
Simultaneous Heavy Metal-Polycyclic Aromatic Hydrocarbon Removal by Native Tunisian Fungal Species

 ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Strains
2.2. Culture Conditions
2.2.1. Phenanthrene and Benz[a]anthracene Biodegradation Experiments
2.2.2. Bioremediation of Combined Pollutants
2.2.3. Sampling
2.3. Enzymatic Activity
2.4. Chemical Analyses
2.4.1. Estimation of Glucose Content
2.4.2. Phenanthrene and Benzo[a]anthracene Quantification and Metabolite Identification
2.4.3. Heavy Metal Analysis
2.4.4. Transmission Electron Microscopy Coupled with Energy-Dispersive X-ray Spectroscopy (TEM EDX)
2.5. Toxicity Bioassays
2.5.1. Phytotoxicity Test
- G and G0: number of germinated seeds in the samples and the control;
- L and L0: root length values in the samples and the control.
2.5.2. Microtox®Test
3. Results
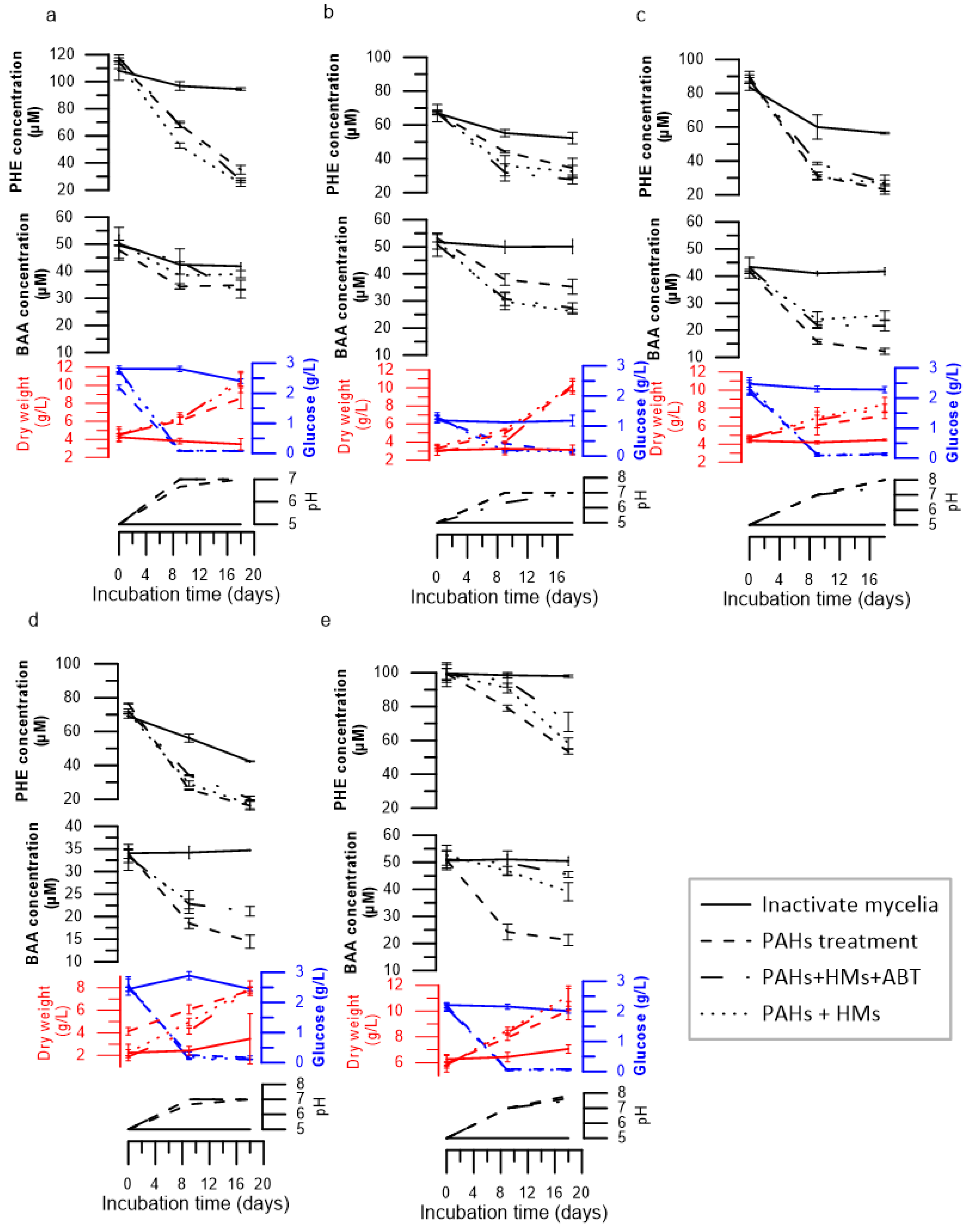
3.1. Removal of Phenanthrene and Benz[a]anthracene
3.2. Enzyme Production during Degradation
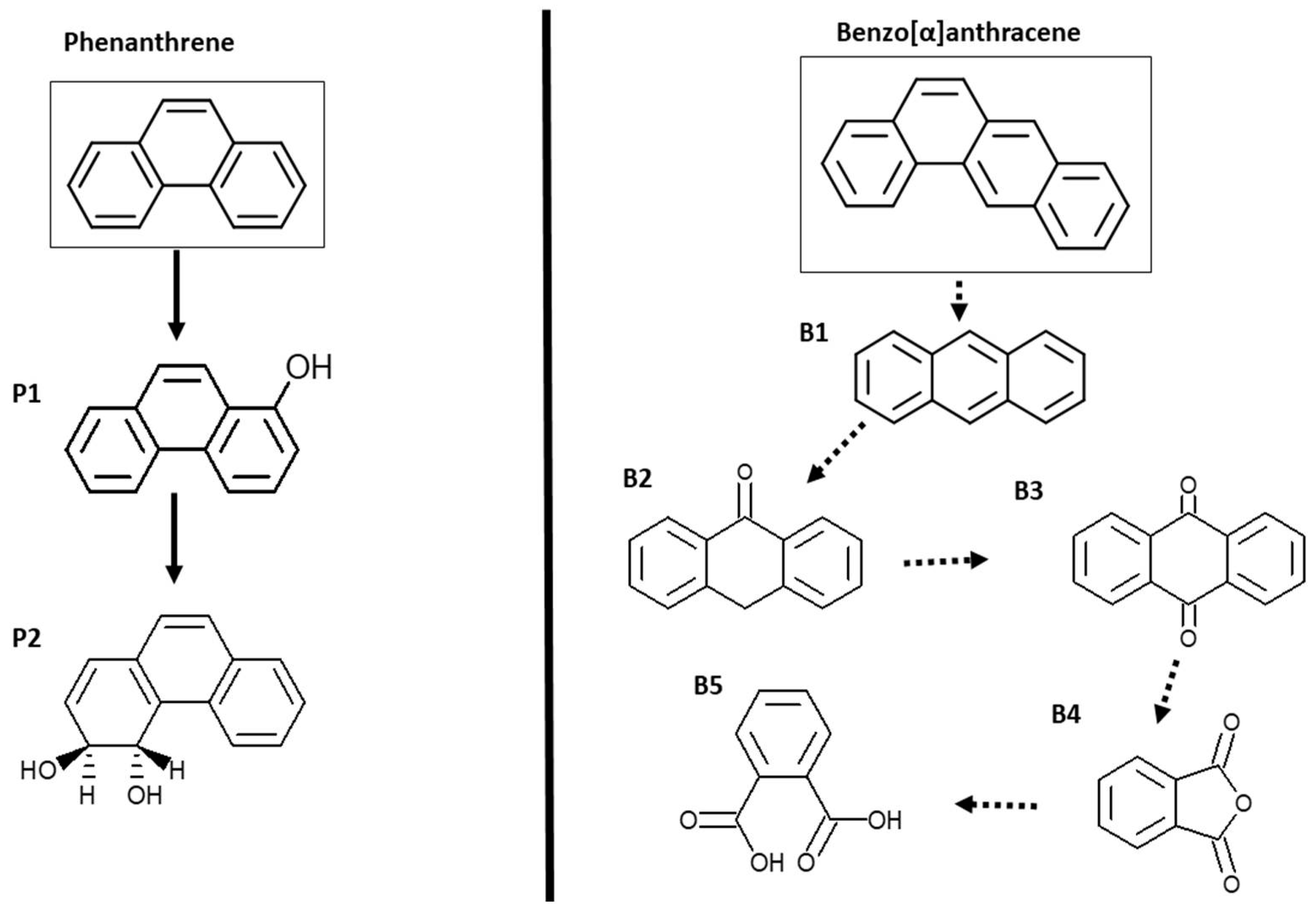
3.3. Metabolic Products after PHE and BAA Biodegradation
3.4. Heavy Metal Removal
3.4.1. Residual Heavy Metal Content
3.4.2. Transmission Electron Microscopy Coupled with EDX Analysis
3.5. Ecotoxicological Studies
3.5.1. Phytotoxicity Analysis
3.5.2. Microtox®Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Elemental Composition | Chemical Structure | Molecular Weigh | Error (ppm) | Retention Time (min) |
|---|---|---|---|---|---|
| Phenanthrene | |||||
| Original molecule | C14H10 |  | 179.0861 | 1.7 | 5.30 |
| P1 | C14H10O |  | 195.0810 | −10.3 | 0.70 |
| P2 | C14H12O2 |  | 213.0916 | −6.1 | 0.52 |
