
Metabolic Regulation of Two pksCT Gene Transcripts in Monascus ruber Impacts Citrinin Biosynthesis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Construction of the Fungal Expression Vector pC3300-neo
2.3. Plasmid Construction for Silencing pksCTα and pksCT (α + β) Genes
2.4. Agrobacterium Tumefaciens-Mediated Transformation (ATMT)
2.5. Plasmid Integration Confirmation
2.6. Measurement of CIT Content
2.7. Library Construction and Sequencing
2.8. Quality Control and Read Mapping
2.9. Differentially Expressed Genes (DEGs) and Functional Enrichment
2.10. Reverse Transcription Quantitative PCR (RT-qPCR) Analysis of Genes Involved in CIT Biosynthesis
2.11. Analysis of Alternative Splicing
2.12. Statistical Analysis
3. Results
3.1. CIT Content Analysis
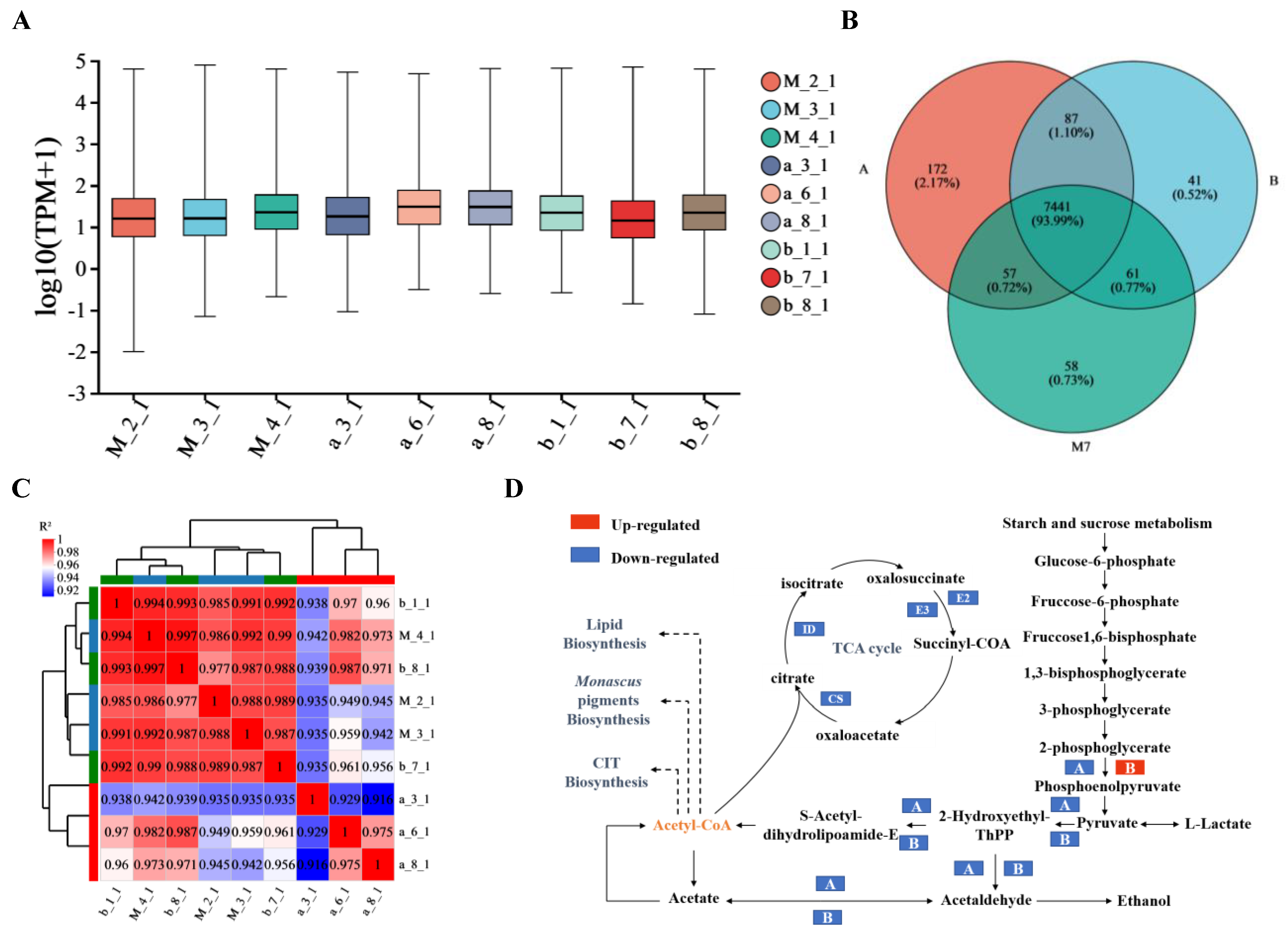
3.2. RNA Sequencing and Transcript Assembling
3.3. Gene Expression Analysis
3.4. DEG Analysis
3.5. Functional Annotation Analysis
3.6. Functional Enrichment Analysis
3.7. Analysis of AS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Chen, W.P.; Chen, R.F.; Liu, Q.P.; He, Y.; He, K.; Ding, X.L.; Kang, L.J.; Guo, X.X.; Xie, N.N.; Zhou, Y.X.; et al. Orange, red, yellow: Biosynthesis of azaphilone pigments in Monascus fungi. Chem. Sci. 2017, 8, 4917–4925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liang, J.; Zhang, A.A.; Hao, S.; Zhang, H.; Zhu, Q.Q.; Sun, B.G.; Wang, C.T. Overexpression of monacolin K biosynthesis genes in the Monascus purpureus azaphilone polyketide pathway. J. Agric. Food Chem. 2019, 67, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Long, P.C.; Zhu, L.S.; Lai, H.F.; Xu, S.Y.; Dong, X.X.; Shao, Y.C.; Wang, L.L.; Cheng, S.Y.; Liu, G.; He, J.R.; et al. Monascus red pigment liposomes: Microstructural characteristics, stability, and anticancer activity. Foods 2023, 12, 447. [Google Scholar] [CrossRef]
- Zhu, B.; Qi, F.Y.; Wu, J.I.; Yin, G.Q.; Hua, J.W.; Zhang, Q.Y.; Qin, L.P. Red yeast rice: A systematic review of the traditional uses, chemistry, pharmacology, and quality control of an important Chinese folk medicine. Front. Pharmacol. 2019, 10, 1449. [Google Scholar] [CrossRef]
- Yoon, H.R.; Ku, D.; Han, S.; Shin, S.C.; Kim, H.W.; Kim, H.J. Safety evaluation of mycotoxin citrinin production from Monascus ruber through whole-genome sequencing and analytical evaluation. 3 Biotech 2022, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, C.L.; Yang, M.R.; Liang, Z.E.N.; Wu, Y.; Kong, X.Y.; Fan, H.; Wang, S.Q.; Ning, C.; Xiao, W.G.; et al. The role of ER stress and ATP/AMPK in oxidative stress meditated hepatotoxicity induced by citrinin. Ecotoxicol. Environ. Saf. 2022, 237, 113531. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Gupta, A.; Pandhi, S.; Sharma, N.; Sharma, B.; Mishra, S.; Arora, S.; Selvakumar, R.; Saurabh, V.; et al. Citrinin mycotoxin contamination in food and feed: Impact on agriculture, human health, and detection and management strategies. Toxins 2022, 14, 85. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Yang, C.L.; Molnár, I.; Chen, S. Comparative transcriptomic analysis of key genes involved in citrinin biosynthesis in Monascus purpureus. J. Fungi 2023, 9, 200. [Google Scholar] [CrossRef]
