
Silver(I) 1,10-Phenanthroline Complexes Are Active against Fonsecaea pedrosoi Viability and Negatively Modulate Its Potential Virulence Attributes
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Growth Conditions
2.2. Test Compounds
2.3. Antifungal Susceptibility Test
2.4. Effect of Phen and Its Silver Complexes on F. pedrosoi Biofilm
2.5. Effect of Silver Complexes on F. pedrosoi Differentiation
2.6. Effect of Silver Complexes on the Enzymatic Activities of F. pedrosoi
2.6.1. Extracellular Metallo- and Aspartic Peptidase Activities
2.6.2. Ectophosphatase Activity
2.6.3. Phospholipase and Esterase Activities
2.7. Effect of Silver Complexes on F. pedrosoi Melanin Production
2.8. Effect of Silver Complexes on Induction of ROS in F. pedrosoi
2.9. Statistical Analysis
3. Results and Discussion
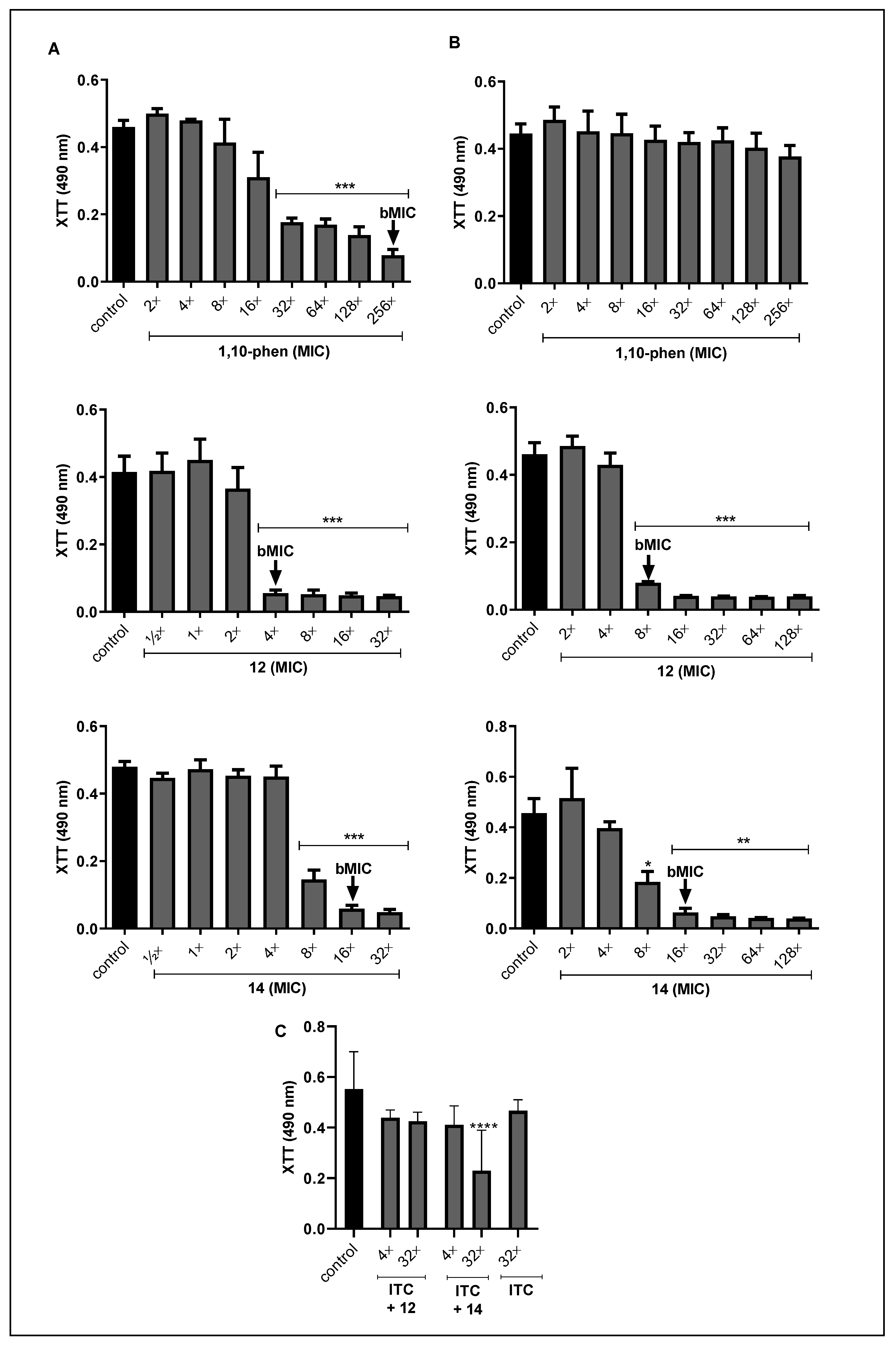
3.1. Effect of Compounds on the Viability of Fonsecaea spp.
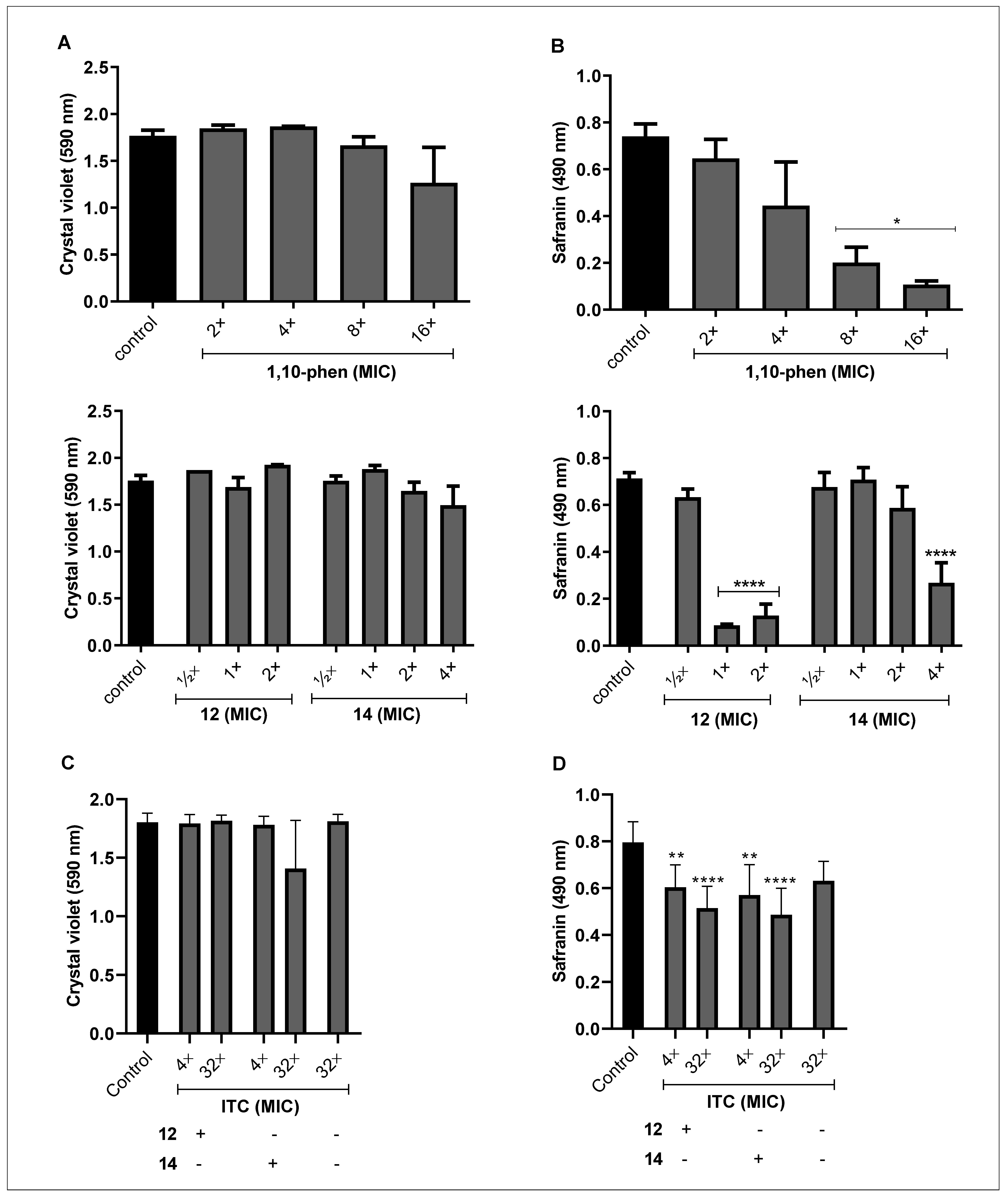
3.2. Effect of Phen and Its Silver Complexes on the Biofilm Formation and Maturation in F. pedrosoi
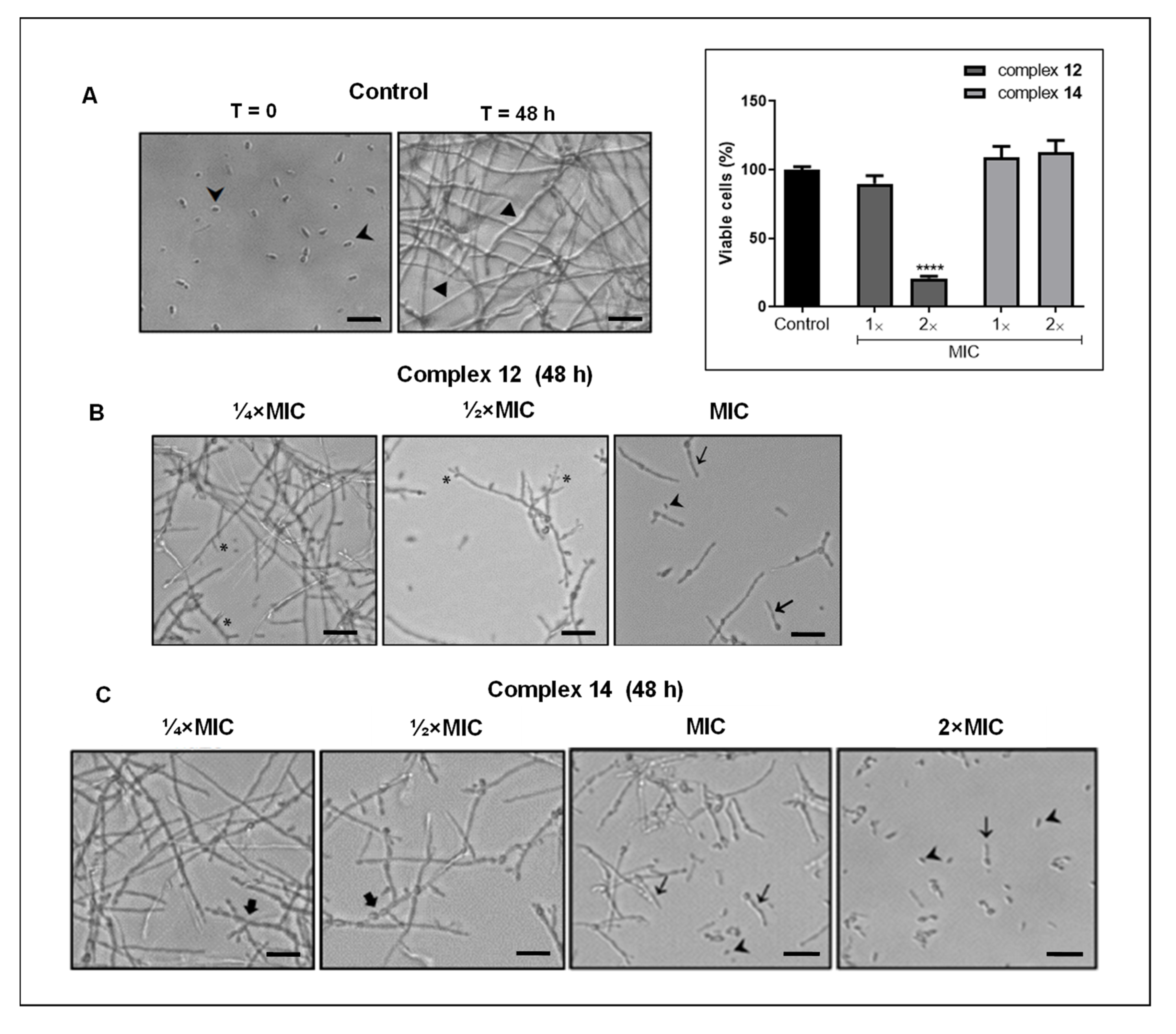
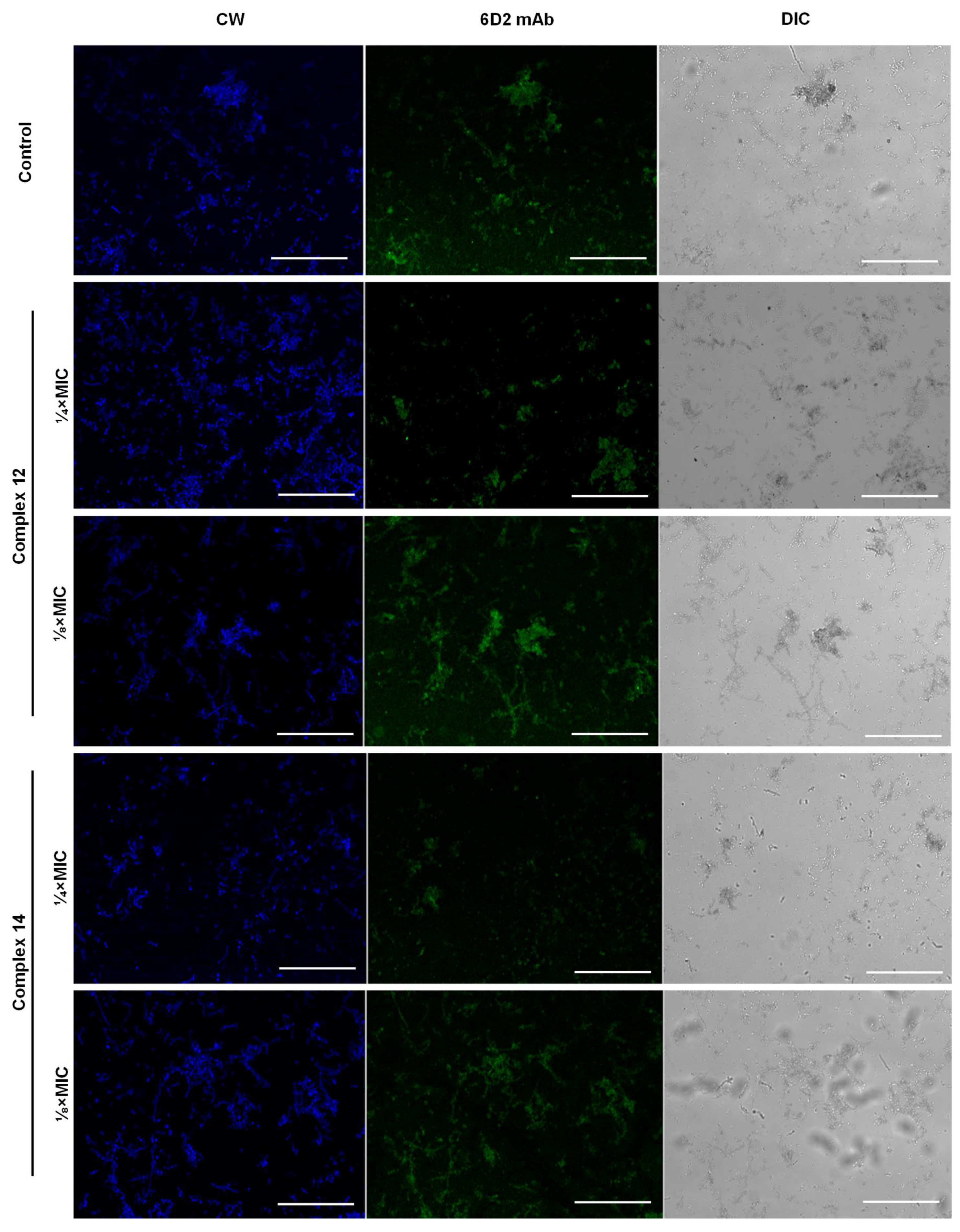
3.3. Effect of Silver Complexes on F. pedrosoi Differentiation
3.4. Effect of Silver Complexes on Enzymes Produced by F. pedrosoi
3.5. Effect of Silver Complexes on the Melanin Production by F. pedrosoi
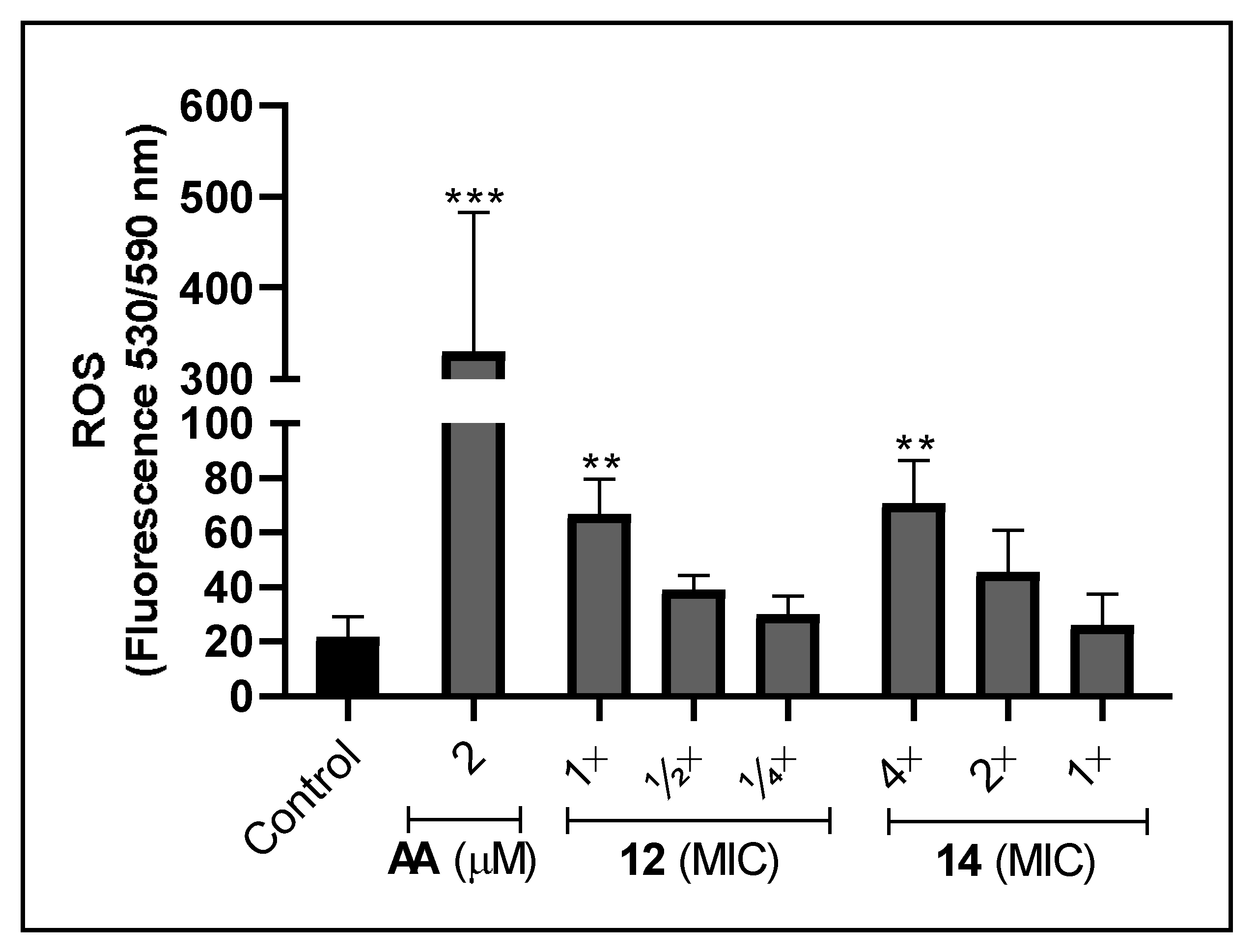
3.6. Effect of Silver Complexes on the Production of ROS in F. pedrosoi
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Tenth Report of the Strategic and Technical Advisory Group for Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Queiroz-Telles, A.F.; De Hoog, S.; Santos, D.W.C.L.; Salgado, C.G.; Vicente, V.A.; Bonifaz, A.; Roilides, E.; Liyan, X.; Azevedo, C.M.P.S.; Silva, M.B.; et al. Chromoblastomycosis. Clin. Microbiol. Rev. 2017, 30, 233–276. [Google Scholar] [CrossRef]
- Brito, A.C.; Bittencourt, M.J.S. Chromoblastomycosis: An etiological, epidemiological, clinical, diagnostic, and treatment update. An. Bras. Dermatol. 2018, 93, 495–506. [Google Scholar] [CrossRef]
- Santos, D.W.C.L.; Azevedo, C.M.P.E.S.; Vicente, V.A.; Queiroz-Telles, F.; Rodrigues, A.M.; De Hoog, S.; Denning, D.W.; Colombo, A.L. The global burden of chromoblastomycosis. PLoS Negl. Trop. Dis. 2021, 15, e0009611. [Google Scholar] [CrossRef]
- Viganor, L.; Howe, O.; Mccarron, P.; Mccann, M.; Devereux, M. The antibacterial activity of metal complexes containing 1,10- phenanthroline: Potential as alternative therapeutics in the era of antibiotic resistance. Curr. Top. Med. Chem. 2017, 17, 1280–1302. [Google Scholar] [CrossRef]
- Alviano, D.S.; Franzen, A.J.; Travassos, L.R.; Holandino, C.; Rozental, S.; Ejzemberg, R.; Alviano, C.S.; Rodrigues, M.L. Melanin from Fonsecaea pedrosoi induces production of human antifungal antibodies and enhances the antimicrobial efficacy of phagocytes. Infect. Immun. 2004, 72, 229–237. [Google Scholar] [CrossRef]
- Bocca, A.L.; Brito, P.P.M.S.; Figueiredo, F.; Tosta, C.E. Inhibition of nitric oxide production by macrophages in chromoblastomycosis: A role for Fonsecaea pedrosoi melanin. Mycopathologia 2006, 161, 195–203. [Google Scholar] [CrossRef]