| Benz[α]anthracene | |||||
| Original molecule | C18H12 |  | 229.1017 | −4.8 | 5.90 |
| B1 | C14H10 |  | 179.0861 | 1.7 | 5.30 |
| B2 | C14H10O |  | 195.0810 | −1.5 | 5.72 |
| B3 | C14H8O2 |  | 209.0603 | 2.4 | 6.84 |
| B4 | C8H4O3 |  | 149.0239 | −9.4 | 6.31 |
| B5 | C8H6O4 |  | 167.0344 | −3.6 | 6.27 |
References
- Arul, M.N.; Alemu, A.K.; Goswami, L.; Pakshirajan, K.; Pugazhenthi, G. Waste litchi peels for Cr (VI) removal from synthetic wastewater in batch and continuous systems: Sorbent characterization, regeneration and reuse study. J. Environ. Eng. 2016, 142, C4016001. [Google Scholar] [CrossRef]
- Titaley, I.A.; Chlebowski, A.; Truong, L.; Tanguay, R.L.; Massey Simonich, S.L. Identification and toxicological evaluation of unsubstituted PAHs and novel PAH derivatives in pavement sealcoat products. Environ. Sci. Tech. Let. 2016, 3, 234–242. [Google Scholar] [CrossRef]
- Wang, J.; Deng, P.; Wei, X.; Zhang, X.; Liu, J.; Huang, Y.; She, J.; Liu, Y.; Wan, Y.; Hu, H.; et al. Hidden risks from potentially toxic metal (loid) s in paddy soils-rice and source apportionment using lead isotopes: A case study from China. Sci. Total Environ. 2023, 856, 158883. [Google Scholar] [CrossRef]
- Ali, M.; Song, X.; Ding, D.; Wang, Q.; Zhang, Z.; Tang, Z. Bioremediation of PAHs and heavy metals co-contaminated soils: Challenges and enhancement strategies. Environ. Pollut. 2021, 295, 118686. [Google Scholar] [CrossRef] [PubMed]
- Kadri, T.; Rouissi, T.; Brar, S.K.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Janicki, T.; Długoński, J.; Krupiński, M. Detoxification and simultaneous removal of phenolic xenobiotics and heavy metals with endocrine-disrupting activity by the non-ligninolytic fungus Umbelopsisisabellina. J. Hazard. Mater. 2018, 360, 661–669. [Google Scholar] [CrossRef]