- Hetherington, A.C.; Raistrick, H. Studies in the biochemistry of micro-organisms. Part XIV.—On the production and chemical constitution of a new yellow colouring mater, citrinin, produced from glucose by Penicillium. Philos. Trans. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character 1931, 220, 269–295. [Google Scholar] [CrossRef]
- Blanc, P.J.; Loret, M.O.; Goma, G. Production of citrinin by various species of Monascus. Biotechnol. Lett. 1995, 17, 291–294. [Google Scholar] [CrossRef]
- Birch, A.J.; Holloway, P.W.; Rickards, R.W. The biosynthesis of noviose, a branched-chain monosaccharide. Biochim. Biophy. Acta. Bioenerg. 1962, 57, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Sankawa, U.; Ebizuka, Y.; Noguchi, H.; Isikawa, Y.; Kitaghawa, S.; Yamamoto, Y.; Kobayashi, T.; Iitak, Y.; Seto, H. Biosynthesis of citrinin in Aspergillus terreus: Incorporation studies with [2-13C, 2-2H3], [1-13C, 18O2] and [1-13C, 17O]-acetate. Tetrahedron 1983, 39, 3583–3591. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tseng, C.P.; Chien, I.L.; Wang, W.Y.; Liaw, L.L.; Yuan, G.F. Exploring the distribution of citrinin biosynthesis related genes among Monascus species. J.Agric. Food Chem. 2008, 56, 11767–11772. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoshita, H.; Nihira, T. Identification and in vivo functional analysis by gene disruption of ctnA, an activator gene involved in citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2007, 73, 5097–5103. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef]
- Liu, H.Q.; Huang, Z.F.; Yang, S.Z.; Tian, X.F.; Wu, Z.Q. Inducing red pigment and inhibiting citrinin production by adding lanthanum (III) ion in Monascus purpureus fermentation. Appl. Environ. Microbiol. 2021, 105, 1905–1912. [Google Scholar] [CrossRef]
- He, S.S.; Wang, Y.L.; Xie, J.H.; Gao, H.; Li, X.J.; Huang, Z.B. 1H NMR-based metabolomic study of the effects of flavonoids on citrinin production by Monascus. Food Res. Int. 2020, 137, 109532. [Google Scholar] [CrossRef] [PubMed]
- Grützmann, K.; Szafranski, K.; Pohl, M.; Voigt, K.; Petzold, A.; Schuster, S. Fungal alternative splicing is associated with multicellular complexity and virulence: A genome-wide multi-species study. DNA Res. 2013, 21, 27–39. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Miyake, T. Development and media regulate alternative splicing of a methyltransferase pre-mRNA in Monascus pilosus. J. Agric. Food Chem. 2009, 57, 4162–4167. [Google Scholar] [CrossRef]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef]
- Yuan, Y.H.; Liu, C.J.; Zhao, G.; Gong, X.W.; Dang, K.; Yang, Q.H.; Feng, B.L. Transcriptome analysis reveals the mechanism associated with dynamic changes in fatty acid and phytosterol content in foxtail millet (Setaria italica) during seed development. Food Res. Inter. 2021, 145, 110429. [Google Scholar] [CrossRef]
