promoting fungus Penicillium spp. GP15-1 enhances growth and confers protection against damping-off and anthracnose in the cucumber. J. Oleo Sci. 2014, 63, 391–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): Phytostimulation and Induced Systemic Resistance. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: Singapore, 2017; pp. 135–191. [Google Scholar]
- Murali, M.; Naziya, B.; Ansari, M.A.; Alomary, M.N.; AlYahya, S.; Almatroudi, A.; Thriveni, M.C.; Gowtham, H.G.; Singh, S.B.; Aiyaz, M.; et al. Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture. J. Fungi 2021, 7, 314. [Google Scholar] [CrossRef]
- Yin, Z.; Shi, F.; Jiang, H.; Roberts, D.P.; Chen, S.; Fan, B. Phosphate solubilization and promotion of maize growth by Penicillium oxalicum P4 and Aspergillus niger P85 in a calcareous soil. Can. J. Microbiol. 2015, 61, 913–923. [Google Scholar] [CrossRef]
- Choi, W.Y.; Rim, S.O.; Lee, J.H.; Lee, J.M.; Lee, I.J.; Cho, K.J.; Rhee, I.K.; Kwon, J.B.; Kim, J.G. Isolation of Gibberellins-Producing Fungi from the Root of Several Sesamum indicum Plants. J. Microbiol. Biotechnol. 2005, 15, 22–28. [Google Scholar]
- Babu, A.G.; Kim, S.W.; Yadav, D.R.; Hyum, U.; Adhikari, M.; Lee, Y.S. Penicillium menonorum: A Novel Fungus to Promote Growth and Nutrient Management in Cucumber Plants. Mycobiology 2015, 43, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Dovana, F.; Mucciarelli, M.; Mascarello, M.; Fusconi, A. In Vitro Morphogenesis of Arabidopsis to Search for Novel Endophytic Fungi Modulating Plant Growth. PLoS ONE 2015, 10, e0143353. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Lee, I.J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Luan, Y.; An, L.; Yu, K. Arbuscular mycorrhizae formed by Penicillium pinophilum improve the growth, nutrient uptake and photosynthesis of strawberry with two inoculum-types. Biotechnol. Lett. 2008, 30, 1489–1494. [Google Scholar] [CrossRef] [PubMed]
- Khalmuratova, I.; Kim, H.; Nam, Y.J.; Oh, Y.; Jeong, M.J.; Choi, H.R.; You, Y.H.; Choo, Y.S.; Lee, I.J.; Shin, J.H.; et al. Diversity and Plant Growth Promoting Capacity of Endophytic Fungi Associated with Halophytic Plants from the West Coast of Korea. Mycobiology 2015, 43, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, R.; Kang, S.-M.; Baek, I.-Y.; Lee, I.-J. Characterization of plant growth-promoting traits of Penicillium species against the effects of high soil salinity and root disease. J. Plant Interact. 2014, 9, 754–762. [Google Scholar] [CrossRef]
- Selvaraj, P.; Shen, Q.; Yang, F.; Naqvi, N.I. Cpk2, a Catalytic Subunit of Cyclic AMP-PKA, Regulates Growth and Pathogenesis in Rice Blast. Front. Microbiol. 2017, 8, 2289. [Google Scholar] [CrossRef] [Green Version]
- Soundararajan, S.; Jedd, G.; Li, X.; Ramos-Pamploña, M.; Chua, N.H.; Naqvi, N.I. Woronin body function in Magnaporthe grisea is essential for efficient pathogenesis and for survival during nitrogen starvation stress. Plant Cell 2004, 16, 1564–1574. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Zhou, J.; He, Y.; Xie, Q.; Chen, A.; Zheng, H.; Shi, L.; Zhao, X.; Zhang, C.; Huang, Q.; et al. Retromer Is Essential for Autophagy-Dependent Plant Infection by the Rice Blast Fungus. PLoS Genet. 2015, 11, e1005704. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Jung, H.Y.; Lee, J.H.; Lee, I.J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Morrison, E.N.; Knowles, S.; Hayward, A.; Thorn, R.G.; Saville, B.J.; Emery, R.J.N. Detection of phytohormones in temperate forest fungi predicts consistent abscisic acid production and a common pathway for cytokinin biosynthesis. Mycologia 2015, 107, 245–257. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Umashankar, S.; Liang, X.; Lee, H.W.; Swarup, S.; Ong, C.N. Characterization of Plant Volatiles Reveals Distinct Metabolic Profiles and Pathways among 12 Brassicaceae Vegetables. Metabolites 2018, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Hiruma, K.; Gerlach, N.; Sacristan, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramirez, D.; Bucher, M.; O’Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, R.; Lee, S.; Bennett, J.W. Arabidopsis thaliana as a model system for testing the effect of Trichoderma volatile organic compounds. Fungal Ecol. 2013, 6, 19–26. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kimura, M.; Miyazawa, M.; Hyakumachi, M. Analysis of volatile organic compounds emitted by plant growth-promoting fungus Phoma sp. GS8-3 for growth promotion effects on tobacco. Microbes Environ. 