- Deshmukh, R.; Khardenavis, A.A.; Purohit, H.J. Diverse metabolic capacities of fungi for bioremediation. Indian J. Microbiol. 2016, 56, 247–264. [Google Scholar] [CrossRef] [Green Version]
- Priyadarshini, E.; Priyadarshini, S.S.; Cousins, B.G.; Pradhan, N. Metal-Fungus interaction: Review on cellular processes underlying heavy metal detoxification and synthesis of metal nanoparticles. Chemosphere 2021, 274, 129976. [Google Scholar] [CrossRef]
- Al-Dossary, M.A.; Abood, S.A.; Al-Saad, H.T. Factors affecting polycyclic aromatic hydrocarbon biodegradation by Aspergillus flavus. Remediation 2020, 30, 17–25. [Google Scholar] [CrossRef]
- Agrawal, N.; Barapatre, A.; Shahi, M.P.; Shahi, S.K. Biodegradation pathway of polycyclic aromatic hydrocarbons by ligninolytic fungus Podoscyphaelegans strain FTG4 and phytotoxicity evaluation of their metabolites. Environ. Process. 2021, 8, 1307–1335. [Google Scholar] [CrossRef]
- Chen, S.H.; Cheow, Y.L.; Ng, S.L.; Ting, A.S.Y. Mechanisms for metal removal established via electron microscopy and spectroscopy: A case study on metal tolerant fungi Penicilliumsimplicissimum. J. Hazard. Mater. 2019, 362, 394–402. [Google Scholar] [CrossRef]
- Zhang, D.; Yin, C.; Abbas, N.; Mao, Z.; Zhang, Y. Multiple heavy metal tolerance and removal by an earthworm gut fungus Trichodermabrevicompactum QYCD-6. Sci Rep. 2020, 10, 6940. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhuang, L.; Yuan, Y.; Tong, L.; Tsang, D.C. Enhancement of phenanthrene adsorption on a clayey soil and clay minerals by coexisting lead or cadmium. Chemosphere 2011, 83, 302–310. [Google Scholar] [CrossRef]