- Pinto, L.; Granja, L.F.Z.; Almeida, M.A.; Alviano, D.S.; Silva, M.H.; Ejzemberg, R.; Rozental, S.; Alviano, C.S. Melanin particles isolated from the fungus Fonsecaea pedrosoi activates the human complement system. Mem. Inst. Oswaldo Cruz 2018, 113, e180120. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Palmeira, V.F.; Pinheiro, A.A.S.; Alviano, C.S.; Rozental, S.; Travassos, L.R.; Meyer-Fernandes, J.R. Phosphatase activity on the cell wall of Fonsecaea pedrosoi. Med. Mycol. 2003, 41, 469–477. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Rodrigues, M.L.; Holandino, C.; Esteves, F.F.; Souto-Padrón, T.; Alviano, C.S.; Travassos, L.R.; Meyer-Fernades, J.R. Ectophosphatase activity in conidial forms of Fonsecaea pedrosoi is modulated by exogenous phosphate and influences fungal adhesion to mammalian cells. Microbiology 2004, 150, 3355–3362. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Kneipp, L.F.; Alviano, C.S.; Santos, A.L.S. The major chromoblastomycosis fungal pathogen, Fonsecaea pedrosoi, extracellularly releases proteolytic enzymes whose expression is modulated by culture medium composition: Implications on the fungal development and cleavage of key’s host structures. FEMS Immunol. Med. Microbiol. 2006, 46, 21–29. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Kneipp, L.F.; Alviano, C.S.; Santos, A.L.S. Secretory aspartyl peptidase activity from mycelia of the human fungal pathogen Fonsecaea pedrosoi: Effect of HIV aspartyl proteolytic inhibitors. Res. Microbiol. 2006, 157, 819–826. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Kneipp, L.F.; Rozental, S.; Alviano, C.S.; Santos, A.L.S. Beneficial effects of HIV peptidase inhibitors on Fonsecaea pedrosoi: Promising compounds to arrest key fungal biological processes and virulence. PLoS ONE 2008, 3, e3382. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Kneipp, L.F.; Alviano, C.S. Phospholipase and esterase production by clinical strains of Fonsecaea pedrosoi and their interactions with epithelial cells. Mycopathologia 2010, 170, 31–37. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Alviano, D.S.; Braga-Silva, L.A.; Goulart, F.R.V.; Granato, M.Q.; Rozental, S.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. HIV Aspartic peptidase inhibitors modulate surface molecules and enzyme activities involved with physiopathological events in Fonsecaea pedrosoi. Front. Microbiol. 2017, 8, 918. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Goulart, F.R.V.; Granato, M.Q.; Alviano, D.S.; Alviano, C.S.; Kneipp, L.F.; Santos, A.L.S. Fonsecaea pedrosoi sclerotic cells: Secretion of aspartic-type peptidase and susceptibility to peptidase inhibitors. Front. Microbiol. 2018, 9, 1383. [Google Scholar] [CrossRef]
- Sousa, I.S.; Mello, P.; Pereira, E.P.; Granato, M.Q.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. Biofilm Formation by chromoblastomycosis fungi Fonsecaea pedrosoi and Phialophora verrucosa: Involvement with antifungal resistance. J. Fungi 2022, 8, 963. [Google Scholar] [CrossRef]
- Granato, M.Q.; Massapust, P.A.; Rozental, S.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. 1,10-Phenanthroline inhibits the metallopeptidase secreted by Phialophora verrucosa and modulates its growth, morphology and differentiation. Mycopathologia 2015, 179, 231–242. [Google Scholar] [CrossRef]
- Granato, M.Q.; Gonçalves, D.S.; Seabra, S.H.; McCann, M.; Devereux, M.; Santos, A.L.S.; Kneipp, L.F. 1,10-phenanthroline-5,6-dione–based compounds are effective in disturbing crucial physiological events of Phialophora verrucosa. Front. Microbiol. 2017, 8, 76. [Google Scholar] [CrossRef]
- Granato, M.Q.; Sousa, I.S.; Rosa, T.L.S.A.; Gonçalves, D.S.; Seabra, S.H.; Alviano, D.S.; Pessolani, M.C.V.; Santos, A.L.S.; Kneipp, L.F. Aspartic peptidase of Phialophora verrucosa as target of HIV peptidase inhibitors: Blockage of its enzymatic activity and interference with fungal growth and macrophage interaction. J. Enzyme Inhib. Med. Chem. 2020, 35, 629–638. [Google Scholar] [CrossRef]
