
Global Molecular Response of Paracoccidioides brasiliensis to Zinc Deprivation: Analyses at Transcript, Protein and MicroRNA Levels
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Growth Conditions
2.2. RNA Extraction and cDNA Sequencing
2.3. Identification of MicroRNAs
2.4. Analysis of the Expression and Prediction of Target Genes of P. brasiliensis MicroRNAs
2.5. Preparation of Protein Extracts
2.6. Proteomic Analysis by Liquid Chromatography Coupled to NanoUPLC-MSE Mass Spectrometry, Data Processing and Protein Identification
2.7. Proteomic Data Analysis
2.8. mRNA-Seq Analysis
2.9. Statistical Analysis
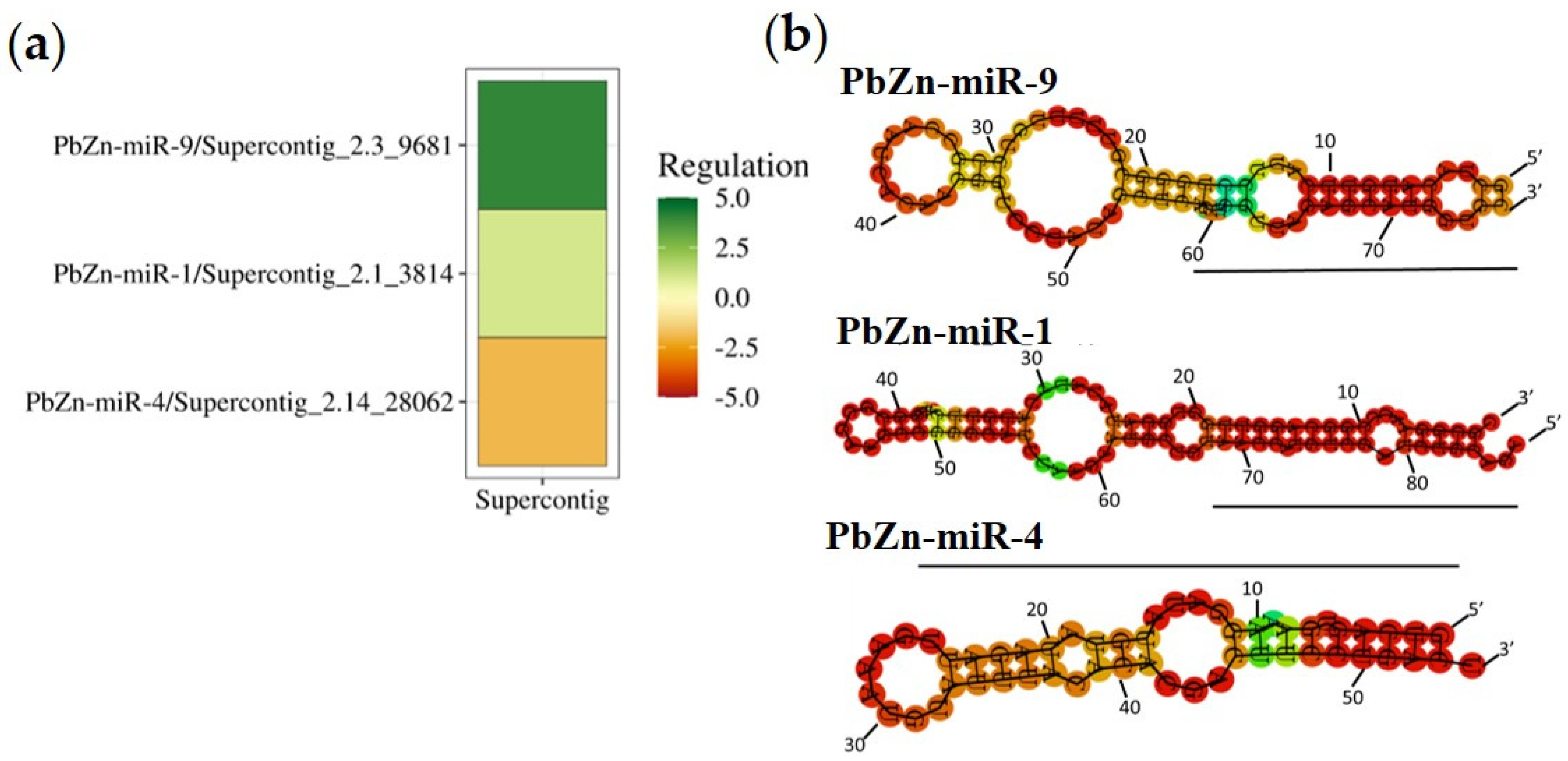
2.10. Construction of Heat Maps and MicroRNAs Images
3. Results
3.1. Viability of Cells Exposed to Zinc Deprivation
3.2. Identification of MiRNAs
3.3. Some Prominent RNAs Putatively Regulated by Micro RNAs during Zinc Deprivation
3.4. Transcriptional and Proteomic Analysis Support the Regulation by MicroRNAs
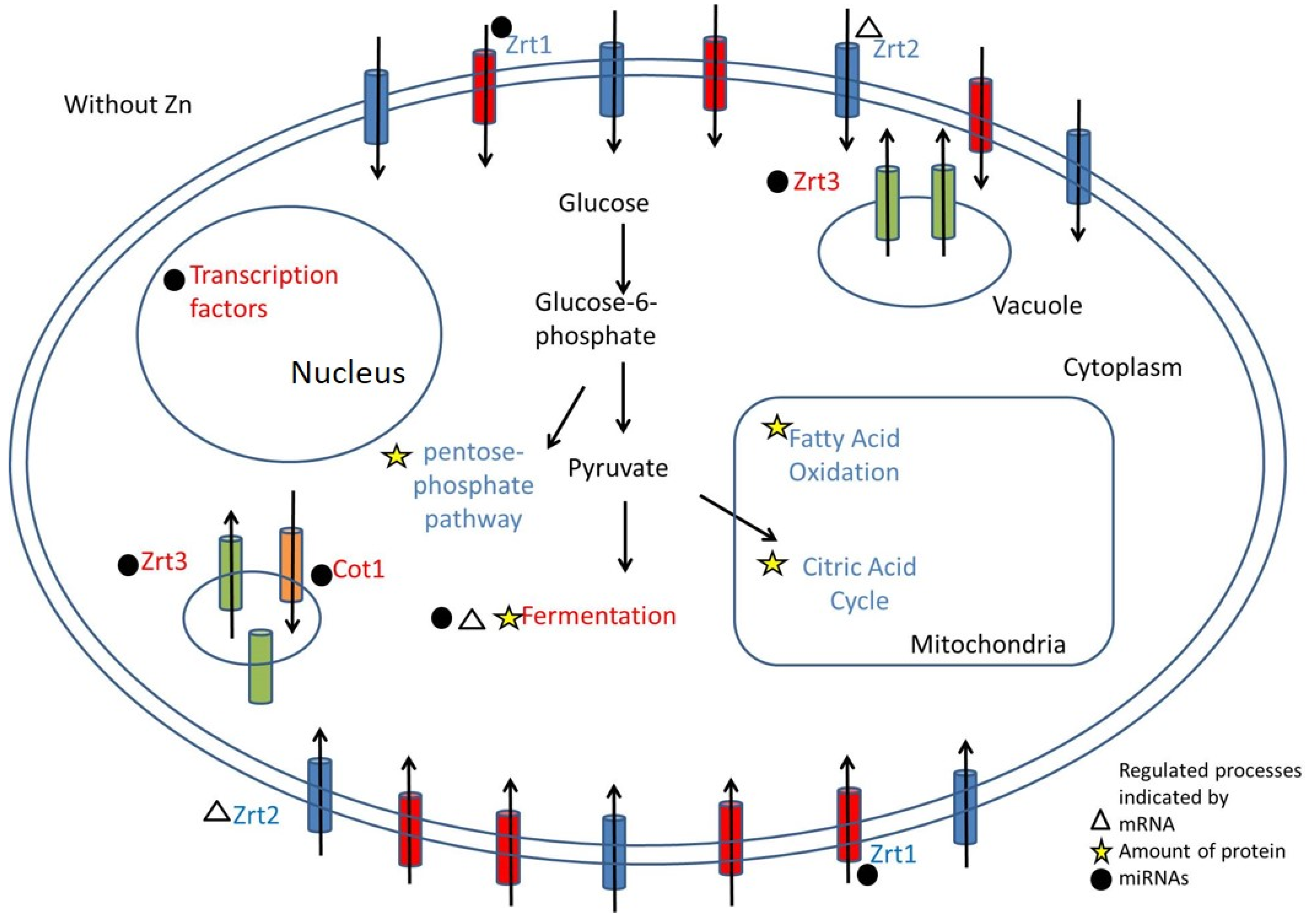
3.5. Metabolic Changes Indicated by Proteome and Transcriptome Analysis
3.6. Integrative Molecular Mechanisms that Promote Fungal Adaptation to Low Zinc
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal Ions in Biological Catalysis: From Enzyme Databases to General Principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc Coordination Environments in Proteins as Redox Sensors and Signal Transducers. Antioxid. Redox Signal. 2006, 8, 1419–1441. [Google Scholar] [CrossRef] [PubMed]
- Hambidge, K.M.; Krebs, N.F. Zinc Deficiency: A Special Challenge. J. Nutr. 2007, 137, 1101–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, L.M.; Rink, L.; Wessels, I. Increase of the Intracellular Zinc Concentration Leads to an Activation and Internalisation of the Epidermal Growth Factor Receptor in A549 Cells. Int. J. Mol. Sci. 2020, 22, 326. [Google Scholar] [CrossRef]
- Stefanidou, M.; Maravelias, C.; Dona, A.; Spiliopoulou, C. Zinc: A Multipurpose Trace Element. Arch. Toxicol. 2005, 80, 1–9. [Google Scholar] [CrossRef]
- Gitan, R.S.; Eide, D.J. Zinc-Regulated Ubiquitin Conjugation Signals Endocytosis of the Yeast ZRT1 Zinc Transporter. Biochem. J. 2000, 346, 329–336. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Cao, C.L.; Liu, Y.L.; Wang, J.; Li, J.; Li, S.Y.; Deng, Y. Identification of the Genetic Requirements for Zinc Tolerance and Toxicity in Saccharomyces Cerevisiae. G3 Genes Genomes Genet. 2020, 10, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Lulloff, S.J.; Hahn, B.L.; Sohnle, P.G. Fungal Susceptibility to Zinc Deprivation. J. Lab. Clin. Med. 2004, 144, 208–214. [Google Scholar] [CrossRef]
- Ding, C.; Hu, G.; Jung, W.H.; Kronstad, J.W. Essential Metals in Cryptococcus Neoformans: Acquisition and Regulation. Curr. Fungal Infect. Rep. 2014, 8, 153–162. [Google Scholar] [CrossRef]