- Chen, S.; Yin, H.; Ye, J.; Peng, H.; Liu, Z.; Dang, Z.; Chang, J. Influence of co-existed benzo [a] pyrene and copper on the cellular characteristics of Stenotrophomonasmaltophilia during biodegradation and transformation. Bioresour. Technol. 2014, 158, 181–187. [Google Scholar] [CrossRef]
- Ma, X.K.; Ding, N.; Peterson, E.C.; Daugulis, A.J. Heavy metals species affect fungal-bacterial synergism during the bioremediation of fluoranthene. Appl. Microbiol. Biotechnol. 2016, 100, 7741–7750. [Google Scholar] [CrossRef]
- Sandrin, T.R.; Maier, R.M. Impact of metals on the biodegradation of organic pollutants. Environ. Health Perspect. 2003, 111, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.Y.; Liu, Y.; Wen, X.; Chen, S.F. Effects of algicide on the growth of Microcystisflos-aquae and adsorption capacity to heavy metals. Int. J. Environ. Sci. Technol. 2015, 12, 2339–2348. [Google Scholar] [CrossRef] [Green Version]
- Kuang, D.; Zhang, W.; Deng, Q.; Zhang, X.; Huang, K.; Guan, L.; Hu, D.; Tangchun, W.; Guo, H. Dose-response relationships of polycyclic aromatic hydrocarbons exposure and oxidative damage to DNA and lipid in coke oven workers. Environ. Sci. Technol. 2013, 47, 7446–7456. [Google Scholar] [CrossRef] [PubMed]
- Hkiri, N.; Aounallah, F.; Fouzai, K.; Chouchani, C.; Asses, N. Ability of marine-derived fungi isolated from polluted saline environment for enzymatic hydrocarbon remediation. Braz. J. Microbiol. Submitted (BJMI-D-22-01135).