- Granato, M.Q.; Mello, T.P.; Nascimento, R.S.; Pereira, M.D.; Rosa, T.L.S.A.; Pessolani, M.C.V.; McCann, M.; Devereux, M.; Branquinha, M.H.; Santos, A.L.S.; et al. Silver(I) and copper(II) complexes of 1,10-phenanthroline-5,6-dione against Phialophora verrucosa: A focus on the interaction with human macrophages and Galleria mellonella larvae. Front. Microbiol. 2021, 12, 641258. [Google Scholar] [CrossRef]
- Santos, A.L.S. Aspartic proteases of human pathogenic fungi are prospective targets for the generation of novel and effective antifungal inhibitors. Curr. Enzym. Inhib. 2011, 7, 96–118. [Google Scholar] [CrossRef]
- Santos, A.L.S.; Braga-silva, L.A.; Gonçalves, D.S.; Ramos, L.S.; Oliveira, S.S.C.; Souza, L.O.P.; Oliveira, V.S.; Lins, R.D.; Pinto, M.R.; Muñoz, J.E.; et al. Repositioning lopinavir, an HIV protease inhibitor, as a promising antifungal drug: Lessons learned from Candida albicans- in silico, in vitro and in vivo approaches. J. Fungi 2021, 7, 424. [Google Scholar] [CrossRef]
- McCann, M.; Kellett, A.; Kavanagh, K.; Devereux, M.; Santos, A.L.S. Deciphering the antimicrobial activity of phenanthroline chelators. Curr. Med. Chem. 2012, 19, 2703–2714. [Google Scholar] [CrossRef]
- Santos, A.L.S.; Sodré, C.L.; Valle, R.S.; Silva, B.A.; Silva, L.V.; Sangenito, L.S.; Gonçalves, D.S.; Souza, L.O.P.; Palmeira, V.F. Antimicrobial action of chelating agents: Repercussions on the microorganism development, virulence and pathogenesis. Curr. Med. Chem. 2012, 19, 2715–2737. [Google Scholar] [CrossRef]
- Bencini, A.; Lippolis, V. 1,10-phenanthroline: A versatile building block for the construction of ligands for various purposes. Coord. Chem. Rev. 2010, 254, 2096–2180. [Google Scholar] [CrossRef]
- Wu, F.; Xie, J.; Zhu, Z. 1,10-phenanthroline: A versatile ligand to promote copper-catalyzed cascade reactions. Appl. Organomet. Chem. 2020, 34, e5926. [Google Scholar] [CrossRef]
- Raman, N.; Mahalakshmi, R.; Mitu, L. Bio-sensitive activities of coordination compounds containing 1,10-phenanthroline as co-ligand: Synthesis, structural elucidation and DNA binding properties of metal(II) complexes. Spectrochim. Acta—Part A Mol. Biomol. Spectrosc. 2014, 131, 355–364. [Google Scholar] [CrossRef]
- Đurić, S.; Mojicevic, M.; Vojnovic, S.; Wadepohl, H.; Andrejević, T.P.; Stevanović, N.L.; Nikodinovic-Runic, J.; Djuran, M.I.; Glišić, B. Silver(I) complexes with 1,10-phenanthroline-based ligands: The influence of epoxide function on the complex structure and biological activity. Inorganica Chim. Acta 2020, 502, 119357. [Google Scholar] [CrossRef]
- Lin, Y.; Betts, H.; Keller, S.; Cariou, K.; Gasser, G. Recent developments of metal-based compounds against fungal pathogens. Chem. Soc. Rev. 2021, 50, 10346–10402. [Google Scholar] [CrossRef]
- Geraghty, M.; Cronin, J.F.; Devereux, M.; Mccann, M. Synthesis and antimicrobial activity of copper (II) and manganese (II) α, ω -dicarboxylate complexes. BioMetals 2000, 13, 1–8. [Google Scholar] [CrossRef]
- McCann, M.; Geraghty, M.; Devereux, M.; O’Shea, D.; Mason, J.; O’Sullivan, L. Insights into the mode of action of the anti-candida activity of 1,10-phenanthroline and its metal chelates. Met. Based. Drugs 2000, 7, 185–193. [Google Scholar] [CrossRef]
- Coyle, B.; Kavanagh, K.; Mccann, M.; Devereux, M.; Geraghty, M. Mode of anti-fungal activity of 1,10-phenanthroline and its Cu (II), Mn (II) and Ag (I) complexes. BioMetals 2003, 16, 321–329. [Google Scholar] [CrossRef]