- Subramanian Vignesh, K.; Deepe, G.S. Immunological Orchestration of Zinc Homeostasis: The Battle between Host Mechanisms and Pathogen Defenses. Arch. Biochem. Biophys. 2016, 611, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Hamer, D.H. Metallothionein. Annu. Rev. Biochem. 1986, 55, 913–951. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D. Cloning and Functional Characterization of a Mammalian Zinc Transporter That Confers Resistance to Zinc. EMBO J. 1995, 14, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Zinc Transporters and the Cellular Trafficking of Zinc. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2006, 1763, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, L.W.; Bailão, A.M.; Soares, C.M.D.A.; Bailão, M.G.S. Zinc at the Host–Fungus Interface: How to Uptake the Metal? J. Fungi 2020, 6, 305. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Eide, D. The Yeast ZRT1 Gene Encodes the Zinc Transporter Protein of a High-Affinity Uptake System Induced by Zinc Limitation. Proc. Natl. Acad. Sci. USA 1996, 93, 2454–2458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Eide, D. The Gene Encodes the Low Affinity Zinc Transporter In. J. Biol. Chem. 1996, 271, 23203–23210. [Google Scholar] [CrossRef] [Green Version]
- MacDiarmid, C.W.; Gaither, L.A.; Eide, D. Zinc Transporters That Regulate Vacuolar Zinc Storage in Saccharomyces Cerevisiae. EMBO J. 2000, 19, 2845–2855. [Google Scholar] [CrossRef] [Green Version]
- Do, E.; Hu, G.; Caza, M.; Kronstad, J.; Jung, W.H. The ZIP Family Zinc Transporters Support the Virulence of Cryptococcus Neoformans. Med. Mycol. 2016, 54, 605–615. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Schneider, R.; Diehl, C.; dos Santos, F.M.; Piffer, A.C.; Garcia, A.W.A.; Kulmann, M.I.R.; Schrank, A.; Kmetzsch, L.; Vainstein, M.H.; Staats, C.C. Effects of Zinc Transporters on Cryptococcus Gattii Virulence. Sci. Rep. 2015, 5, 10104. [Google Scholar] [CrossRef] [Green Version]
- Amich, J.; Vicentefranqueira, R.; Mellado, E.; Ruiz-Carmuega, A.; Leal, F.; Calera, J.A. The ZrfC Alkaline Zinc Transporter Is Required for Aspergillus Fumigatus Virulence and Its Growth in the Presence of the Zn/Mn-Chelating Protein Calprotectin. Cell. Microbiol. 2014, 16, 548–564. [Google Scholar] [CrossRef]
- Crawford, A.C.; Lehtovirta-Morley, L.E.; Alamir, O.; Niemiec, M.J.; Alawfi, B.; Alsarraf, M.; Skrahina, V.; Costa, A.C.B.P.; Anderson, A.; Yellagunda, S.; et al. Biphasic Zinc Compartmentalisation in a Human Fungal Pathogen. PLoS Pathog. 2018, 14, e1007013. [Google Scholar] [CrossRef] [Green Version]
- Citiulo, F.; Jacobsen, I.D.; Miramón, P.; Schild, L.; Brunke, S.; Zipfel, P.; Brock, M.; Hube, B.; Wilson, D. Candida Albicans Scavenges Host Zinc via Pra1 during Endothelial Invasion. PLoS Pathog. 2012, 8, e1002777. [Google Scholar] [CrossRef] [Green Version]
- Kamizono, A.; Nishizawa, M.; Teranishi, Y.; Murata, K.; Kimura, A. Identification of a Gene Conferring Resistance to Zinc and Cadmium Ions in the Yeast Saccharomyces Cerevisiae. Mol. Gen. Genet. MGG 1989, 219, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Conklin, D.S.; Mcmaster, J.A.; Culbertson, M.R.; Kung, C. COT1, a Gene Involved in Cobalt Accumulation in Saccharomyces Cerevisiae. Mol. Cell. Biol. 1992, 12, 3678–3688. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Frey, A.G.; Eide, D.J. Transcriptional Regulation of the Zrg17 Zinc Transporter of the Yeast Secretory Pathway. Biochem. J. 2011, 435, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, A.J.; Zhao, H.; Luo, H.; Jensen, L.T.; Srinivasan, C.; Evans-Galea, M.; Winge, D.R.; Eide, D.J. A Dual Role for Zinc Fingers in Both DNA Binding and Zinc Sensing by the Zap1 Transcriptional Activator. EMBO J. 2000, 19, 3704–3713. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.Y.; Bird, A.J.; Chung, L.M.; Newton, M.A.; Winge, D.R.; Eide, D.J. Differential Control of Zap1-Regulated Genes in Response to Zinc Deficiency in Saccharomyces Cerevisiae. BMC Genom. 2008, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Vicentefranqueira, R.; Amich, J.; Marín, L.; Sánchez, C.I.; Leal, F.; Calera, J.A. The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus Fumigatus. Genes 2018, 9, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, M.A. The Ecology of Paracoccidioides Brasiliensis: A Puzzle Still Unsolved. Med. Mycol. 1985, 23, 323–334. [Google Scholar] [CrossRef]
- Segrelles-Calvo, G.; De, G.R.; Ará Ujo, S.; Frases, S.; Jim’, J.; Diaz, J. Systemic Mycoses: A Potential Alert for Complications in COVID-19 Patients. Future Microbiol. 2020, 15, 1405–1413. [Google Scholar] [CrossRef]
- Song, G.; Liang, G.; Liu, W. Fungal Co-Infections Associated with Global COVID-19 Pandemic: A Clinical and Diagnostic Perspective from China. Mycopathologia 2020, 185, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Bailão, A.M.; Schrank, A.; Borges, C.L.; Dutra, V.; Walquíria Inês Molinari-Madlum, E.E.; Soares Felipe, M.S.; Soares Mendes-Giannini, M.J.; Martins, W.S.; Pereira, M.; Maria de Almeida Soares, C. Differential Gene Expression by Paracoccidioides Brasiliensis in Host Interaction Conditions: Representational Difference Analysis Identifies Candidate Genes Associated with Fungal Pathogenesis. Microbes Infect. 2006, 8, 2686–2697. [Google Scholar] [CrossRef] [PubMed]
- Bailão, A.M.; Shrank, A.; Borges, C.L.; Parente, J.A.; Dutra, V.; Felipe, M.S.S.; Fiúza, R.B.; Pereira, M.; Soares, C.M.D.A.; Soares, A. The Transcriptional Profile of Paracoccidioides Brasiliensis Yeast Cells Is Influenced by Human Plasma. FEMS Immunol. Med. Microbiol. 2007, 51, 43–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.G.; Schrank, A.; Bailão, E.F.L.C.; Bailão, A.M.; Borges, C.L.; Staats, C.C.; Parente, J.A.; Pereira, M.; Salem-Izacc, S.M.; Mendes-Giannini, M.J.S.; et al. The Homeostasis of Iron, Copper, and Zinc in Paracoccidioides Brasiliensis, Cryptococcus Neoformans Var. Grubii, and Cryptococcus Gattii: A Comparative Analysis. Front. Microbiol. 2011, 2, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parente, A.F.A.; de Rezende, T.C.V.; de Castro, K.P.; Bailão, A.M.; Parente, J.A.; Borges, C.L.; Silva, L.P.; de Almeida Soares, C.M. A Proteomic View of the Response of Paracoccidioides Yeast Cells to Zinc Deprivation. Fungal Biol. 2013, 117, 399–410. [Google Scholar] [CrossRef]
- de Curcio, J.S.; Silva, M.G.; Silva Bailão, M.G.; Báo, S.N.; Casaletti, L.; Bailão, A.M.; de Almeida Soares, C.M. Identification of Membrane Proteome of Paracoccidioides Lutzii and Its Regulation by Zinc. Future Sci. OA 2017, 3, FSO232. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian MicroRNA Host Genes and Transcription Units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Schnitger, A.K.D.; Machova, A.; Mueller, R.U.; Androulidaki, A.; Schermer, B.; Pasparakis, M.; Krönke, M.; Papadopoulou, N. Listeria Monocytogenes Infection in Macrophages Induces Vacuolar-Dependent Host MiRNA Response. PLoS ONE 2011, 6, e27435. [Google Scholar] [CrossRef]