- Darwish, A.M.G.; Abdel-Azeem, A.M. Chaetomium Enzymes and Their Applications. In Recent Developments on Genus Chaetomium; Springer: Cham, Switzerland, 2020; pp. 241–249. [Google Scholar] [CrossRef]
- Wang, J.; Shao, S.; Liu, C.; Song, Z.; Liu, S.; Wu, S. The genus Paraconiothyrium: Species concepts, biological functions, and secondary metabolites. Crit. Rev. Microbiol. 2021, 47, 781–810. [Google Scholar] [CrossRef]
- Debnath, R.; Mistry, P.; Roy, P.; Roy, B.; Saha, T. Partial purification and characterization of a thermophilic and alkali-stable laccase of Phomaherbarum isolate KU4 with dye-decolorization efficiency. Prep. Biochem. Biotechnol. 2021, 51, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.H.; Kim, D.H.; Park, J.H.; Park, Y.U.; Lee, M.Y.; Choi, M.H.; Lee, D.H.; Lee, J.K. Identification, enzymatic activity, and decay ability of basidiomycetous fungi isolated from the decayed bark of Mongolian oak (Quercus mongolica Fisch. Ex Ledeb.). J. For. Environ. Sci. 2021, 37, 52–61. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1988; Volume 161, pp. 238–249. [Google Scholar]
- Giardina, P.; Palmieri, G.; Fontanella, B.; Rivieccio, V.; Sannia, G. Manganese peroxidase isoenzymes produced by Pleurotus ostreatus grown on wood sawdust. Arch. Biochem. Biophys. 2000, 376, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, R.; Nüske, J.; Scheibner, K.; Spantzel, J.; Hofrichter, M. Novel haloperoxidase from the agaric basidiomycete Agrocybeaegerita oxidizes aryl alcohols and aldehydes. Appl. Environ. Microbiol. 2004, 70, 4575–4581. [Google Scholar] [CrossRef] [Green Version]
- Tien, M.; Kirk, T.K. Lignin-degrading enzyme from Phanerochaete chrysosporium: Purification, characterization, and catalytic properties of a unique H2O2-requiring oxygenase. Proc. Natl. Acad. Sci. USA 1984, 81, 2280–2284. [Google Scholar] [CrossRef] [Green Version]
- Novotný, Č.; Erbanová, P.; Šašek, V.; Kubátová, A.; Cajthaml, T.; Lang, E.; Krahl, J.; Zadražil, F. Extracellular oxidative enzyme production and PAH removal in soil by exploratory mycelium of white rot fungi. Biodegradation 1999, 10, 159–168. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Aranda, E.; Kinne, M.; Kluge, M.; Ullrich, R.; Hofrichter, M. Conversion of dibenzothiophene by the mushrooms Agrocybe aegerita and Coprinellus radians and their extracellular peroxygenases. Appl. Microbiol. Biotechnol. 2009, 82, 1057–1066. [Google Scholar] [CrossRef]
- Zucconi, J.M.; Moreels, G.; Parisot, J.P. Photolytic effects of solar radiation at stratospheric levels during sunrise and sunset. Can. J. Phys. 1981, 59, 1158–1169. [Google Scholar] [CrossRef]
- Onorati, F.; Mecozzi, M. Effects of two diluents in the Microtox toxicity bioassay with marine sediments. Chemosphere 2004, 54, 679–687. [Google Scholar] [CrossRef]
