

Moonlighting Proteins: Diverse Functions Found in Fungi

Abstract
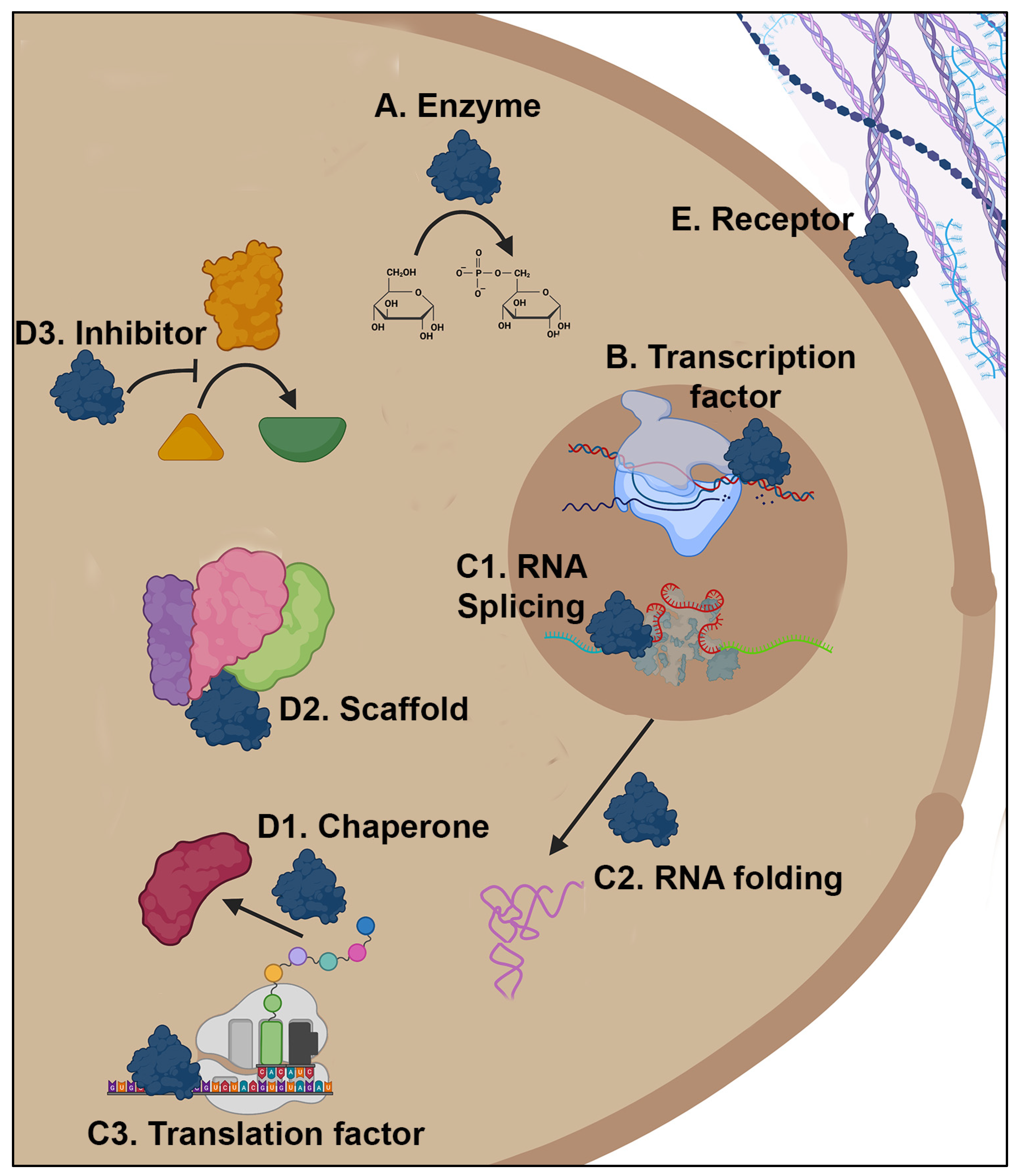
:1. Introduction
2. Examples of Moonlighting Proteins in Fungi
2.1. Affecting Transcription, DNA Metabolism, or DNA Maintenance
2.2. Affecting Translation or RNA Metabolism
2.3. Affecting Another Protein’s Activity
2.4. Intracellular Proteins with an Additional Function on the Cell Surface
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, H.; Zabad, S.; Rivera, N.; Rowin, E.; Hassan, M.; Gomez De Jesus, S.M.; Llinas Santos, P.S.; Kravchenko, K.; Mikhova, M.; et al. MoonProt 3.0: An update of the moonlighting proteins database. Nucleic Acids Res. 2020, 49, D368–D372. [Google Scholar] [CrossRef]
- Bolduc, J.M.; Spiegel, P.C.; Chatterjee, P.; Brady, K.L.; Downing, M.E.; Caprara, M.G.; Waring, R.B.; Stoddard, B.L. Structural and biochemical analyses of DNA and RNA binding by a bifunctional homing endonuclease and group I intron splicing factor. Genes Dev. 2003, 17, 2875–2888. [Google Scholar] [CrossRef]
- Ho, Y.; Kim, S.J.; Waring, R.B. A protein encoded by a group I intron in Aspergillus nidulans directly assists RNA splicing and is a DNA endonuclease. Proc. Natl. Acad. Sci. USA 1997, 94, 8994–8999. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.; Walker-Jonah, A.; Hollenberg, C.P. Galactokinase encoded by GAL1 is a bifunctional protein required for induction of the GAL genes in Kluyveromyces lactis and is able to suppress the gal3 phenotype in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 5454–5461. [Google Scholar] [PubMed]
- Bhat, P.J.; Oh, D.; Hopper, J.E. Analysis of the GAL3 signal transduction pathway activating GAL4 protein dependent transcription in Saccharomyces cerevisiae. Genetics 1990, 125, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Montanini, B.; Levati, E.; Bolchi, A.; Kohler, A.; Morin, E.; Tisserant, E.; Martin, F.; Ottonello, S. Genome-wide search and functional identification of transcription factors in the mycorrhizal fungus Tuber melanosporum. New Phytol. 2011, 189, 736–750. [Google Scholar] [CrossRef]
- Levati, E.; Sartini, S.; Bolchi, A.; Ottonello, S.; Montanini, B. Moonlighting transcriptional activation function of a fungal sulfur metabolism enzyme. Sci. Rep. 2016, 6, 25165. [Google Scholar] [CrossRef]
- Bateman, J.M.; Perlman, P.S.; Butow, R.A. Mutational bisection of the mitochondrial DNA stability and amino acid biosynthetic functions of ilv5p of budding yeast. Genetics 2002, 161, 1043–1052. [Google Scholar] [CrossRef]