- Modesto, I.; Inácio, V.; van de Peer, Y.; Miguel, C.M. MicroRNA-Mediated Post-Transcriptional Regulation of Pinus Pinaster Response and Resistance to Pinewood Nematode. Sci. Rep. 2022, 12, 5160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton Plants Export MicroRNAs to Inhibit Virulence Gene Expression in a Fungal Pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- de Curcio, J.S.; Paccez, J.D.; Novaes, E.; Brock, M.; de Almeida Soares, C.M. Cell Wall Synthesis, Development of Hyphae and Metabolic Pathways Are Processes Potentially Regulated by MicroRNAs Produced between the Morphological Stages of Paracoccidioides Brasiliensis. Front. Microbiol. 2018, 9, 3057. [Google Scholar] [CrossRef]
- de Curcio, J.S.; Oliveira, L.N.; Batista, M.P.; Novaes, E.; de Almeida Soares, C.M. MiRNAs Regulate Iron Homeostasis in Paracoccidioides Brasiliensis. Microbes Infect. 2021, 23, 104772. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Jimenez, B.E. Growth of Paracoccidioides Brasiliensis Yeast Phase in a Chemically Defined Culture Medium. J. Clin. Microbiol. 1980, 12, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, A.A.; Neves, B.J.; do Carmo Silva, L.; de Almeida Soares, C.M.; Andrade, C.H.; Pereira, M. Drug Repurposing for Paracoccidioidomycosis through a Computational Chemogenomics Framework. Front. Microbiol. 2019, 10, 1301. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; MacKowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. MiRDeep2 Accurately Identifies Known and Hundreds of Novel MicroRNA Genes in Seven Animal Clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Rehmsmeier, M.; Steffen, P.; Höchsmann, M.; Giegerich, R. Fast and Effective Prediction of MicroRNA/Target Duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Murad, A.M.; Souza, G.H.M.F.; Garcia, J.S.; Rech, E.L. Detection and Expression Analysis of Recombinant Proteins in Plant-Derived Complex Mixtures Using NanoUPLC-MSE. J. Sep. Sci. 2011, 34, 2618–2630. [Google Scholar] [CrossRef] [Green Version]
- Baeza, L.C.; da Mata, F.R.; Pigosso, L.L.; Pereira, M.; de Souza, G.H.M.F.; Coelho, A.S.G.; de Almeida Soares, C.M. Differential Metabolism of a Two-Carbon Substrate by Members of the Paracoccidioides Genus. Front. Microbiol. 2017, 8, 2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geromanos, S.J.; Vissers, J.P.C.; Silva, J.C.; Dorschel, C.A.; Li, G.Z.; Gorenstein, M.V.; Bateman, R.H.; Langridge, J.I. The Detection, Correlation, and Comparison of Peptide Precursor and Product Ions from Data Independent LC-MS with Data Dependant LC-MS/MS. Proteomics 2009, 9, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Priebe, S.; Kreisel, C.; Horn, F.; Guthke, R.; Linde, J. FungiFun2: A Comprehensive Online Resource for Systematic Analysis of Gene Lists from Fungal Species. Bioinformatics 2015, 31, 445–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putignano, V.; Rosato, A.; Banci, L.; Andreini, C. MetalPDB in 2018: A Database of Metal Sites in Biological Macromolecular Structures. Nucleic Acids Res. 2018, 46, D459–D464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- de Curcio, J.S.; Batista, M.P.; Paccez, J.D.; Novaes, E.; de Almeida Soares, C.M. In Silico Characterization of MicroRNAs-like Sequences in the Genome of Paracoccidioides Brasiliensis. Genet. Mol. Biol. 2019, 42, 95–107. [Google Scholar] [CrossRef]