2013, 28, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, F.; Zafari, D.; Salari, H. Volatile organic compounds of some Trichoderma spp. increase growth and induce salt tolerance in Arabidopsis thaliana. Fungal Ecol. 2017, 29, 67–75. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Sopher, C.R.; Vettakkorumakankav, N.N. Modulation of gibberellins protects plants from environmental stresses. Indian J. Plant Physiol. 2000, 5, 115–126. [Google Scholar]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- Schwechheimer, C. Gibberellin signaling in plants—The extended version. Front. Plant Sci. 2011, 2, 107. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Chakrabarty, S.K. Gibberellic acid in plant: Still a mystery unresolved. Plant Signal. Behav. 2013, 8, e25504. [Google Scholar] [CrossRef] [Green Version]
- Hasan, H.A.H. Gibberellin and auxin production by plant root-fungi and their biosynthesis under salinity-calcium interaction. Plant Soil Environ. 2002, 48, 101–106. [Google Scholar] [CrossRef]
- Nassar, A.H.; El-Tarabily, K.A.; Sivasithamparam, K. Promotion of plant growth by an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (Zea mays L.) roots. Biol. Fertil. Soils 2005, 42, 97–108. [Google Scholar] [CrossRef]
- Rim, S.O.; Lee, J.H.; Choi, W.Y.; Hwang, S.K.; Suh, S.J.; Lee, I.J.; Rhee, I.K.; Kim, J.G. Fusarium proliferatum KGL0401 as a New Gibberellin-Producing Fungus. J. Microbiol. Biotechnol. 2005, 15, 809–814. [Google Scholar]
- Claeys, H.; De Bodt, S.; Inze, D. Gibberellins and DELLAs: Central nodes in growth regulatory networks. Trends Plant Sci. 2014, 19, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, A.; Roldan, A.; Albacete, A.; Pascual, J.A. The interaction with arbuscular mycorrhizal fungi or Trichoderma harzianum alters the shoot hormonal profile in melon plants. Phytochemistry 2011, 72, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Ritter, C.; Venus, Y.; Camehl, I.; Varma, A.; Shahollari, B.; Novak, O.; Strnad, M.; Ludwig-Muller, J.; Oelmuller, R. The role of auxins and cytokinins in the mutualistic interaction between Arabidopsis and Piriformospora indica. Mol. Plant Microbe Interact. 2008, 21, 1371–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, M.; Piechulla, B. Plant growth promotion due to rhizobacterial volatiles--an effect of CO2? FEBS Lett. 2009, 583, 3473–3477. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Faubert, P.; Hagen, M.; Zu Castell, W.; Polle, A.; Schnitzler, J.P.; Rosenkranz, M. Volatile profiles of fungi—Chemotyping of species and ecological functions. Fungal Genet. Biol. 2013, 54, 25–33. [Google Scholar] [CrossRef]
- Hiltpold, I.; Turlings, T.C. Belowground chemical signaling in maize: When simplicity rhymes with efficiency. J. Chem. Ecol. 2008, 34, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Ditengou, F.A.; Muller, A.; Rosenkranz, M.; Felten, J.; Lasok, H.; van Doorn, M.M.; Legue, V.; Palme, K.; Schnitzler, J.P.; Polle, A. Volatile signalling by sesquiterpenes from ectomycorrhizal fungi reprogrammes root architecture. Nat. Commun. 2015, 6, 6279. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K. Plant methyl salicylate induces defense responses in the rhizobacterium Bacillus subtilis. Environ. Microbiol. 2015, 17, 1365–1376. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol.—Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Hung, R.; Yap, M.; Bennett, J.W. Age matters: The effects of volatile organic compounds emitted by Trichoderma atroviride on plant growth. Arch. Microbiol. 2015, 197, 723–727. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, K.; Chen, C.-Y.; Selvaraj, P.; Pavagadhi, S.; Yeap, Y.T.; Swarup, S.; Zheng, W.; Naqvi, N.I. Penicillium citrinum Provides Transkingdom Growth Benefits in Choy Sum (Brassica rapa var. parachinensis). J. Fungi 2023, 9, 420. https://doi.org/10.3390/jof9040420
Gu K, Chen C-Y, Selvaraj P, Pavagadhi S, Yeap YT, Swarup S, Zheng W, Naqvi NI. Penicillium citrinum Provides Transkingdom Growth Benefits in Choy Sum (Brassica rapa var. parachinensis). Journal of Fungi. 2023; 9(4):420. https://doi.org/10.3390/jof9040420
Chicago/Turabian StyleGu, Keyu, Cheng-Yen Chen, Poonguzhali Selvaraj, Shruti Pavagadhi, Yoon Ting Yeap, Sanjay Swarup, Wenhui Zheng, and Naweed I. Naqvi. 2023. "Penicillium citrinum Provides Transkingdom Growth Benefits in Choy Sum (Brassica rapa var. parachinensis)" Journal of Fungi 9, no. 4: 420. https://doi.org/10.3390/jof9040420