- Hall, D.A.; Zhu, H.; Zhu, X.; Royce, T.; Gerstein, M.; Snyder, M. Regulation of gene expression by a metabolic enzyme. Science 2004, 306, 482–484. [Google Scholar] [CrossRef]
- El Bakkoury, M.; Dubois, E.; Messenguy, F. Recruitment of the yeast MADS box proteins, ArgRI and Mcm1 by the pleiotropic factor ArgRIII is required for their stability. Mol. Microbiol. 2000, 35, 15–31. [Google Scholar] [CrossRef]
- Ringel, A.E.; Ryznar, R.; Picariello, H.; Huang, K.L.; Lazarus, A.G.; Holmes, S.G. Yeast Tdh3 (glyceraldehyde 3-phosphate dehydrogenase) is a Sir2-interacting factor that regulates transcriptional silencing and rDNA recombination. PLoS Genet. 2013, 9, e1003871. [Google Scholar] [CrossRef]
- Ahuatzi, D.; Herrero, P.; de la Cera, T.; Moreno, F. The glucose regulated nuclear localization of hexokinase 2 in Saccharomyces cerevisiae is Mig1-dependent. J. Biol. Chem. 2004, 279, 14440–14446. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; De La Cera, T.; Herrero, P.; Moreno, F. The hexokinase 2 protein regulates the expression of the GLK1, HXK1 and HXK2 genes of Saccharomyces cerevisiae. Biochem. J. 2001, 355 Pt 3, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, B.A.; Kolesar, J.E.; Perlman, P.S.; Butow, R.A. A function for the mitochondrial chaperonin Hsp60 in the structure and transmission of mitochondrial DNA nucleoids in Saccharomyces cerevisiae. J. Cell. Biol. 2003, 163, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Torres-Machorro, A.L.; Aris, J.P.; Pillus, L. A moonlighting metabolic protein influences repair at DNA double-stranded breaks. Nucleic Acids Res. 2015, 43, 1646–1658. [Google Scholar] [CrossRef]
- Bermingham-McDonogh, O.; Gralla, E.B.; Valentine, J.S. The copper, zinc superoxide dismutase gene of Saccharomyces cerevisiae: Cloning, sequencing, and biological activity. Proc. Natl. Acad. Sci. USA 1988, 85, 4789–4793. [Google Scholar] [CrossRef]
- Tsang, C.K.; Liu, Y.; Thomas, J.; Zhang, Y.; Zheng, X.F. Superoxide dismutase 1 acts as a nuclear transcription factor to regulate oxidative stress resistance. Nat. Commun. 2014, 5, 3446. [Google Scholar] [CrossRef]
- Coschigano, P.W.; Magasanik, B. The URE2 gene product of Saccharomyces cerevisiae plays an important role in the cellular response to the nitrogen source and has homology to glutathione S-transferases. Mol. Cell. Biol. 1991, 11, 822–832. [Google Scholar]
- Blinder, D.; Coschigano, P.W.; Magasanik, B. Interaction of the GATA factor Gln3p with the nitrogen regulator Ure2p in Saccharomyces cerevisiae. J. Bacteriol. 1996, 178, 4734–4736. [Google Scholar] [CrossRef]
