
Respiration, Production, and Growth Efficiency of Marine Pelagic Fungal Isolates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Fungal Cultures
2.2. Quantification of Fungal Biomass
2.3. Fungal Abundance
2.4. Fungal Respiration
2.5. Calculation of Fungal Growth Efficiency and Q10
2.6. Data Analyses and Statistics
3. Results
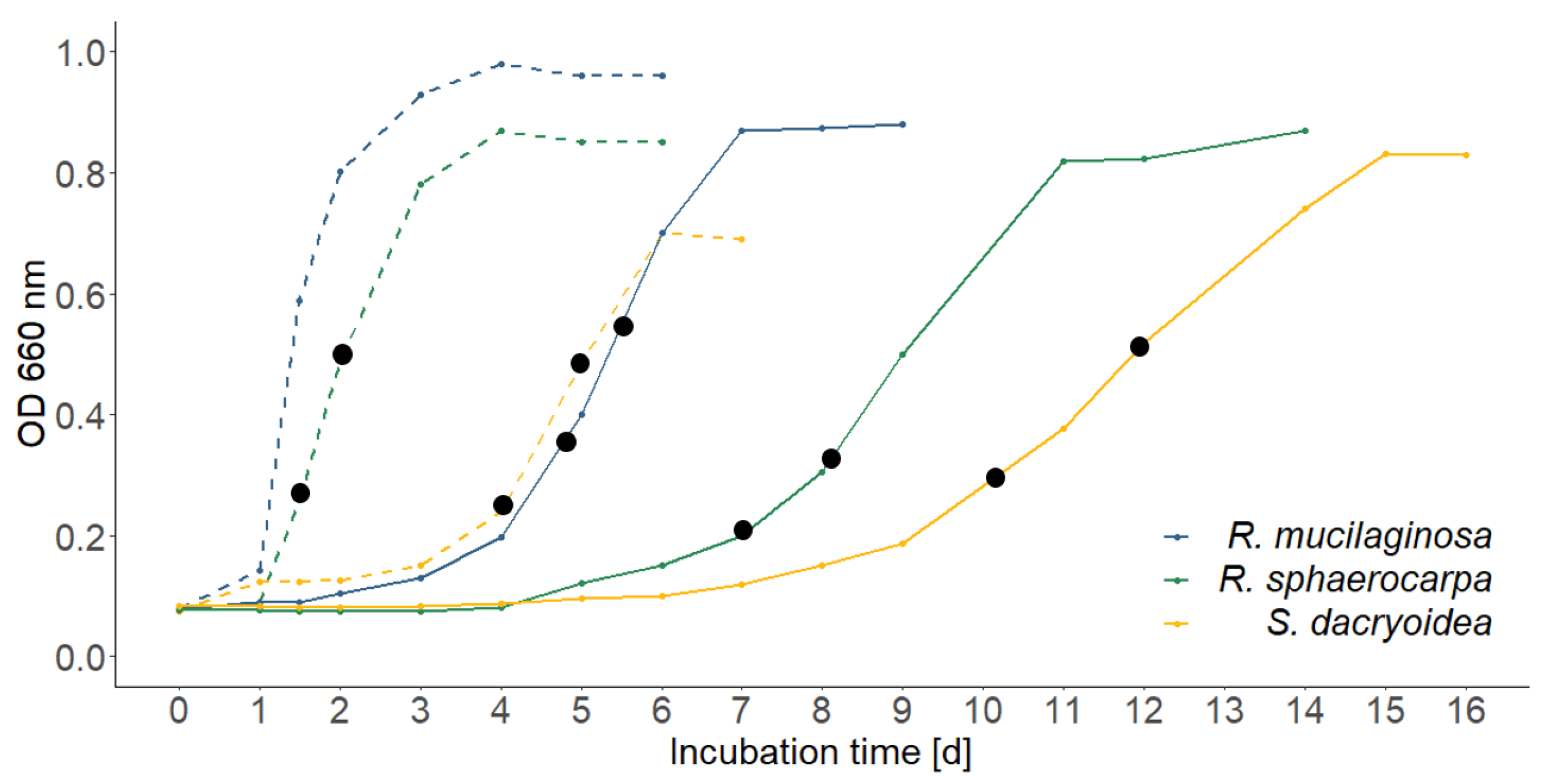
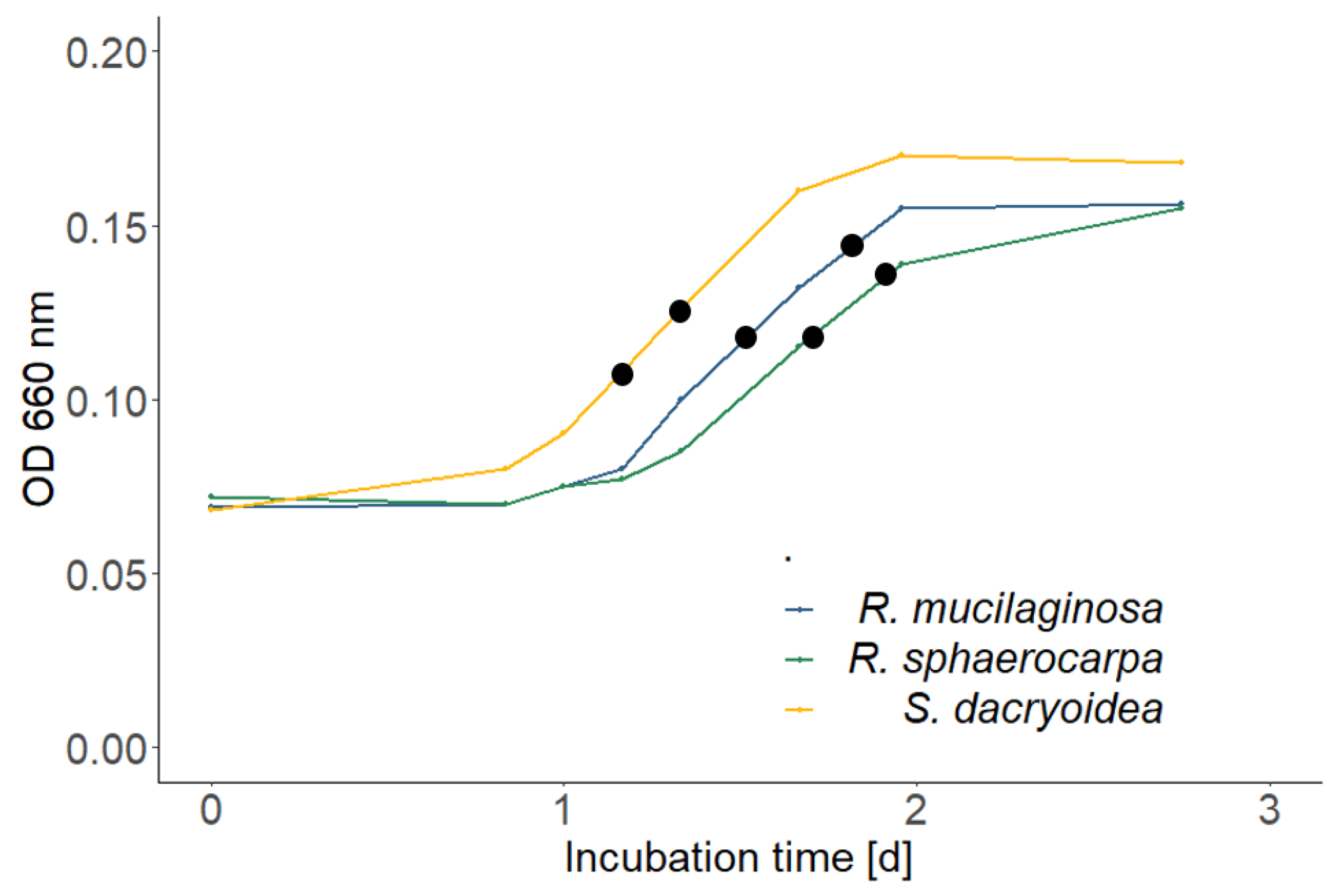
3.1. Growth Dynamics of the Fungal Cultures
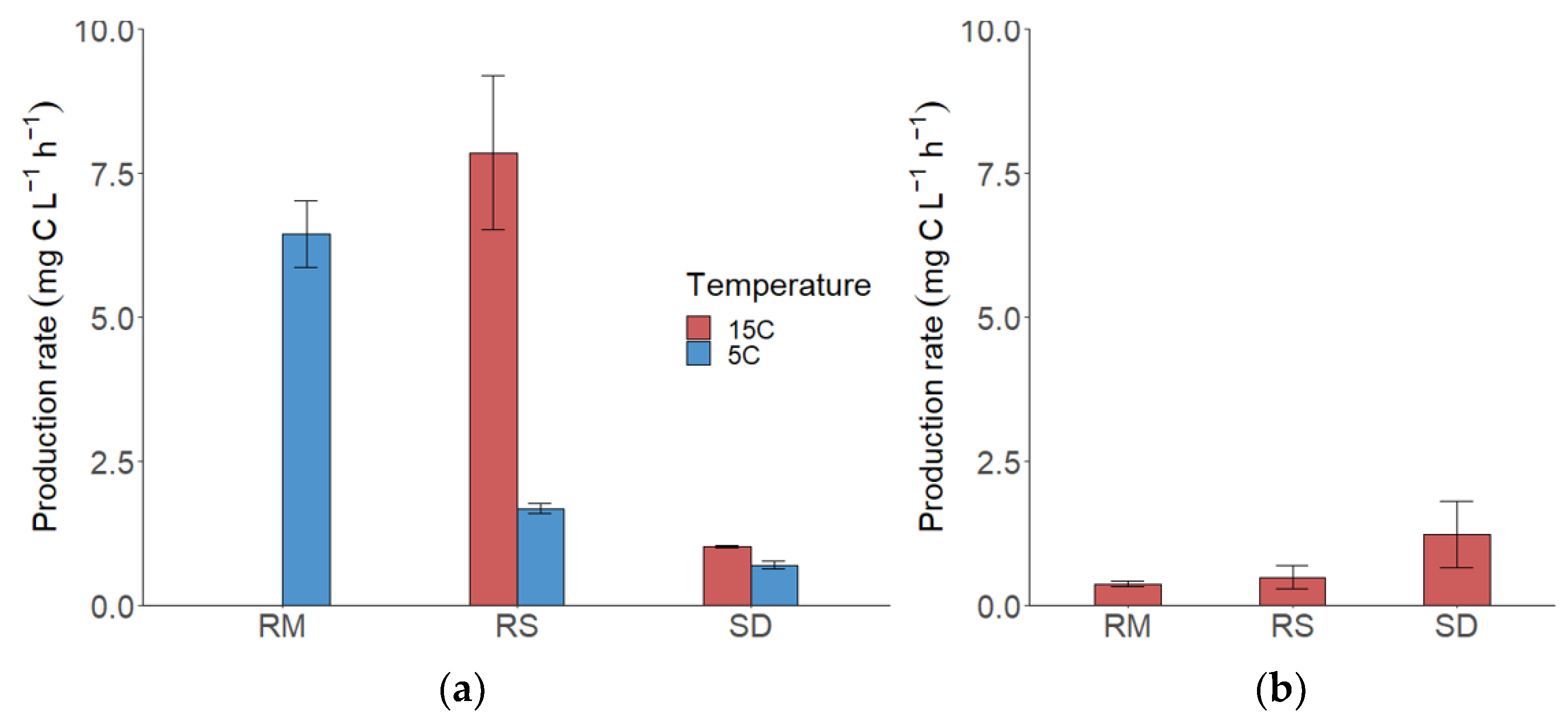
3.2. Fungal Production (FP)
3.3. Fungal Abundance
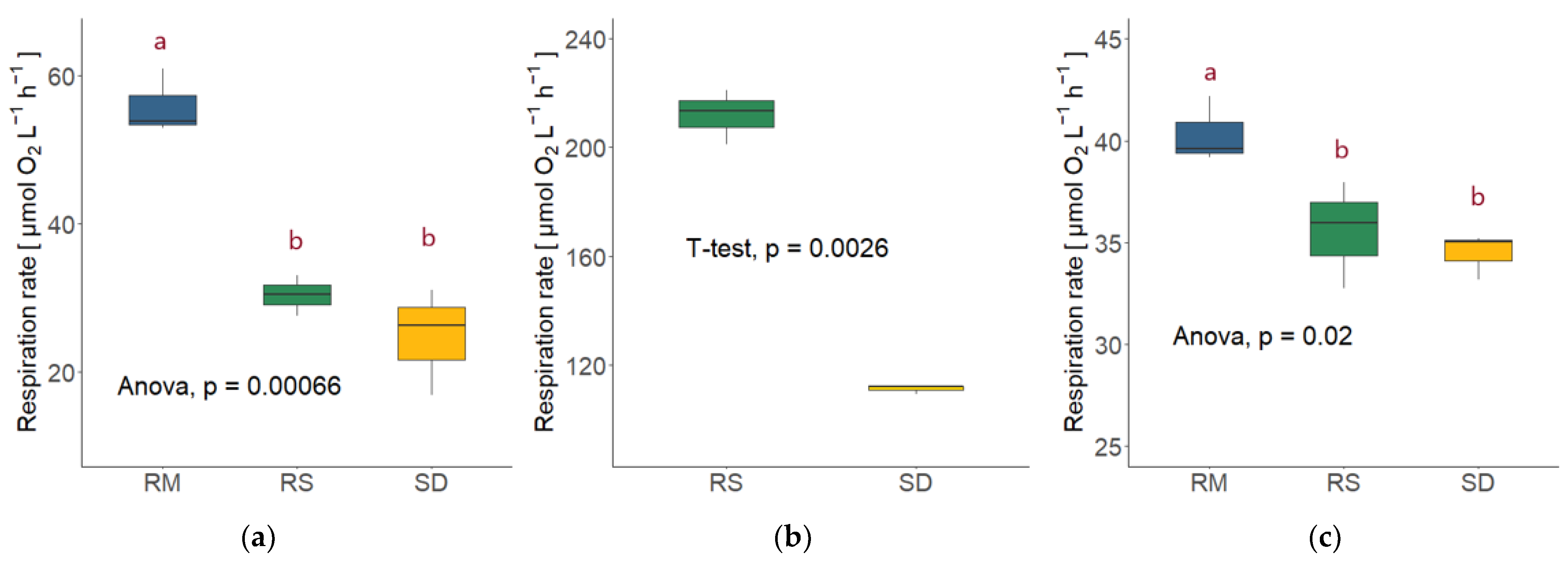
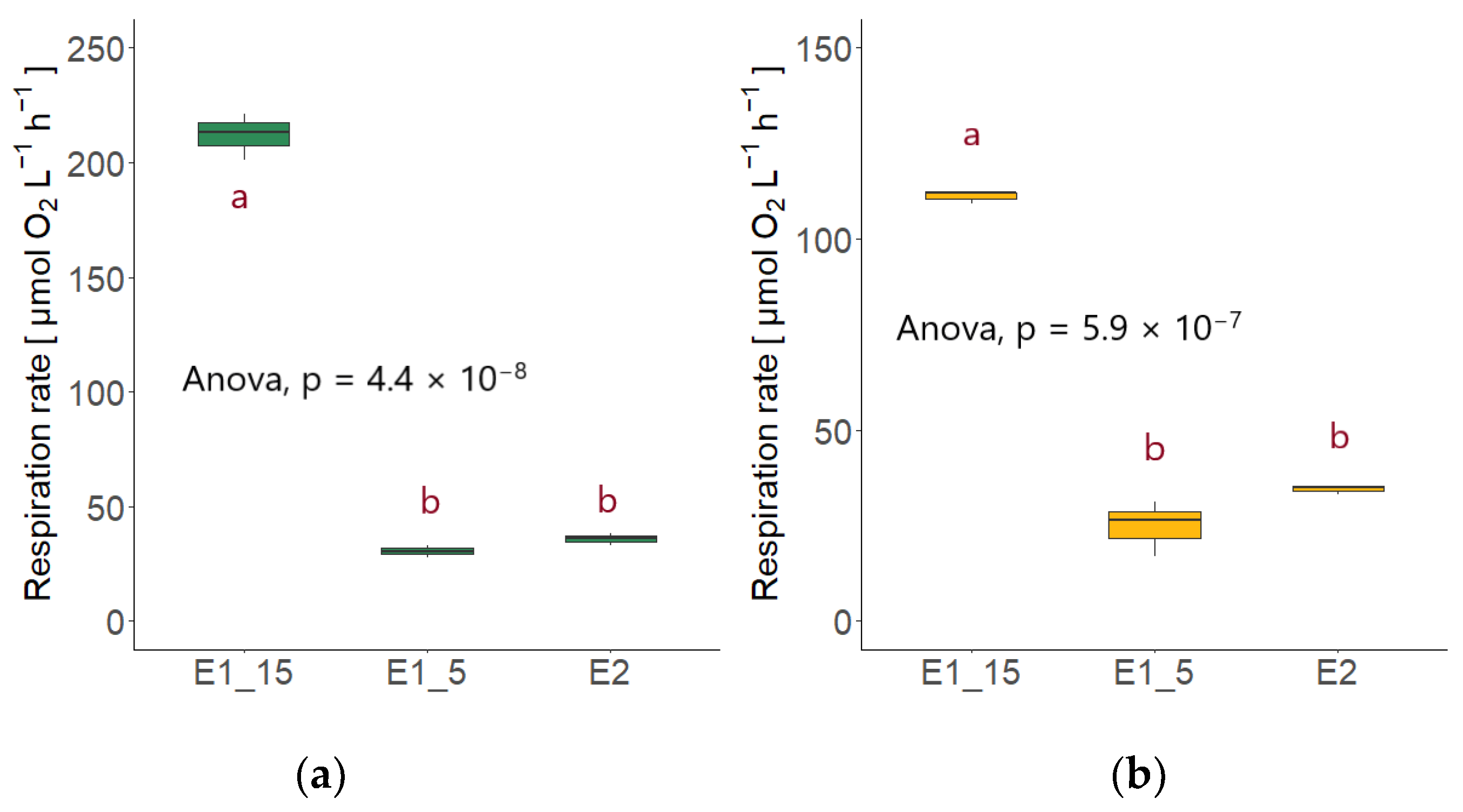
3.4. Fungal Respiration (FR)
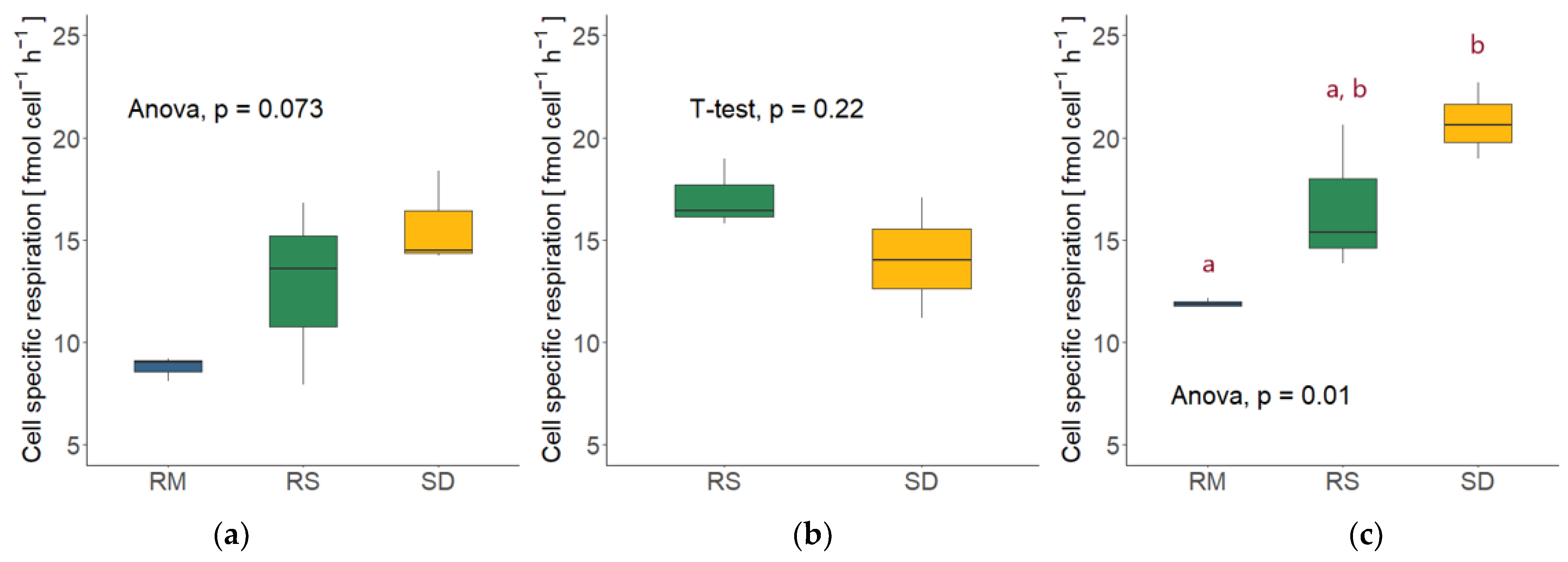
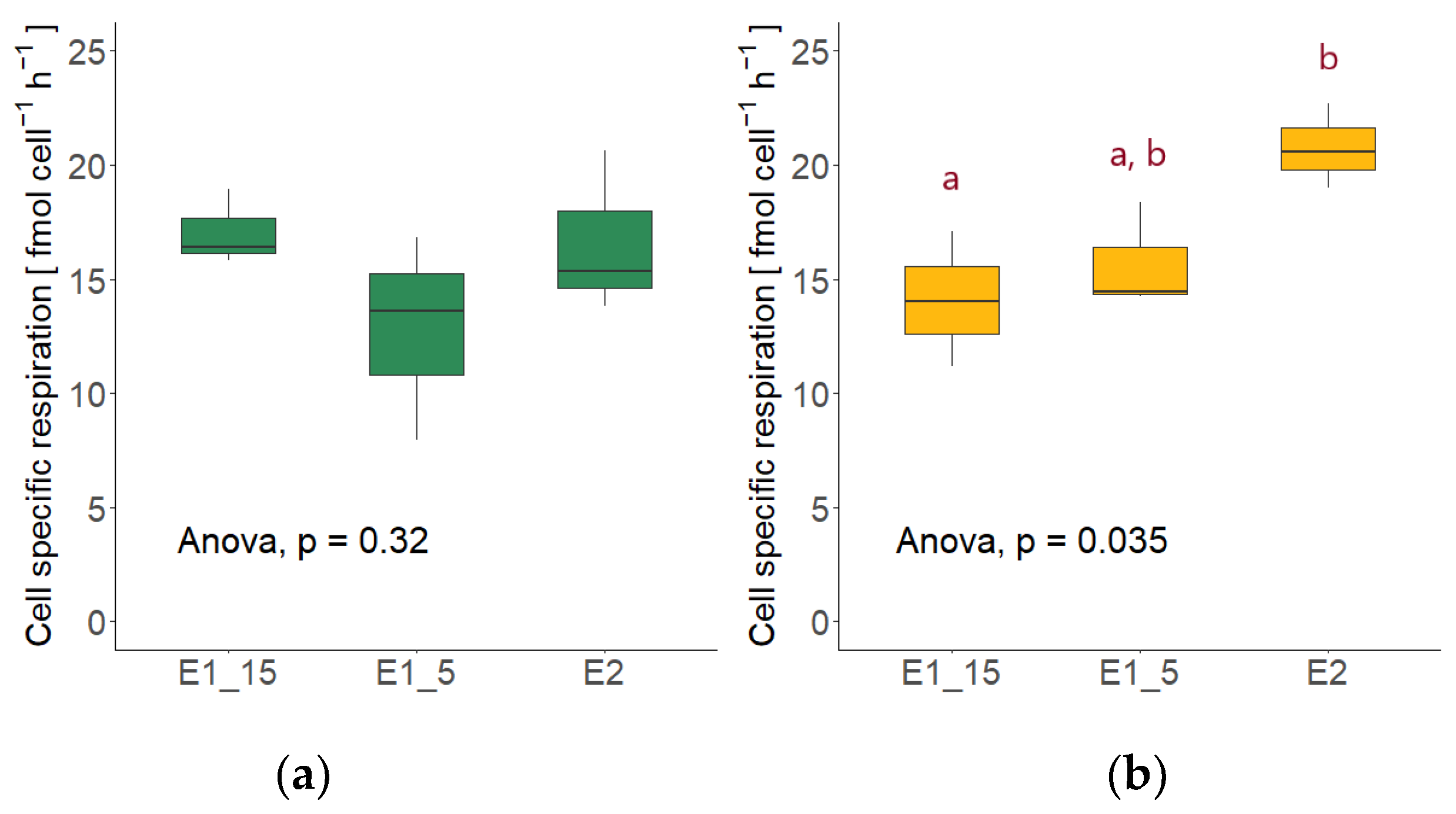
3.5. Cell-Specific Respiration
3.6. Fungal Growth Efficiency (FGE)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Worden, A.Z.; Follows, M.J.; Giovannoni, S.J.; Wilken, S.; Zimmerman, A.E.; Keeling, P.J. Rethinking the marine carbon cycle: Factoring in the multifarious lifestyles of microbes. Science 2015, 347, 1257594. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Legendre, L. Biogenic carbon cycling in the upper ocean: Effects of microbial respiration. Science 2001, 291, 2398–2400. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.R.; Ducklow, H.W. Microbial loop carbon cycling in ocean environments studied using a simple steady-state model. Aquat. Microb. Ecol. 2001, 26, 37–49. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.; Gray, J.; Meyer-Reil, L.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Del Giorgio, P.A.; Cole, J.J. Bacterial growth efficiency in natural aquatic systems. Annu. Rev. Ecol. Syst. 1998, 29, 503–541. [Google Scholar] [CrossRef]
- Robinson, C. Microbial respiration, the engine of ocean deoxygenation. Front. Mar. Sci. 2019, 5, 533. [Google Scholar] [CrossRef]
- Grossart, H.P.; Rojas-Jimenez, K. Aquatic fungi: Targeting the forgotten in microbial ecology. Curr. Opin. Microbiol. 2016, 31, 140–145. [Google Scholar] [CrossRef]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. MBio 2019, 10, e01189-18. [Google Scholar] [CrossRef]
- Bochdansky, A.B.; Clouse, M.A.; Herndl, G.J. Eukaryotic microbes, principally fungi and labyrinthulomycetes, dominate biomass on bathypelagic marine snow. ISME J. 2017, 11, 362–373. [Google Scholar] [CrossRef]
- Morales, S.E.; Biswas, A.; Herndl, G.J.; Baltar, F. Global structuring of phylogenetic and functional diversity of pelagic fungi by depth and temperature. Front. Mar. Sci. 2019, 6, 131. [Google Scholar] [CrossRef]
- Gao, Z.; Johnson, Z.I.; Wang, G. Molecular characterization of the spatial diversity and novel lineages of mycoplankton in Hawaiian coastal waters. ISME J. 2010, 4, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Yurkov, A.M.; Göker, M.; Lumbsch, H.T.; Leavitt, S.D.; Groenewald, M.; Theelen, B.; Liu, X.Z.; Boekhout, T.; Bai, F.Y. Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 2015, 81, 149–189. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.H.; Jara, A.M.; Pantoja, S. Fungal parasites infect marine diatoms in the upwelling ecosystem of the Humboldt current system off central Chile. Environ. Microbiol. 