- Hettema, E.H.; Valdez-Taubas, J.; Pelham, H.R.B. Bsd2 Binds the Ubiquitin Ligase Rsp5 and Mediates the Ubiquitination of Transmembrane Proteins. EMBO J. 2004, 23, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Portnoy, M.E.; Liu, X.F.; Culotta, V.C. Saccharomyces Cerevisiae Expresses Three Functionally Distinct Homologues of the Nramp Family of Metal Transporters. Mol. Cell. Biol. 2000, 20, 7893–7902. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D. An Evolutionary Perspective on Zinc Uptake by Human Fungal Pathogens. Metallomics 2015, 7, 979–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamata, T.; Horie, T.; Matsunami, M.; Sasaki, M.; Ohsumi, Y. Zinc Starvation Induces Autophagy in Yeast. J. Biol. Chem. 2017, 292, 8520–8530. [Google Scholar] [CrossRef] [Green Version]
- Assunção, L.D.P.; Moraes, D.; Soares, L.W.; Silva-Bailão, M.G.; de Siqueira, J.G.; Baeza, L.C.; Báo, S.N.; Soares, C.M.D.A.; Bailão, A.M. Insights Into Histoplasma Capsulatum Behavior on Zinc Deprivation. Front. Cell. Infect. Microbiol. 2020, 10, 711. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Weisenhorn, E.; Macdiarmid, C.W.; Andreini, C.; Bucci, M.; Taggart, J.; Banci, L.; Russell, J.; Coon, J.J.; Eide, D.J. The Cellular Economy of the Saccharomyces Cerevisiae Zinc Proteome. Metallomics 2018, 10, 1755–1776. [Google Scholar] [CrossRef]
- Li, Q.; Abrashev, R.; Harvey, L.M.; McNeil, B. Oxidative Stress-Associated Impairment of Glucose and Ammonia Metabolism in the Filamentous Fungus, Aspergillus Niger B1-D. Mycol. Res. 2008, 112, 1049–1055. [Google Scholar] [CrossRef]
- Culotta, V.C.; Yang, M.; O’Halloran, T.V. Activation of Superoxide Dismutases: Putting the Metal to the Pedal. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2006, 1763, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Bird, A.J.; Gordon, M.; Eide, D.J.; Winge, D.R. Repression of ADH1 and ADH3 during Zinc Deficiency by Zap1-Induced Intergenic RNA Transcripts. EMBO J. 2006, 25, 5726. [Google Scholar] [CrossRef] [Green Version]
- Amini, S.; Jacobsen, A.; Ivanova, O.; Lijnzaad, P.; Heringa, J.; Holstege, F.C.P.; Feenstra, K.A.; Kemmeren, P. The Ability of Transcription Factors to Differentially Regulate Gene Expression Is a Crucial Component of the Mechanism Underlying Inversion, a Frequently Observed Genetic Interaction Pattern. PLoS Comput. Biol. 2019, 15, e1007061. [Google Scholar] [CrossRef] [Green Version]
- Nicola, A.M.; Andrade, R.V.; Dantas, A.S.; Andrade, P.A.; Arraes, F.B.M.; Fernandes, L.; Silva-Pereira, I.; Felipe, M.S.S. The Stress Responsive and Morphologically Regulated Hsp90 Gene from Paracoccidioides Brasiliensis Is Essential to Cell Viability. BMC Microbiol. 2008, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Tamayo, D.; Muñoz, J.F.; Torres, I.; Almeida, A.J.; Restrepo, A.; McEwen, J.G.; Hernández, O. Involvement of the 90 KDa Heat Shock Protein during Adaptation of Paracoccidioides Brasiliensis to Different Environmental Conditions. Fungal Genet. Biol. 2013, 51, 34–41. [Google Scholar] [CrossRef]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 Chaperone Machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Genest, O.; Wickner, S.; Doyle, S.M. Hsp90 and Hsp70 Chaperones: Collaborators in Protein Remodeling. J. Biol. Chem. 2019, 294, 2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.V.; Topper, S.E.; Hubler, S.L.; Hose, J.; Wenger, C.D.; Coon, J.J.; Gasch, A.P. A Dynamic Model of Proteome Changes Reveals New Roles for Transcript Alteration in Yeast. Mol. Syst. Biol. 2011, 7, 514. [Google Scholar] [CrossRef] [PubMed]