- McCann, M.; Coyle, B.; McKay, S.; McCormack, P.; Kavanagh, K.; Devereux, M.; McKee, V.; Kinsella, P.; O’Connor, R.; Clynes, M. Synthesis and X-ray crystal structure of [Ag(phendio)2]ClO4 (phendio=1,10-phenanthroline-5,6-dione) and its effects on fungal and mammalian cells. BioMetals 2004, 17, 635–645. [Google Scholar] [CrossRef]
- Hoffman, A.E.; Miles, L.; Greenfield, T.J.; Shoen, C.; Destefano, M.; Cynamon, M.; Doyle, R.P. Clinical isolates of Candida albicans, Candida tropicalis, and Candida krusei have different susceptibilities to Co(II) and Cu(II) complexes of 1,10-phenanthroline. BioMetals 2015, 28, 415–423. [Google Scholar] [CrossRef]
- Gandra, R.M.; Carron, P.M.; Fernandes, M.F.; Ramos, L.S.; Mello, T.P.; Aor, A.C.; Branquinha, M.H.; McCann, M.; Devereux, M.; Santos, A.L.S. Antifungal potential of copper(II), Manganese(II) and silver(I) 1,10-phenanthroline chelates against multidrug-resistant fungal species forming the Candida haemulonii complex: Impact on the planktonic and biofilm lifestyles. Front. Microbiol. 2017, 8, 1257. [Google Scholar] [CrossRef]
- Coyle, B.; Kinsella, P.; McCann, M.; Devereux, M.; O’Connor, R.; Clynes, M.; Kavanagh, K. Induction of apoptosis in yeast and mammalian cells by exposure to 1,10-phenanthroline metal complexes. Toxicol. Vitr. 2004, 18, 63–70. [Google Scholar] [CrossRef]
- McCann, M.; Santos, A.L.S.; Silva, B.A.; Romanos, M.T.V.; Pyrrho, A.S.; Devereux, M.; Kavanagh, K.; Fichtner, I.; Kellett, A. In vitro and in vivo studies into the biological activities of 1,10-phenanthroline, 1,10-phenanthroline-5,6-dione and its copper(ii) and silver(i) complexes. Toxicol. Res. (Camb) 2012, 1, 47–54. [Google Scholar] [CrossRef]
- Gandra, R.M.; McCarron, P.; Viganor, L.; Fernandes, M.F.; Kavanagh, K.; McCann, M.; Branquinha, M.H.; Santos, A.L.S.; Howe, O.; Devereux, M. In vivo activity of copper(II), manganese(II), and silver(I) 1,10-phenanthroline chelates against Candida haemulonii using the Galleria mellonella model. Front. Microbiol. 2020, 11, 470. [Google Scholar] [CrossRef]
- Ronconi, L.; Sadler, P.J. Using coordination chemistry to design new medicines. Coord. Chem. Rev. 2007, 251, 1633–1648. [Google Scholar] [CrossRef]
- Frei, A.; Elliott, A.G.; Kan, A.; Dinh, H.; Br, S.; Bruce, A.E.; Bruce, M.R.; Chen, F.; Humaidy, D.; Jung, N.; et al. Metal complexes as antifungals? From a crowd-sourced compound library to the first. J. Am. Chem. Soc. 2022, 2, 2277–2294. [Google Scholar] [CrossRef]
- Boros, E.; Dyson, P.J.; Gasser, G. Classification of metal-based drugs according to their mechanisms of action. Chem 2020, 6, 41–60. [Google Scholar] [CrossRef]
- Devereux, M.; Mccann, M.; Leon, V.; Geraghty, M.; Mckee, V.; Wikaira, J. Syntesis and biological activity of manganese (II) complexes of phthalic and isophthalic acid: X-Ray crystal structures of [Mn(ph)(phen)2(H2O)]·4H2O, [Mn(phen)2(H2O2)]2(isoph)(phen)·12H2O and {[Mn(isoph)(bipy)]4·2.75bipy}n (phH2 = phthalic acid; isoph = I. Met. Based Drugs 2000, 7, 275–288. [Google Scholar] [CrossRef]
- Kellett, A.; Howe, O.; O’Connor, M.; McCann, M.; Creaven, B.S.; McClean, S.; Foltyn-Arfa Kia, A.; Casey, A.; Devereux, M. Radical-induced DNA damage by cytotoxic square-planar copper(II) complexes incorporating o-phthalate and 1,10-phenanthroline or 2,2′-dipyridyl. Free Radic. Biol. Med. 2012, 53, 564–576. [Google Scholar] [CrossRef]
- Kellett, A.; Connor, M.O.; Mccann, M.; Mcnamara, M.; Lynch, P.; Rosair, G.; Mckee, V.; Creaven, B.; Walsh, M.; Mcclean, S.; et al. Bis-phenanthroline copper(ii) phthalate complexes are potent in vitro antitumour agents with ‘self-activating’ metallo-nuclease and dna binding properties. Dalt. Trans. 2011, 40, 1024–1027. [Google Scholar] [CrossRef]