2016, 18, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, M.; Hollingsworth, A.; Bain, C.; Sharma, V.; Taylor, J.D. Algal polysaccharide utilisation by saprotrophic planktonic marine fungi. Fungal Ecol. 2017, 30, 135–138. [Google Scholar] [CrossRef]
- Priest, T.; Fuchs, B.; Amann, R.; Reich, M. Diversity and biomass dynamics of unicellular marine fungi during a spring phytoplankton bloom. Environ. Microbiol. 2021, 23, 448–463. [Google Scholar] [CrossRef]
- Baltar, F.; Zhao, Z.; Herndl, G.J. Potential and expression of carbohydrate utilization by marine fungi in the global ocean. Microbiome 2021, 9, 106. [Google Scholar] [CrossRef]
- Breyer, E.; Zhao, Z.; Herndl, G.J.; Baltar, F. Global contribution of pelagic fungi to protein degradation in the ocean. Microbiome 2022, 10, 143. [Google Scholar] [CrossRef]
- Chrismas, N.; Cunliffe, M. Depth-dependent mycoplankton glycoside hydrolase gene activity in the open ocean—Evidence from the Tara Oceans eukaryote metatranscriptomes. ISME J. 2020, 14, 2361–2365. [Google Scholar] [CrossRef]
- Orsi, W.; Vuillemin, A.; Coskun, Ö.K.; Rodriguez, P.; Oertel, Y.; Niggemann, J.; Mohrholz, V.; Gomez-Saez, G.V. Carbon assimilating fungi from surface ocean to subseafloor revealed by coupled phylogenetic and stable isotope analysis. ISME J. 2022, 16, 1245–1261. [Google Scholar] [CrossRef]
- Grossart, H.P.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 17, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.E.; Quiñones, R.A.; Gutiérrez, M.H.; Pantoja, S. Effects of temperature and glucose concentration on the growth and respiration of fungal species isolated from a highly productive coastal upwelling ecosystem. Fungal Ecol. 2015, 13, 135–149. [Google Scholar] [CrossRef]
- Wohlers, J.; Engel, A.; Zöllner, E.; Breithaupt, P.; Jürgens, K.; Hoppe, H.G.; Sommer, U.; Riebesell, U. Changes in biogenic carbon flow in response to sea surface warming. Proc. Natl. Acad. Sci. USA 2009, 106, 7067–7072. [Google Scholar] [CrossRef] [PubMed]
- Arnosti, C.; Bell, C.; Moorhead, D.L.; Sinsabaugh, R.L.; Steen, A.D.; Stromberger, M.; Wallenstein, M.; Weintraub, M.N. Extracellular enzymes in terrestrial, freshwater, and marine environments: Perspectives on system variability and common research needs. Biogeochemistry 2014, 117, 5–21. [Google Scholar] [CrossRef]
- UNESCO. Ocean Flux Study (JGOFS) Protocols for the Joint Global Core Measurements; IOC Manuals and Guides, 29; UNESCO: Paris, France, 1994. [Google Scholar]
- Precision Sensing GmbH. Oxygen Sensor Spots PSt3/PSt6; PreSens Precision Sensing GmbH: Regensburg, Germany, 2018. [Google Scholar]
- Warkentin, M.; Freese, H.M.; Karsten, U.; Schumann, R. New and fast method to quantify respiration rates of bacterial and plankton communities in freshwater ecosystems by using optical oxygen sensor spots. Appl. Environ. Microbiol. 2007, 73, 6722–6729. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2022. Available online: https://www.R-project.org/ (accessed on 23 March 2023).