- Ouyang, W.B.; Liu, X.; Wang, Y.L.; Huang, Z.B.; Li, X.J. Addition of genistein to the fermentation process reduces citrinin production by Monascus via changes at the transcription level. Food Chem. 2021, 343, 128410. [Google Scholar] [CrossRef] [PubMed]
- Zorin, E.A.; Afonin, A.M.; Kulaeva, O.A.; Gribchenko, E.S.; Shtark, O.Y.; Zhukov, V.A. Transcriptome analysis of alternative splicing events induced by arbuscular mycorrhizal fungi (Rhizophagus irregularis) in pea (Pisum sativum L.) roots. Plants 2020, 9, 1700. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.C.; Ding, Y.D.; Zhao, Y.; Yang, S.; Xie, B.J.; Chen, F.S. Characteristic analysis of transformants in T-DNA mutation library of Monascus ruber. World J. Microbiol. Biotechnol. 2009, 25, 989–995. [Google Scholar] [CrossRef]
- Xiao, Y.H.; Yin, M.H.; Hou, L.; Pei, Y. Direct amplification of intron-containing hairpin RNA construct from genomic DNA. Biotechniques 2006, 41, 548–552. [Google Scholar] [CrossRef]
- Guo, X.Y.; Atehli, D.; Chen, M.H.; Chen, D.; Wang, Y.R. A Zn(II)(2)Cys(6) transcription factor MPsGeI suppresses pigment biosynthesis in Monascus. Int. J. Biol. Macromol. 2023, 233, 123504. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Xie, L.M.; Xie, J.H.; Chen, X.X.; Tao, X.; Xie, J.Y.; Shi, X.Y.; Huang, Z.B. Comparative transcriptome analysis of Monascus purpureus at different fermentation times revealed candidate genes involved in exopolysaccharide biosynthesis. Food Res. Int. 2022, 160, 111700. [Google Scholar] [CrossRef]
- Shen, S.H.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Nati. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef]
- Yang, X.C.; Li, X.Y.; Zhao, J.L.; Xie, M.S.; Li, X.Y.; Jia, B.; Huang, Y. Transcriptome analysis reveals the involvement of alternative splicing in the nitrogen starvation response of Chlamydomonas reinhardtii. Processes 2022, 10, 2719. [Google Scholar] [CrossRef]
- Huang, Z.B.; Zhang, S.Y.; Xu, Y.; Li, L.S.; Li, Y.P. Metabolic effects of the pksCT gene on Monascus aurantiacus Li As3.4384 using gas chromatography--time-of-flight mass spectrometry-based metabolomics. J. Agric. Food Chem. 2016, 64, 1565–1574. [Google Scholar] [CrossRef]
- Li, Y.P.; Tang, X.; Wu, W.; Xu, Y.; Huang, Z.B.; He, Q.H. The ctnG gene encodes carbonic anhydrase involved in mycotoxin citrinin biosynthesis from Monascus aurantiacus. Food Addi. Contam. A 2015, 32, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Liang, X.L.; Li, Y.D.; Wang, P.M.; Keller, N.P. Genetic Regulation of Mycotoxin Biosynthesis. J. Fungi 2023, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.N.; Liu, Q.P.; Chen, F.S. Deletion of pigR gene in Monascus ruber leads to loss of pigment production. Biotechnol. Lett. 2013, 35, 1425–1432. [Google Scholar] [CrossRef]
- Li, Y.P.; Wang, N.; Jiao, X.X.; Tu, Z.; He, Q.H.; Fu, J.H. The ctnF gene is involved in citrinin and pigment synthesis in Monascus aurantiacus. J. Basic Microbiol. 2020, 60, 873–881. [Google Scholar] [CrossRef]