- Purswani, J.; Guisado, I.M.; Coello-Cabezas, J.; González-López, J.; Pozo, C. Social microbial inocula confer functional stability in a methyl tert-butyl ether extractive membrane biofilm bioreactor. Environ. Pollut. 2019, 244, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, J.; Gadd, G.M. Fungal bioremediation of soil co-contaminated with petroleum hydrocarbons and toxic metals. Appl. Microbiol. Biotechnol. 2020, 104, 8999–9008. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.; Malik, A.; Mishra, A.; Singh, D.K.; von Bergen, M.; Jehmlich, N. Mechanistic insight to mycoremediation potential of a metal resistant fungal strain for removal of hazardous metals from multimetal pesticide matrix. Environ. Pollut. 2020, 262, 114255. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Tsang, D.C.W. Redox-induced transformation of potentially toxic elements with organic carbon in soil. Carbon Res. 2022, 1, 9. [Google Scholar] [CrossRef]
- Liu, S.-H.; Zeng, G.-M.; Niu, Q.-Y.; Liu, Y.; Zhou, L.; Jiang, L.-H.; Tan, X.-F.; Xu, P.; Zhang, C.; Cheng, M. Bioremediation mechanisms of combined pollution of PAHs and heavy metals by bacteria and fungi: A mini review. Bioresour. Technol. 2017, 224, 25–33. [Google Scholar] [CrossRef]
- Li, X.; Pan, Y.; Hu, S.; Cheng, Y.; Wang, Y.; Wu, K.; Zhang, S.; Yang, S. Diversity of phenanthrene and benz [a] anthracene metabolic pathways in white rot fungus Pycnoporus sanguineus 14. Int. Biodeterior. Biodegrad. 2018, 134, 25–30. [Google Scholar] [CrossRef]
- Hong, J.W.; Park, J.Y.; Gadd, G.M. Pyrene degradation and copper and zinc uptake by Fusarium solani and Hypocrea lixii isolated from petrol station soil. J. Appl. Microbiol. 2010, 108, 2030–2040. [Google Scholar] [CrossRef]
- Ye, J.S.; Yin, H.; Qiang, J.; Peng, H.; Qin, H.M.; Zhang, N.; He, B.Y. Biodegradation of anthracene by Aspergillus fumigatus. J. Hazard. Mater. 2011, 185, 174–181. [Google Scholar] [CrossRef]
- Chen, A.; Zeng, G.; Chen, G.; Fan, J.; Zou, Z.; Li, H.; Hu, X.; Long, F. Simultaneous cadmium removal and 2, 4-dichlorophenol degradation from aqueous solutions by Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 2011, 91, 811–821. [Google Scholar] [CrossRef]
- Palmieri, G.; Giardina, P.; Bianco, C.; Fontanella, B.; Sannia, G. Copper induction of laccase isoenzymes in the ligninolytic fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 2000, 66, 920–924. [Google Scholar] [CrossRef] [Green Version]
- Luo, A.; Wu, Y.R.; Xu, Y.; Kan, J.; Qiao, J.; Liang, L.; Huang, T.; Hu, Z. Characterization of a cytochrome P450 monooxygenase capable of high molecular weight PAHs oxidization from Rhodococcus sp. P14. Process. Biochem. 2016, 51, 2127–2133. [Google Scholar] [CrossRef]