- Salvadores, V.L. Synthesis, characterization and catalytic and biological activity of new manganase(II) carboxylate complexes. Doctoral thesis, Technological University Dublin, Dublin, Ireland, 2000. [Google Scholar] [CrossRef]
- Casey, M.T.; Mccann, M.; Devereux, M.; Curran, M.; Cardin, C.; Harding, C. Synthesis and structure of the Mn2 (II, II) Complex Salt [Mn2(oda)(phen)4(H2O)2] [Mn2(oda)2(phen)4] (odaH2=octanedioic acid): A catalyst for H2O2 disproportionation. Chem. Soc. Chem. Commun. 1994, 2, 2643–2645. [Google Scholar] [CrossRef]
- Devereux, M.; McCann, M.; Cronin, J.; Ferguson, G.; McKee, V. Binuclear and polymeric copper (II) dicarboxylate complexes: Synthesis and crystal structures of [Cu2(pda)(phen)4](ClO4)2·5H2O·EtOH, [Cu2(oda)(phen)4](ClO4)2·2.67H2O·EtOH and {Cu2(pda)2(NH3)4(H2O)2·4H2O}n (odaH2=octanedioic acid; pdaH2= pentanedioic Ac. Polyhedron 1999, 18, 2141–2148. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method M38-A2 for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 2nd ed.; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2008. [Google Scholar]
- Liu, M.; Seidel, V.; Katerere, D.R.; Gray, A.I. Colorimetric broth microdilution method for the antifungal screening of plant extracts against yeasts. Methods. 2007, 42, 325–329. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Sheehan, D.J.; Rex, J.H. Determination of fungicidal activities against yeasts and molds: Lessons Learned from bactericidal testing and the need for standardization. Clin. Microbiol. Rev. 2004, 17, 268–280. [Google Scholar] [CrossRef]
- Mowat, E.; Butcher, J.; Lang, S.; Williams, C.; Ramage, G. Development of a simple model for studying the effects of antifungal agents on multicellular communities of Aspergillus fumigatus. J. Med. Microbiol. 2007, 56, 1205–1212. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Twining, S. Fluorescein isothiocyanate-labeled casein assay for proteolytic. Anal. Biochem. 1984, 143, 30–34. [Google Scholar] [CrossRef]
- Santos, L.O.; Vitorio, B.S.; Branquinha, M.H.; Silva, C.M.P.; Santos, A.L.S.; D’Avila-Levy, C.M. Nelfinavir is effective in inhibiting the multiplication and aspartic peptidase activity of Leishmania species, including strains obtained from HIV-positive patients. J. Antimicrob. Chemother. 2013, 68, 348–353. [Google Scholar] [CrossRef]
- Macêdo-sales, P.A.; Olegario, L.; Souza, P.; Della-terra, P.P.; Lozoya-pérez, N.E.; Luiz, R.; Machado, D.; Martins, E.; Lopes-bezerra, L.M.; Guimarães, A.J.; et al. Coinfection of domestic felines by distinct Sporothrix brasiliensis in the Brazilian sporotrichosis hyperendemic area. Fungal Genet. Biol. 2020, 140, 103397. [Google Scholar] [CrossRef]
- Votyakova, T.V.; Reynolds, I.J. Detection of hydrogen peroxide with Amplex Red: Interference by NADH and reduced glutathione auto-oxidation. Arch. Biochem. Biophys. 2004, 431, 138–144. [Google Scholar] [CrossRef]
- O’Shaughnessy, M.; Piatek, M.; Mccarron, P.; Mccann, M.; Devereux, M.; Kavanagh, K.; Howe, O. In vivo activity of metal complexes containing Pseudomonas aeruginosa infection in Galleria mellonella larvae. Biomed. Artic. 2022, 10, 1–21. [Google Scholar]
- McCarron, P.; McCann, M.; Devereux, M.; Kavanagh, K.; Skerry, C.; Karakousis, P.C.; Aor, A.C.; Mello, T.P.; Santos, A.L.S.; Campos, D.L.; et al. Antitubercular activitiy of manganese (II) complexes containing dicarboxylate ligands: Increased activity, superior selectivity, and lower toxicity in comparison to their copper(II) analogs. Front. Microbiol. 2018, 9, 1432. [Google Scholar] [CrossRef]