- Duan, Y.L.; Du, Y.; Yi, Z.; Wang, Z.; Pei, X.L.; Wei, X.T.; Li, M. Systematic metabolic engineering for the production of azaphilones in Monascus purpureus HJ11. J. Agric. Food Chem. 2022, 70, 1589–1600. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef]
- Fang, S.M.; Hou, X.; Qiu, K.H.; He, R.; Feng, X.S.; Liang, X.L. The occurrence and function of alternative splicing in fungi. Fungal Biol. Rev. 2020, 34, 178–188. [Google Scholar] [CrossRef]
- Liu, X.Y.; Fan, L.; Gao, J.; Shen, X.Y.; Hou, C.L. Global identification of alternative splicing in Shiraia bambusicola and analysis of its regulation in hypocrellin biosynthesis. Appl. Environ. Microbiol. 2020, 104, 211–223. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Sequence (5′-3′) |
|---|---|---|
| neo | Xho I-neo Forward | CCGCTCGAGCCAACTCAACCCCATCG |
| EcoR I-neo Reverse | CGGAATTCATCATCATGCAACATGC | |
| PtrpC (α) | Hind III-PtrpC (α) Forward | CCCAAGCTTGTCGACAGAAGATGATATTG |
| PtrpC (α)-Reverse | CCCAATCACTCTTGATGATTGGGGCGTATGATCATACATATCGATGCTTGGGTAGAATA | |
| TtrpC (α) | TtrpC (α) Forward | ATCATCAAGAGTGATTGGGGCGTATGATCATATTTTTCCAAGCAGCAAAGAGTGCCTTCTAG |
| Xba I-TtrpC (α) Reverse | GCTCTAGAAAGAAGGATTACCTC | |
| PtrpC (α + β) | Hind III-PtrpC Forward | CCCAAGCTTGTCGACAGAAGATGATATTG |
| PtrpC Reverse | TCAATCTCCTCTTGAGAGATTGATGAGGTTCTGCATACATATCGATGCTTGGGTAGAATA | |
| TtrpC (α + β) | TtrpC Forward | TCTCTCAAGAGGAGATTGATGAGGTTCTGCATTTTTCCAAGCAGCAAAGAGTGCCTTCTAG |
| Xba I-TtrpC Reverse | GCTCTAGAAAGAAGGATTACCTC |
| Gene | Predict Function | M7 vs. ihpRNA-pksCTα | M7 vs. ihpRNA-pksCT (α + β) | ||||
|---|---|---|---|---|---|---|---|
| Log2FC | Regulate | Significant (p < 0.05) | Log2FC | Regulate | Significant (p < 0.05) | ||
| mrl1 | Serine hydrolase | 0.09 | up | no | −0.06 | down | no |
| mrl2 | 2-oxoglutarate-dependent dioxygenase | 0 | - | no | 0.17 | up | no |
| mrl3 | Citrinin biosynthesis transcriptional activator | −0.49 | down | no | −0.57 | down | no |
| mrl4 | Aldehyde dehydrogenase | −0.58 | down | no | −0.48 | down | no |
| mrl5 | Glyoxalase-like domain | −1.31 | down | yes | −1.50 | down | yes |
| mrl6 | Short chain dehydrogenase | −3.59 | down | yes | −3.2 | down | yes |
| mrl7 | Glucose methanol choline (GMC) oxidoreductase | −1.21 | down | yes | −0.27 | down | no |
| pksCT | Methyltransferase activity | −0.73 | down | no | −0.43 | down | no |
| mrr1 | Major facilitator superfamily (MFS) protein | −1.05 | down | no | −1.59 | down | yes |
| mrr2 | Histidine phosphatase | 0.073 | up | no | 0.258 | up | no |
| mrr3 | Function unknown | 0.08 | up | no | 0.13 | up | no |
| mrr4 | WD repeat protein | −0.43 | down | no | 0.11 | up | no |
| mrr5 | Carbonic anhydrase | −0.22 | down | no | −8.95 | down | no |
| mrr8 | AMP-binding enzyme | −0.36 | down | no | −0.19 | down | no |
| pigR | pigment biosynthesis activator | −0.11 | down | no | 0.01 | up | no |
| Category | Pathway | Putative Functions of DEGs | Regulation |
|---|---|---|---|