- Hadibarata, T.; Kristanti, R.A. Biodegradation and metabolite transformation of pyrene by basidiomycetes fungal isolate Armillaria sp. F022. Bioprocess. Biosyst. Eng. 2013, 36, 461–468. [Google Scholar] [CrossRef]
- Khudhair, A.B.; Hadibarata, T.; Yusoff, A.R.M.; Teh, Z.C.; Adnan, L.A.; Kamyab, H. Pyrene metabolism by new species isolated from soil Rhizoctonia zeae SOL3. Water Air Soil Pollut. 2015, 226, 186. [Google Scholar] [CrossRef]
- Agrawal, N.; Shahi, S.K. Degradation of polycyclic aromatic hydrocarbon (pyrene) using novel fungal strain Coriolopsisbyrsina strain APC5. Int. Biodeterior. Biodegrad. 2017, 122, 69–81. [Google Scholar] [CrossRef]
- Chen, M.; Ding, S.; Gao, S.; Fu, Z.; Tang, W.; Wu, Y.; Gong, M.; Wang, D.; Wang, Y. Efficacy of dredging engineering as a means to remove heavy metals from lake sediments. Sci. Total Environ. 2019, 665, 181–190. [Google Scholar] [CrossRef]
- Igiri, B.E.; Okoduwa, S.I.; Idoko, G.O.; Akabuogu, E.P.; Adeyi, A.O.; Ejiogu, I.K. Toxicity and bioremediation of heavy metals contaminated ecosystem from tannery wastewater: A review. J. Toxicol. 2018, 16, 2568038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, N.N.N.A.; Shahadat, M.; Omar, F.M.; Chew, A.W.; Kadir, M.O.A. Dry Trichoderma biomass: Biosorption behavior for the treatment of toxic heavy metal ions. Desalin. Water Treat. 2016, 57, 13106–13112. [Google Scholar] [CrossRef]
- Gola, D.; Malik, A.; Namburath, M.; Ahammad, S.Z. Removal of industrial dyes and heavy metals by Beauveria bassiana: FTIR, SEM, TEM and AFM investigations with Pb (II). Environ. Sci. Pollut. Res. 2018, 25, 20486–20496. [Google Scholar] [CrossRef] [PubMed]
- Pakshirajan, K.; Swaminathan, T. Biosorption of copper and cadmium in packed bed columns with live immobilized fungal biomass of Phanerochaete chrysosporium. Appl. Biochem. Biotechnol. 2019, 157, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Garcia, M.; López, M.J.; Suárez-Estrella, F.; Moreno, J. Compost as a source of microbial isolates for the bioremediation of heavy metals: In vitro selection. Sci. Total Environ. 2012, 431, 62–67. [Google Scholar] [CrossRef]
- Comte, S.; Guibaud, G.; Baudu, M. Biosorption properties of extracellular polymeric substances (EPS) towards Cd, Cu and Pb for different pH values. J. Hazard. Mater. 2008, 151, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Pinel-Cabello, M.; Jroundi, F.; López-Fernández, M.; Geffers, R.; Jarek, M.; Jauregui, R.; Link, A.; Vílchez-Vargas, R.; Merroun, M.L. Multisystem combined uranium resistance mechanisms and bioremediation potential of Stenotrophomonas bentonitica BII-R7: Transcriptomics and microscopic study. J. Hazard. Mater. 2021, 403, 123858. [Google Scholar] [CrossRef] [PubMed]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: Physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7, e07609. [Google Scholar] [CrossRef] [PubMed]