- RStudio Team. RStudio: Integrated Development Environment for R. RStudio, PBC, Boston, MA. 2022. Available online: http://www.rstudio.com/ (accessed on 23 March 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Auguie, B. _gridExtra: Miscellaneous Functions for "Grid" Graphics. R package version 2.3. 2017. Available online: https://CRAN.R-project.org/package=gridExtra (accessed on 23 March 2023).
- Kassambra, A. _ggpubr: ‘ggplot2’ Based Publication Ready Plots_. R package version 0.5.0. 2022. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 23 March 2023).
- Motegi, C.; Nagata, T.; Miki, T.; Weinbauer, M.G.; Legendre, L.; Rassoulzadegand, F. Viral control of bacterial growth efficiency in marine pelagic environments. Limnol. Oceanogr. 2009, 54, 1901–1910. [Google Scholar] [CrossRef]
- Teira, E.; Fernández, A.; Álvarez-Salgado, X.A.; García-Martín, E.E.; Serret, P.; Sobrino, C. Response of two marine bacterial isolates to high CO2 concentration. Mar. Ecol. Prog. Ser. 2012, 453, 27–36. [Google Scholar] [CrossRef]
- White, P.A.; Kalff, J.; Rasmussen, J.B.; Gasol, J.M. The effect of temperature and algal biomass on bacterial production and specific growth rate in freshwater and marine habitats. Microb. Ecol. 1991, 21, 99–118. [Google Scholar] [CrossRef]
- Choi, J.W.; Peters, F. Effects of temperature on two psychrophilic ecotypes of a heterotrophic nanoflagellate, Paraphysomonas imperforata. Appl. Environ. Microbiol. 1992, 58, 593–599. [Google Scholar] [CrossRef]
- Del Giorgio, P.A.; Condon, R.; Bouvier, T.; Longnecker, K.; Bouvier, C.; Sherr, E.; Gasol, J.M. Coherent patterns in bacterial growth, growth efficiency, and leucine metabolism along a northeastern Pacific inshore-offshore transect. Limnol. Oceanogr. 2011, 56, 1–16. [Google Scholar] [CrossRef]
- Baltar, F.; Arístegui, J.; Gasol, J.M.; Sintes, E.; Herndl, G.J. Evidence of prokaryotic metabolism on suspended particulate organic matter in the dark waters of the subtropical North Atlantic. Limnol. Oceanogr. 2009, 54, 182–193. [Google Scholar] [CrossRef]
- Reinthaler, T.; Van Aken, H.; Veth, C.; Arístegui, J.; Robinson, C.; Williams, P.J.; Le, B.; Lebaron, P.; Herndl, G.J. Prokaryotic respiration and production in the meso- and bathypelagic realm of the eastern and western North Atlantic basin. Limnol. Oceanogr. 2006, 51, 1262–1273. [Google Scholar] [CrossRef]
- Mazuecos, I.P.; Arístegui, J.; Vázquez-Domínguez, E.; Ortega-Retuerta, E.; Gasol, J.M.; Reche, I. Temperature control of microbial respiration and growth efficiency in the mesopelagic zone of the South Atlantic and Indian Oceans. Deep. Res. Part I Oceanogr. Res. Pap. 2015, 95, 131–138. [Google Scholar] [CrossRef]
- Ikeda, T.; Kanno, Y.; Ozaki, K.; Shinada, A. Metabolic rates of epipelagic marine copepods as a function of body mass and temperature. Mar. Biol. 2001, 139, 587–596. [Google Scholar] [CrossRef]
- Zweifel, U.L.; Norrman, B.; Hagstrom, A. Consumption of dissolved organic carbon by marine bacteria and demand for inorganic nutrients. Mar. Ecol. Prog. Ser. 1993, 101, 23–32. [Google Scholar] [CrossRef]
- Kroer, N. Bacterial growth efficiency on natural dissolved organic matter. Limnol. Oceanogr. 1993, 38, 1282–1290. [Google Scholar] [CrossRef]
- Goldman, J.C.; Dennett, M.R. Growth of marine bacteria in batch and continuous culture under carbon and nitrogen limitation. Limnol. Oceanogr. 2000, 45, 789–800. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Hoffman, K.A.; Weaver, R.; Hutchins, D.A. Regulation of growth and energetics of a marine bacterium by nitrogen source and iron availability. Mar. Ecol. Prog. Ser. 2003, 250, 291–296. [Google Scholar] [CrossRef]