- O’Shaughnessy, M.; McCarron, P.; Viganor, L.; McCann, M.; Devereux, M.; Howe, O. The antibacterial and anti-biofilm activity of metal complexes incorporating 3,6,9-trioxaundecanedioate and 1,10-phenanthroline ligands in clinical isolates of Pseudomonas aeruginosa from irish cystic fibrosis patients. Antibiotics 2020, 9, 674. [Google Scholar] [CrossRef]
- Accorsi, G.; Listorti, A.; Yoosaf, K.; Armaroli, N. 1,10-phenanthrolines: Versatile building blocks for luminescent molecules, materials and metal complexes. Chem. Soc. Rev. 2009, 38, 1690–1700. [Google Scholar] [CrossRef]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C.; Hospital, R.; Yorkhill, C.; Street, D.; Glasgow, G. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. The extracellular matrix of fungal biofilms. Adv. Exp. Med. Biol. 2016, 931, 21–35. [Google Scholar] [CrossRef]
- Zhang, K.; Li, X.; Yu, C.; Wang, Y.; Wang, Y. Promising therapeutic strategies against microbial biofilm challenges. Front. Cell Infect. Microbiol. 2020, 10, 359. [Google Scholar] [CrossRef]
- Eshwika, A.; Coyle, B.; Devereux, M.; Mccann, M.; Kavanagh, K. Metal complexes of 1,10-phenanthroline-5,6-dione alter the susceptibility of the yeast Candida albicans to amphotericin B and miconazole. BioMetals 2004, 17, 415–422. [Google Scholar] [CrossRef]
- Chudzik, B.; Tracz, I.B.; Czernel, G.; Fiołka, M.J.; Borsuk, G.; Gagos, M. Amphotericin B–copper(II) complex as a potential agent with higher antifungal activity against Candida albicans. Eur. J. Pharm. Sci. 2013, 49, 850–857. [Google Scholar] [CrossRef]
- Tits, J.; Cammue, B.P.A.; Thevissen, K. Combination therapy to treat fungal biofilm-based infections. Int. J. Mol. Sci. 2020, 21, 8873. [Google Scholar] [CrossRef]
- Ramage, G.; Vandewalle, K.; López-Ribot, J.L.; Wickes, B.L. The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. FEMS Microbiol. Lett. 2002, 214, 95–100. [Google Scholar] [CrossRef]
- Tan, X.; Fuchs, B.B.; Wang, Y.; Chen, W.; Yuen, G.J.; Chen, R.B.; Jayamani, E.; Anastassopoulou, C.; Pukkila-worley, R.; Coleman, J.J.; et al. The Role of Candida albicans SPT20 in filamentation, biofilm formation and pathogenesis. PLoS ONE 2014, 9, e94468. [Google Scholar] [CrossRef]
- Nemecek, J.C.; Wüthrich, M.; Klein, B.S. Global control of dimorphism and virulence in fungi. Science 2006, 312, 583–588. [Google Scholar] [CrossRef]
- Lin, X.; Alspaugh, J.A.; Liu, H.; Harris, S. Fungal morphogenesis. Cold Spring Harb. Perspect. Med. 2015, 5, a019679. [Google Scholar] [CrossRef]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis definition of phospholipases and rationale in considering them for. Clin. Microbiol. Rev. 2000, 13, 122–143. [Google Scholar] [CrossRef]
- Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R. Biochemical properties and possible roles of ectophosphatase activities in fungi. Int. J. Mol. Sci. 2014, 15, 2289–2304. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.S.; Palmeira, V.F.; Rozental, S.; Kneipp, L.F.; Nimrichter, L.; Alviano, D.S.; Rodrigues, M.L.; Alviano, C.S. Biology and pathogenesis of Fonsecaea pedrosoi, the major etiologic agent of chromoblastomycosis. FEMS Microbiol. Rev. 2007, 31, 570–591. [Google Scholar] [CrossRef] [PubMed]
- Kellett, A.; Prisecaru, A.; Slator, C.; Molphy, Z. Metal-based antimicrobial protease inhibitors. Curr. Med. Chem. 2013, 20, 3134–3151. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhu, M. Protein tyrosine phosphatase inhibition by metals and metal complexes. Antioxidants Redox Signal. 2014, 20, 2210–2224. [Google Scholar] [CrossRef]