- Reimegård, J.; Tarbier, M.; Danielsson, M.; Schuster, J.; Baskaran, S.; Panagiotou, S.; Dahl, N.; Friedländer, M.R.; Gallant, C.J. A Combined Approach for Single-Cell MRNA and Intracellular Protein Expression Analysis. Commun. Biol. 2021, 4, 624. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, M.D.; Beynon, R.J.; Gethings, L.A.; Claydon, A.J.; Langridge, J.I.; Vissers, J.P.C.; Brown, A.J.P.; Hammond, D.E. Specificity of the Osmotic Stress Response in Candida Albicans Highlighted by Quantitative Proteomics. Sci. Rep. 2018, 8, 14492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enjalbert, B.; Smith, D.A.; Cornell, M.J.; Alam, I.; Nicholls, S.; Brown, A.J.P.; Quinn, J. Role of the Hog1 Stress-Activated Protein Kinase in the Global Transcriptional Response to Stress in the Fungal Pathogen Candida Albicans. Mol. Biol. Cell 2006, 17, 1018–1032. [Google Scholar] [CrossRef] [Green Version]
- Amich, J.; Leal, F.; Calera, J.A. Repression of the Acid ZrfA/ZrfB Zinc-Uptake System of Aspergillus Fumigatus Mediated by PacC under Neutral, Zinc-Limiting Conditions. Int. Microbiol. 2009, 12, 39–47. [Google Scholar] [CrossRef]
- Bird, A.J.; Blankman, E.; Stillman, D.J.; Eide, D.J.; Winge, D.R. The Zap1 Transcriptional Activator Also Acts as a Repressor by Binding Downstream of the TATA Box in ZRT2. EMBO J. 2004, 23, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Bailão, E.F.L.C.; Parente, A.F.A.; Parente, J.A.; Silva-Bailão, M.G.; de Castro, K.P.; Kmetzsch, L.; Staats, C.C.; Schrank, A.; Vainstein, M.H.; Borges, C.L.; et al. Metal Acquisition and Homeostasis in Fungi. Curr. Fungal Infect. Rep. 2012, 6, 257–266. [Google Scholar] [CrossRef]
- Aguilar, C.; Mano, M.; Eulalio, A. MicroRNAs at the Host-Bacteria Interface: Host Defense or Bacterial Offense. Trends Microbiol. 2019, 27, 206–218. [Google Scholar] [CrossRef]
- Ho, J.; Chan, H.; Wong, S.H.; Wang, M.H.T.; Yu, J.; Xiao, Z.; Liu, X.; Choi, G.; Leung, C.C.H.; Wong, W.T.; et al. The Involvement of Regulatory Non-Coding RNAs in Sepsis: A Systematic Review. Crit. Care 2016, 20, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amich, J.; Calera, J.A. Zinc Acquisition: A Key Aspect in Aspergillus Fumigatus Virulence. Mycopathologia 2014, 178, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Eide, D.J. Zap1p, a Metalloregulatory Protein Involved in Zinc-Responsive Transcriptional Regulation in Saccharomyces Cerevisiae. Mol. Cell. Biol. 1997, 17, 5044–5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Ferguson, J.; Chang, J.T.; Kluger, Y. Inter- and Intra-Combinatorial Regulation by Transcription Factors and MicroRNAs. BMC Genom. 2007, 8, 396. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Library a | Mature Sequence | Star Sequence | Precursor Sequence | Id b |
|---|---|---|---|---|
| C1, C2, C3, D1, D2, D3 | uaaccaugucgaucugcaga | cgcggaacccggcagguuggu | cgcggaacccggcagguuggugcauauauauauguucauuggcgggaagccgggcauuuaagaaugcuguaaccaugucgaucugcaga | PbZn-miR-1 * |