- Courchesne, W.E.; Magasanik, B. Regulation of nitrogen assimilation in Saccharomyces cerevisiae: Roles of the URE2 and GLN3 genes. J. Bacteriol. 1988, 170, 708–713. [Google Scholar] [CrossRef]
- Bai, M.; Zhou, J.M.; Perrett, S. The yeast prion protein Ure2 shows glutathione peroxidase activity in both native and fibrillar forms. J. Biol. Chem. 2004, 279, 50025–50030. [Google Scholar] [CrossRef]
- Ducett, J.K.; Peterson, F.C.; Hoover, L.A.; Prunuske, A.J.; Volkman, B.F.; Craig, E.A. Unfolding of the C-terminal domain of the J-protein Zuo1 releases autoinhibition and activates Pdr1 dependent transcription. J. Mol. Biol. 2013, 425, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Caprara, M.G.; Mohr, G.; Lambowitz, A.M. A tyrosyl-tRNA synthetase protein induces tertiary folding of the group I intron catalytic core. J. Mol. Biol. 1996, 257, 512–531. [Google Scholar] [CrossRef]
- Elzinga, S.D.; Bednarz, A.L.; van Oosterum, K.; Dekker, P.J.; Grivell, L.A. Yeast mitochondrial NAD(+)-dependent isocitrate dehydrogenase is an RNA-binding protein. Nucleic Acids Res. 1993, 21, 5328–5331. [Google Scholar] [CrossRef] [PubMed]
- Presutti, C.; Ciafre, S.A.; Bozzoni, I. The ribosomal protein L2 in S. cerevisiae controls the level of accumulation of its own mRNA. EMBO J. 1991, 10, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Eng, F.J.; Warner, J.R. Structural basis for the regulation of splicing of a yeast messenger RNA. Cell 1991, 65, 797–804. [Google Scholar] [CrossRef]
- Sarkar, J.; Poruri, K.; Boniecki, M.T.; McTavish, K.K.; Martinis, S.A. Yeast mitochondrial leucyl-tRNA synthetase CP1 domain has functionally diverged to accommodate RNA splicing at expense of hydrolytic editing. J. Biol. Chem. 2012, 287, 14772–14781. [Google Scholar] [CrossRef]
- Fewell, S.W.; Woolford, J.L., Jr. Ribosomal protein S14 of Saccharomyces cerevisiae regulates its expression by binding to RPS14B pre-mRNA and to 18S rRNA. Mol. Cell. Biol. 1999, 19, 826–834. [Google Scholar] [CrossRef]
- Badis, G.; Saveanu, C.; Fromont-Racine, M.; Jacquier, A. Targeted mRNA degradation by deadenylation-independent decapping. Mol. Cell 2004, 15, 5–15. [Google Scholar] [CrossRef]
- Ozimek, P.; van Dijk, R.; Latchev, K.; Gancedo, C.; Wang, D.Y.; van der Klei, I.J.; Veenhuis, M. Pyruvate carboxylase is an essential protein in the assembly of yeast peroxisomal oligomeric alcohol oxidase. Mol. Biol. Cell 2003, 14, 786–797. [Google Scholar] [CrossRef]
- Ozimek, P.Z.; Klompmaker, S.H.; Visser, N.; Veenhuis, M.; van der Klei, I.J. The transcarboxylase domain of pyruvate carboxylase is essential for assembly of the peroxisomal flavoenzyme alcohol oxidase. FEMS Yeast Res. 2007, 7, 1082–1092. [Google Scholar] [CrossRef]