- Collins, J.; Cilibrizzi, A.; Fedorova, M.; Whyte, G.; Mak, L.H.; Guterman, I.; Leatherbarrow, R.; Woscholski, R.; Vilar, R. Vanadyl complexes with dansyl-labelled di-picolinic acid ligands: Synthesis, phosphatase inhibition activity and cellular uptake studies. Dalt. Trans. 2016, 45, 7104–7113. [Google Scholar] [CrossRef]
- Radisavljević, M.; Kamčeva, T.; Bugarčić, Ž.D.; Petković, M. Inhibitory effect of cisplatin and [Pt(dach)Cl2] on the activity of phospholipase A2. J. Enzyme Inhib. Med. Chem. 2012, 28, 651–660. [Google Scholar] [CrossRef]
- Nešić, M.; Popović, I.; Leskovac, A.; Petković, M. Biological activity and binding properties of [Ru(II)(dcbpy)2Cl2] complex to bovine serum albumin, phospholipase A2 and glutathione. BioMetals 2016, 29, 921–933. [Google Scholar] [CrossRef]
- Ilkimen, H.; Yenikaya, C.; Sari, M.; Bülbül, M.; Aslan, M.; Süzen, Y. Synthesis and characterization of some metal complexes of a proton transfer salt, and their inhibition studies on carbonic anhydrase isozymes and the evaluation of the results by statistical analysis. J. Enzyme Inhib. Med. Chem. 2014, 29, 695–701. [Google Scholar] [CrossRef]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef]
- Rossi, D.C.P.; Spadari, C.D.C.; Nosanchuk, J.D.; Taborda, C.P.; Ishida, K. Miltefosine is fungicidal to Paracoccidioides spp. yeast cells but subinhibitory concentrations induce melanisation. Int. J. Antimicrob. Agents 2017, 49, 465–471. [Google Scholar] [CrossRef]
- Heidrich, D.; Pagani, D.M.; Koehler, A.; de Oliveira Alves, K.; Scrofernekera, M.L. Effect of melanin biosynthesis inhibition on the antifungal susceptibility of chromoblastomycosis agents. Antimicrob. Agents Chemother. 2021, 65, e0054621. [Google Scholar] [CrossRef] [PubMed]
- Coelho, R.A.; Figueiredo-Carvalho, M.H.G.; Silva, J.V.S.; Correa-Junior, D.; Frases, S.; Zancopé-Oliveira, R.M.; Freitas, D.F.S.; Almeida-Paes, R. Does DHN-melanin always protect fungi against antifungal drugs? The Fonsecaea/micafungin paradigma. Microbiol. Res. 2022, 13, 201–209. [Google Scholar] [CrossRef]
- Ribeiro, T.P.; Fernandes, C.; Melo, K.V.; Ferreira, S.S.; Lessa, J.A.; Franco, R.W.A.; Schenk, G.; Pereira, M.D.; Horn, A. Free radical biology and medicine iron, copper, and manganese complexes with in vitro superoxide dismutase and/or catalase activities that keep Saccharomyces cerevisiae cells alive under severe oxidative stress. Free Radic. Biol. Med. 2015, 80, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide James. Ann. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Li, M.; Wang, H.; Dong, X. Metal complexes or chelators with ROS regulation capacity: Promising candidates for cancer treatment. Molecules 2022, 27, 148. [Google Scholar] [CrossRef]
- Rowan, R.; McCann, M.; Kavanagh, K. Analysis of the response of Candida albicans cells to Silver(I). Med. Mycol. 2010, 48, 498–505. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complexes | Synthesis References |
|---|---|
| Phthalic acid (phH2) | |
| (1) [Mn(ph)(phen)(H2O)2] | Devereux et al., 2000 [43] |
| (2) [Cu(ph)(phen)(H2O)2] | Kellett et al., 2012 [44] |
| (3) [Cu(ph)(phen)2].3H2O.2EtOH * | Kellett et al., 2011 [45] |
| Isophthalic acid (isophH2) | |
| (4) [Mn2(isoph)2(phen)3].4H2O | Devereux et al., 2000 [43] |
| (5) [Cu(isoph)(phen)2].6H2O.EtOH | Kellett et al., 2011 [45] |
| Terephthalic acid (terephH2) | |
| (6) [Mn(tereph)(phen)2].5H2O | Salvadores 2000 [46] |
| (7) [{Cu(phen)2}2(terph)](terph).13.5H2O.2EtOH | Kellett et al., 2011 [45] |