| C1, C3, D1, D2, D3 | uuauuuuuggaacuuuuu | uaguucggagucugggu | uaguucggagucuggguauggcuuucuuucuguaaugguuuuguuauguuauuuuauuuuuggaacuuuu | PbZn-miR-2 * |
| C1, C2, C3, D1, D2, D3 | uauaucaggguguguguuggc | caaccccugauugguagg | uauaucaggguguguguuggcagcaagauauugcaaagagccaaccccugauugguagg | PbZn-miR-3 * |
| C1, C2, C3, D1, D2, D3 | gucagugaaaggauauguauagauu | uuacacacgacuuccugacu | gucagugaaaggauauguauagauugaaauguauuuacacacgacuuccugacu | PbZn-miR-4 * |
| C1, C2, C3, D1, D2, D3 | uucuuuaugaacaguggcugg | agucauucauugagaauu | agucauucauugagaauuucucaagcagguucuuuaugaacaguggcugg | PbZn-miR-5 * |
| C1, C2, C3, D1, D2, D3 | ugccuuguagauaucuaaga | ucuuuucaguaggacaca | ucuuuucaguaggacacaucuauauuagaagugccuuguagauaucuaaga | PbZn-miR-6 |
| C1, C3, D1, D2, D3 | uaggucugagacagcuca | agcuguucagaccagu | uaggucugagacagcucacucagagagggcuucaaaaugcuaagaguggacaaucuucucuuauauagcuguucagaccagu | PbZn-miR-7 |
| C1, C2, C3, D1, D2, D3 | cuaguuagguuaguuaguua | gcuaacuaacuaacuaaau | gcuaacuaacuaacuaaauaacuaacuaacuaaccuaguuacuuaacuaguuagguuaguuaguua | PbZn-miR-8 * |
| C1, C2, C3, D1, D2, D3 | aguugguuagagcauggugc | gcuacauguucauuccugg | gcuacauguucauuccugggcguuuuuguccgccaaucacaacgguccuauagcucaguugguuagagcauggugc | PbZn-miR-9 |
| C1, C3, D1, D2 | cacccguggacugugccaugc | auuguacggagcacggguguc | cacccguggacugugccaugcaucuacgugcauuguacggagcacggguguc | PbZn-miR-10 |
| C1, C2, C3, D1, D2, D3 | uaguuagguuaguuaguua | acuaaguuaguuaacaguuaac | acuaaguuaguuaacaguuaacuaacuaguuauuuaguuaacuaguuaguuaguuagguuaguuaguua | PbZn-miR-11 * |
| C1, C2, D1, D2, D3 | aucuugacugucgaaaggg | cuagacagaucaugugaucu | cuagacagaucaugugaucuccauauccgcuggggaucuugacugucgaaaggg | PbZn-miR-12 |
| C1, C2, C3, D1, D2, D3 | ucagaugaugaaaaagaugcugaca | ucagacuggucucugcugaau | ucagacuggucucugcugaaucuccacugcggcaaauggaaguuucagaugaugaaaaagaugcugaca | PbZn-miR-13 * |
| C1, C2, C3, D1, D2, D3 | guggaugaugagagaacuucugagg | uuagcaagugccauuauccauga | guggaugaugagagaacuucugagggcugaacaggagguccuuuagcaagugccauuauccauga | PbZn-miR-14 |
| C1, C2, C3, D1, D2, D3 | aaugggcacuguuaacuaacuu | guugguuaacgguugguuacuaa | uugguuaacgguugguuacuaagaaugggcacuguuaacuaacuu | PbZn-miR-15 |
| C1, C3, D1, D2 | uacuuuuucgauugaggggacgu | gucguucuuaaucccgcg | uacuuuuucgauugaggggacguuccgaggaggagcgucguucuuaaucccgcg | PbZn-miR-16 |
| C1, C2, C3, D1, D2, D3 | ucggaagaugaugaacgagcgg | guucaacugcaucaucaucacc | guucaacugcaucaucaucacccccucuagaggcagggucggaagaugaugaacgagcgg | PbZn-miR-17 ** |
| C1, C2, C3, D1, D2, D3 | uaagacgcgaacuguuugaggu | uuauccaacgguucccauuugg | uaagacgcgaacuguuugagguuucguagaauuauccaacgguucccauuugg | PbZn-miR-18 * |
| C1, C3, D1, D2, D3 | uggaguggucgcccaggcug | gcccaacacccccacgcgcc | uggaguggucgcccaggcugcuagggcccaacaacccaauuagggcccaacagcccaacacccccacgcgcc | PbZn-miR-19 |
| MicroRNA | Superconting | Targets | MFE | Functional Classification | p-Value | Confirmed by | |
|---|---|---|---|---|---|---|---|
| PbZn-miR-1 | Supercontig_2.1_3814 | PADG_03627/2-oxoisovalerate dehydrogenase subunit beta | −23.6 kcal/mol | Amino acid metabolism | 0.033793 | 3′UTR | Proteome |
| PbZn-miR-1 | Supercontig_2.1_3814 | PADG_02134/Coatomer subunit epsilon | −24.1 kcal/mol | Cellular transport, transport facilities and transport routes | 0.027341 | 3′UTR | Proteome |