| MnP | UPO | LiP | Lac | ||||||
|---|---|---|---|---|---|---|---|---|---|
| T9 | T18 | T9 | T18 | T9 | T18 | T9 | T18 | ||
| C. jodhpurense | PAH | 87.6 ± 7.4 | 16.0 ± 1.4 | n.d. | 41.5 ± 6.4 | 32.4 ± 10.3 | 25.7 ± 1.8 | n.d. | 8.8 ± 2.5 |
| PAH + HM + ABT | 8 ± 2 | n.d. | n.d. | 8.98 ± 1.94 | 16.6 ± 3.9 | 43.2 ± 9.2 | n.d. | 8.4 ± 0.9 | |
| PAH + HM | n.d. | n.d. | n.d. | 12.2 ± 2.8 | 23.9 ± 3.8 | 99.4 ± 3.0 | n.d. | 8.0±1.3 | |
| C. maderasense | PAH | 274.0 ± 12.3 | 9.1 ± 4.5 | 15.2 ± 1.4 | 17.8 ± 2.3 | n.d. | n.d. | 43.7 ± 5.1 | 225.1 ± 13.8 |
| PAH + HM + ABT | 259 ± 12 | 9.0 ± 1.7 | 26.0 ±0.5 | n.d. | 10.2 ± 2.3 | n.d. | n.d. | n.d. | |
| PAH + HM | 120.6 ± 5.7 | 3.6 ± 0.7 | 175.7 ± 36.2 | n.d. | 10.9 ± 2.4 | n.d. | n.d. | 122.1 ± 3.5 | |
| PAH | 14.9 ± 0.9 | 85 ± 14 | 55.5 ± 5.6 | 37.9 ± 6.1 | 309.1 ± 41.2 | n.d. | n.d. | n.d. | |
| P. variabile | PAH + HM + ABT | 88.7 ± 3.9 | 66.1 ± 6.2 | 77.2 ± 4.8 | n.d. | 111 ± 31 | n.d. | n.d. | n.d. |
| PAH + HM | 51 ± 11 | 7.4 ± 2.8 | 200.4 ± 37.6 | 181.4 ± 17.0 | 329.4 ± 15.9 | n.d. | n.d. | n.d. | |
| E. lacerata | PAH | 25.0 ± 3.7 | n.d. | n.d. | 41.8 ± 12.1 | 62.2 ± 4.6 | 42.6 ± 2.7 | 29.3 ± 1.7 | n.d. |
| PAH + HM + ABT | n.d. | 66.5 ± 26.1 | n.d. | 105.7 ± 7.3 | 66.2 ± 5.7 | 46.6 ± 5.5 | n.d. | n.d. | |
| PAH + HM | n.d. | 15.6 ± 2.3 | n.d. | 71.4 ± 13.8 | 183.9 ± 25.4 | 62.9 ± 6.7 | n.d. | 0 | |
| P. betae | PAH | 4.2 ± 0.9 | 25.4±8.4 | 21.6 ± 2.4 | 65.0 ± 11.4 | 19.5 ± 2.0 | 69.4 ± 4.7 | 47.3 ± 3.9 | 63.5 ± 6.5 |
| PAH + HM + ABT | 24.7 ± 3.2 | 25.9 ± 3.4 | 22.4 ± 2.4 | 10.5 ± 1.3 | 9.1 ± 2.3 | 77.4 ± 10.5 | 65.6 ± 11.8 | 39.1 ± 0.9 | |
| PAH + HM | 18.5 ± 2.7 | n.d. | 27.3 ± 3.5 | 45.5 ± 10.2 | 32.8 ± 6.9 | 76.0 ± 11.9 | 687.5 ± 28.3 | 523.4 ± 21.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hkiri, N.; Olicón-Hernández, D.R.; Pozo, C.; Chouchani, C.; Asses, N.; Aranda, E. Simultaneous Heavy Metal-Polycyclic Aromatic Hydrocarbon Removal by Native Tunisian Fungal Species. J. Fungi 2023, 9, 299. https://doi.org/10.3390/jof9030299
Hkiri N, Olicón-Hernández DR, Pozo C, Chouchani C, Asses N, Aranda E. Simultaneous Heavy Metal-Polycyclic Aromatic Hydrocarbon Removal by Native Tunisian Fungal Species. Journal of Fungi. 2023; 9(3):299. https://doi.org/10.3390/jof9030299
Chicago/Turabian StyleHkiri, Neila, Dario R. Olicón-Hernández, Clementina Pozo, Chedly Chouchani, Nedra Asses, and Elisabet Aranda. 2023. "Simultaneous Heavy Metal-Polycyclic Aromatic Hydrocarbon Removal by Native Tunisian Fungal Species" Journal of Fungi 9, no. 3: 299. https://doi.org/10.3390/jof9030299