| Octanedioic acid (odaH2) | |
| (8) [Mn2(oda)(phen)4(H2O)2][Mn2(oda)(phen)4(oda)2].4H2O | Casey et al., 1994 [47] |
| (9) [Cu2(oda)(phen)4](ClO4)2.2.76H2O.EtOH | Devereux et al., 1999 [48] |
| Perchlorate salt (ClO4) | |
| (10) [Ag(phendione)2]ClO4 | McCann et al., 2004 [34] |
| (11) [Cu(phendione)3](ClO4)2.4H2O | McCann et al., 2004 [34] |
| (12) [Ag(phen)2]ClO4 | McCann et al., 2004 [34] |
| 3,6,9-trioxaundecanedioate (3,6,9-tddaH2) | |
| (13) {[Cu(3,6,9-tdda)(phen)2].3H2O.EtOH}n | Gandra et al., 2017 [36] |
| (14) [Ag2(3,6,9-tdda)(phen)4].EtOH | Gandra et al., 2017 [36] |
| Complexes | F. monophora | F. nubica | F. pedrosoi | |
|---|---|---|---|---|
| MIC µM (mg/L) | MIC µM (mg/L) | MIC µM (mg/L) | MFC µM (mg/L) | |
| 1 | 25.0 (10.9) | 25.0 (10.9) | 6.2 (2.7) | >100.0 (>43.6) |
| 2 | 100.0 (44.4) | >100.0 (>44.4) | >100.0 (>44.4) | >100.0 (>44.4) |
| 3 | 100.0 (73.5) | 50.0 (36.8) | 50.0 (36.8) | >100.0 (>73.5) |
| 4 | ND | 12.5 (13.2) | 12.5 (13.2) | >100.0 (>105.6) |
| 5 | ND | ND | 25.0 (18.6) | >74.5 (>100.0) |
| 6 | 12.5 (8.5) | 12.5 (8.5) | 6.2 (4.2) | >100.0 (>67.1) |
| 7 | 100.0 (146.0) | 50.0 (73.0) | 100.0 (146.0) | >100.0 (>146.0) |
| 8 | ND | ND | 6.2 (15.5) | >100.0 (>248.3) |
| 9 | ND | ND | 6.2 (8.2) | >100.0 (>131.6) |
| 10 | 6.2 (3.9) | 12.5 (7.9) | 6.2 (3.9) | ND |
| 11 | 6.2 (6.0) | 3.1 (3.0) | 3.1 (3.0) | 3.1 (3.0) * |
| 12 | 0.6 (0.3) | 0.6 (0.3) | 1.2 (0.7) | 5.0 (2.8) * |
| 13 | 6.2 (4.6) | 3.1 (2.3) | 3.1 (2.3) | >100.0 (>74.5) |
| 14 | 1.2 (1.4) | 2.5 (3.0) | 0.6 (0.7) | 2.5 (3.0) * |
| Compounds | MIC µM (mg/L) | MFC µM (mg/L) |
|---|---|---|
| Phen | 3.1 (0.6) | >100.0 (>19.3) |
| Phendione | 3.1 (0.7) | >100.0 (>22.6) |
| Silver perchlorate | 6.2 (1.9) | 25.0 (7.7) * |
| Copper perchlorate | >100.0 (>37.0) | >100.0 (>37.0) |
| Silver nitrate | 6.2 (1.1) | 12.5 (2.2) * |
| Copper sulfate | >100.0 (>16.0) | >100.0 (>16.0) |
| Manganese chloride | >100.0 (>12.6) | >100.0 (>12.6) |
| Complexes | Enzyme Activities | |||||||
|---|---|---|---|---|---|---|---|---|
| Aspartic Peptidase (%) | Metallo Peptidase (%) | Ectophosphatase (%) | Phospholipase Pz Value | Esterase Pz Value | ||||
| 50 µM | 50 µM | 2×MIC | MIC | 2×MIC | MIC | 2×MIC | MIC | |
| 12 | 56.0 ± 6.2 * | 38.9 ± 5.5 * | ND | 114.4 ± 8.0 | ND | 0.70 ± 0.03 | ND | 0.65 ± 0.01 |
| 14 | 65.3 ± 3.8 * | 59.8 ± 6.9 * | 99.3 ± 6.3 | 105.0 ± 7.5 | 0.64 ± 0.03 | 0.69 ± 0.02 | 0.72 ± 0.005 | 0.69 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, I.S.; Vieira, T.D.P.; Menna-Barreto, R.F.S.; Guimarães, A.J.; McCarron, P.; McCann, M.; Devereux, M.; Santos, A.L.S.; Kneipp, L.F. Silver(I) 1,10-Phenanthroline Complexes Are Active against Fonsecaea pedrosoi Viability and Negatively Modulate Its Potential Virulence Attributes. J. Fungi 2023, 9, 356. https://doi.org/10.3390/jof9030356
Sousa IS, Vieira TDP, Menna-Barreto RFS, Guimarães AJ, McCarron P, McCann M, Devereux M, Santos ALS, Kneipp LF. Silver(I) 1,10-Phenanthroline Complexes Are Active against Fonsecaea pedrosoi Viability and Negatively Modulate Its Potential Virulence Attributes. Journal of Fungi. 2023; 9(3):356. https://doi.org/10.3390/jof9030356
Chicago/Turabian StyleSousa, Ingrid S., Tatiana D. P. Vieira, Rubem F. S. Menna-Barreto, Allan J. Guimarães, Pauraic McCarron, Malachy McCann, Michael Devereux, André L. S. Santos, and Lucimar F. Kneipp. 2023. "Silver(I) 1,10-Phenanthroline Complexes Are Active against Fonsecaea pedrosoi Viability and Negatively Modulate Its Potential Virulence Attributes" Journal of Fungi 9, no. 3: 356. https://doi.org/10.3390/jof9030356