| PbZn-miR-1 | Supercontig_2.1_3814 | PADG_05239/Tubulin-specific chaperone A | −22.9 kcal/mol | Cytoskeleton/structural proteins | 0.045399 | 3′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_08119/Fumarate hydratase, mitochondrial | −23.5 kcal/mol | Tricarboxylic-acid pathway (citrate cycle, Krebs cycle, TCA cycle) | 0.035252 | 3′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_03403/aldehyde dehydrogenase | −25.0 kcal/mol | Cell rescue, defense and virulence | 0.018564 | 3′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_08409/hypothetical protein | −24.2 kcal/mol | Unclassified proteins | 0.026205 | 3′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_01174/alcohol dehydrogenase | −23.3 kcal/mol | Fermentation | 0.038359 | 5′UTR | Transcriptome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_04701/alcohol dehydrogenase | −25.4 kcal/mol | Fermentation | 0.015719 | 5′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_05798/ssDNA binding protein, putative | −22.9 kcal/mol | Cell cycle and DNA processing | 0.045208 | 5′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_04966/phosducin family protein | −23.8 kcal/mol | Protein fate (folding, modification, destination) | 0.031050 | 5′UTR | Proteome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_03258/lipid A export ATP-binding/permease protein msbA | −25.6 kcal/mol | Cellular transport, transport facilities and transport routes | 0.014432 | 5′UTR | Transcriptome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_03130/Myosin-10 | −23.8 kcal/mol | Biogenesis of cellular components | 0.031050 | 5′UTR | Transcriptome |
| PbZn-miR-9 | Supercontig_2.3_9681 | PADG_05949/ hypothetical protein | −23.2 kcal/mol | Unclassified proteins | 0.040012 | 5′UTR | Transcriptome |
| PbZn-miR-4 | Supercontig_2.14_28062 | PADG_00171/cytochrome b2 | −24.6 kcal/mol | Electron transport and membrane-associated energy conservation | 0.029065 | 3′UTR | Proteome |
| PbZn-miR-4 | Supercontig_2.14_28062 | PADG_00806/BTB domain-containing protein | −23.6 kcal/mol | Translation | 0.043859 | 5′UTR | Transcriptome |
| PbZn-miR-4 | Supercontig_2.14_28062 | PADG_07281/Guanine nucleotide-binding protein subunit alpha | −23.5 kcal/mol | Cellular communication/signal transduction mechanism | 0.045692 | 5′UTR | Proteome |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesquita, L.A.; Bailão, A.M.; de Curcio, J.S.; da Silva, K.L.P.; da Rocha Fernandes, G.; Silva-Bailão, M.G.; Novaes, E.; de Almeida Soares, C.M. Global Molecular Response of Paracoccidioides brasiliensis to Zinc Deprivation: Analyses at Transcript, Protein and MicroRNA Levels. J. Fungi 2023, 9, 281. https://doi.org/10.3390/jof9030281
Mesquita LA, Bailão AM, de Curcio JS, da Silva KLP, da Rocha Fernandes G, Silva-Bailão MG, Novaes E, de Almeida Soares CM. Global Molecular Response of Paracoccidioides brasiliensis to Zinc Deprivation: Analyses at Transcript, Protein and MicroRNA Levels. Journal of Fungi. 2023; 9(3):281. https://doi.org/10.3390/jof9030281
Chicago/Turabian StyleMesquita, Lorena Alves, Alexandre Melo Bailão, Juliana Santana de Curcio, Kassyo Lobato Potenciano da Silva, Gabriel da Rocha Fernandes, Mirelle Garcia Silva-Bailão, Evandro Novaes, and Célia Maria de Almeida Soares. 2023. "Global Molecular Response of Paracoccidioides brasiliensis to Zinc Deprivation: Analyses at Transcript, Protein and MicroRNA Levels" Journal of Fungi 9, no. 3: 281. https://doi.org/10.3390/jof9030281