| Carbohydrate metabolism | Butanoate metabolism | 3-oxoacyl-[acyl-carrier-protein] reductase (FabG) | down |
| Pentose and glucuronate interconversions | L-arabinitol 4-dehydrogenase | Down (not detected in B group) | |
| D-arabinitol dehydrogenase 1 | up | ||
| Inositol phosphate metabolism | scyllo-inositol 2-dehydrogenase | down | |
| 3-phytase A | up | ||
| Galactose metabolism | D-galactonate dehydratase | up | |
| Starch and sucrose metabolism | Probable beta-glucosidase A | up | |
| Pentose phosphate pathway | Ribokinase | down | |
| Glyoxylate and dicarboxylate metabolism | Catalase | down | |
| Glycerate dehydrogenase | down | ||
| Amino sugar and nucleotide sugar metabolism | Chitin synthase | up | |
| hypothetical protein | up | ||
| N-acetylglucosamine-6-phosphate deacetylase | down | ||
| Glycolysis/Gluconeogenesis | Enolase | up | |
| Pyruvate decarboxylase | down | ||
| Amino acid metabolism | Lysine degradation | L-saccharopine oxidase | down |
| Glycine, serine and threonine metabolism | NAD/NADP-dependent betaine aldehyde dehydrogenase | down | |
| Glycerate dehydrogenase | down | ||
| Tryptophan metabolism | Indoleamine 2,3-dioxygenase | down | |
| cytochrome P450 oxidoreductase | up | ||
| Kynureninase 2 | down | ||
| Phenylalanine metabolism | Phenylacetate 2-hydroxylase | down | |
| Probable 4-hydroxyphenylpyruvate dioxygenase 1 | down | ||
| Valine, leucine and isoleucine degradation | Ketol-acid reductoisomerase, mitochondrial | up | |
| Tyrosine metabolism | Probable 4-hydroxyphenylpyruvate dioxygenase 1 | down | |
| Arginine and proline meta | Ornithine aminotransferase | down | |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | Quinate dehydrogenase | down | |
| Lipid metabolism | Fatty acid degradation | cytochrome P450 oxidoreductase | up |
| Fatty acid biosynthesis | Fatty acid synthase subunit alpha | up | |
| Sphingolipid metabolism | Serine palmitoyltransferase 2 | up | |
| Steroid biosynthesis | Eburicol 14-alpha-demethylase | up | |
| Sterol 24-C-methyltransferase | up | ||
| C-5 sterol desaturase | up | ||
| Methylsterol monooxygenase | up | ||
| Biosynthesis of unsaturated fatty acids | Acyl-CoA desaturase | up | |
| Oleate hydroxylase FAH12 | up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Zhu, L.; Dong, X.; Li, A.; Xu, S.; Wang, L.; Shao, Y. Metabolic Regulation of Two pksCT Gene Transcripts in Monascus ruber Impacts Citrinin Biosynthesis. J. Fungi 2023, 9, 1174. https://doi.org/10.3390/jof9121174
He Y, Zhu L, Dong X, Li A, Xu S, Wang L, Shao Y. Metabolic Regulation of Two pksCT Gene Transcripts in Monascus ruber Impacts Citrinin Biosynthesis. Journal of Fungi. 2023; 9(12):1174. https://doi.org/10.3390/jof9121174
Chicago/Turabian StyleHe, Yi, Lisha Zhu, Xingxing Dong, Aoran Li, Suyin Xu, Liling Wang, and Yanchun Shao. 2023. "Metabolic Regulation of Two pksCT Gene Transcripts in Monascus ruber Impacts Citrinin Biosynthesis" Journal of Fungi 9, no. 12: 1174. https://doi.org/10.3390/jof9121174