- Goto, S.; Tada, Y.; Suzuki, K.; Yamashita, Y. Evaluation of the Production of Dissolved Organic Matter by Three Marine Bacterial Strains. Front. Microbiol. 2020, 11, 584419. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds [g/L] | Agar Plates | High Nutrient Media | Low Nutrient Media |
|---|---|---|---|
| D-(+)-Glucose (Sigma Aldrich) | 10 | 0.5 | 0.05 |
| Malt extract (Merck KGaA) | 5 | - | - |
| Yeast extract (Sigma Aldrich) | 3 | 0.5 | 0.05 |
| BactoTM Peptone (BD) | 5 | 0.5 | 0.05 |
| Agar (Sigma Aldrich) | 20 | - | - |
| Sea Salts (Sigma Aldrich) | 30 | 30 | 30 |
| Chloramphenicol (Sigma Aldrich) | 0.5 | 0.5 | 0.5 |
| Estimated Growth Phase | OD in the High Nutrient Media | OD in the Low Nutrient Media |
|---|---|---|
| Lag phase | <0.2 | <0.1 |
| Exponential phase | 0.2–0.6 | 0.1–0.154 |
| Stationary phase | >0.6 | >0.154 |
| Species | High Nutrient Media | Low Nutrient Media | ||||
|---|---|---|---|---|---|---|
| 15 °C | 5 °C | |||||
| SP1 | SP2 | SP1 | SP2 | SP1 | SP2 | |
| R. mucilaginosa | 0.31 (0.02) | 0.52 (0.04) | 0.13 (0.002) | 0.15 (0.002) | ||
| R. sphaerocarpa | 0.29 (0.01) | 0.47 (0.02) | 0.201 (0.001) | 0.29 (0.04) | 0.12 (0.004) | 0.14 (0.002) |
| S. dacryoidea | 0.23 (0.01) | 0.47 (0.004) | 0.25 (0.03) | 0.55 (0.03) | 0.109 (0.002) | 0.13 (0.002) |
| Species | Total Respiration | Cell-Specific Respiration | Production |
|---|---|---|---|
| Rhodotorula sphaerocarpa | 4.49 | 1.33 | 4.68 |
| Sakaguchia dacryoidea | 2.14 | 0.89 | 1.46 |
| Species | High Nutrient Media | Low Nutrient Media | ||||
|---|---|---|---|---|---|---|
| 15 °C | 5 °C | |||||
| SP1 | SP2 | SP1 | SP2 | SP1 | SP2 | |
| R. mucilaginosa | 1.07 × 1010 (1.28 × 109) | 1.50 × 1010 (1.9 × 109) | 3.39 × 109 (8.53 × 107) | 3.75 × 109 (2.18 × 108) | ||
| R. sphaerocarpa | 7.16 × 109 (9.63 × 108) | 1.25 × 1010 (7.13 × 108) | 1.92 × 109 (3.49 × 108) | 2.61 × 109 (1.06 × 109) | 2.97 × 109 (1.12 × 108) | 2.50 × 109 (3.60 × 108) |
| S. dacryoidea | 1.18 × 109 (2.14 × 108) | 8.39 × 109 (1.47 × 109) | 8.85 × 108 (5.22 × 108) | 3.08 × 109 (6.06 × 108) | 1.67 × 109 (1.98 × 108) | 3.01 × 109 (1.16 × 108) |
| Organism(s) | Culture Conditions | Growth Efficiency | Reference |
|---|---|---|---|
| Bacterial community of the Bothnian Sea | Batch culture with vs. without nutrients addition | 14–58% | [42] |
| 11–54% | |||
| Bacterial community from the Gulf of Mexico | Batch culture | 61% | [43] |
| Bacterial community of the coast off Massachusetts | Batch culture vs. Continuous culture | 34–70% | [44] |
| 43–58% | |||
| Marine bacterial isolate (Vibiro harveyi) | Pure culture with added glucose and iron | 45–55% | [45] |
| Bacterial community of the Mediterranean | Seawater cultures with added phosphorus | 17–70% | [33] |
| Marine bacterial isolates (Roseobacter and Cytophaga) | Pure cultures in seawater and added Zobell media | 50–70% | [34] |
| Marine bacterial isolates (Vibrio splendidus and Phaeobacter gallaeciensis) | Batch cultures with glucose as carbon source | 9–25% | [46] |
| Marine fungal isolates (Basidiomycota) | Pure cultures in defined growth media | 43–87% | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heitger, M.; Baltar, F. Respiration, Production, and Growth Efficiency of Marine Pelagic Fungal Isolates. J. Fungi 2023, 9, 417. https://doi.org/10.3390/jof9040417
Heitger M, Baltar F. Respiration, Production, and Growth Efficiency of Marine Pelagic Fungal Isolates. Journal of Fungi. 2023; 9(4):417. https://doi.org/10.3390/jof9040417
Chicago/Turabian StyleHeitger, Marilena, and Federico Baltar. 2023. "Respiration, Production, and Growth Efficiency of Marine Pelagic Fungal Isolates" Journal of Fungi 9, no. 4: 417. https://doi.org/10.3390/jof9040417