- Yuan, W.; Tuttle, D.L.; Shi, Y.J.; Ralph, G.S.; Dunn, W.A., Jr. Glucose-induced microautophagy in Pichia pastoris requires the alpha-subunit of phosphofructokinase. J. Cell Sci. 1997, 110 Pt 16, 1935–1945. [Google Scholar] [CrossRef]
- Bechet, J.; Wiame, J.M. Indication of a specific regulatory binding protein for ornithinetranscarbamylase in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1965, 21, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Wang, X.; Purring-Koch, C.; Wei, Y.; McLendon, G.L. A mutational epitope for cytochrome C binding to the apoptosis protease activation factor-1. J. Biol. Chem. 2001, 276, 13034–13038. [Google Scholar] [CrossRef] [PubMed]
- Decker, B.L.; Wickner, W.T. Enolase activates homotypic vacuole fusion and protein transport to the vacuole in yeast. J. Biol. Chem. 2006, 281, 14523–14528. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Holliday, L.S.; Zhang, L.; Dunn, W.A., Jr.; Gluck, S.L. Interaction between aldolase and vacuolar H+-ATPase: Evidence for direct coupling of glycolysis to the ATP-hydrolyzing proton pump. J. Biol. Chem. 2001, 276, 30407–30413. [Google Scholar] [CrossRef] [PubMed]
- Casamayor, A.; Arino, J. Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins. J. Fungi. 2022, 8, 1066. [Google Scholar] [CrossRef]
- Ruiz, A.; Gonzalez, A.; Munoz, I.; Serrano, R.; Abrie, J.A.; Strauss, E.; Arino, J. Moonlighting proteins Hal3 and Vhs3 form a heteromeric PPCDC with Ykl088w in yeast CoA biosynthesis. Nat. Chem. Biol. 2009, 5, 920–928. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two enzymes in one; two yeast peroxiredoxins display oxidative stress-dependent switching from a peroxidase to a molecular chaperone function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef]
- Sengupta, J.; Nilsson, J.; Gursky, R.; Spahn, C.M.; Nissen, P.; Frank, J. Identification of the versatile scaffold protein RACK1 on the eukaryotic ribosome by cryo-EM. Nat. Struct. Mol. Biol. 2004, 11, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Kadrmas, J.L.; Smith, M.A.; Pronovost, S.M.; Beckerle, M.C. Characterization of RACK1 function in Drosophila development. Dev. Dyn. 2007, 236, 2207–2215. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Huo, X.; Zhai, Y.; Wang, A.; Xu, J.; Su, D.; Bartlam, M.; Rao, Z. Crystal structure of mitochondrial respiratory membrane protein complex II. Cell 2005, 121, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
- Lemire, B.D.; Oyedotun, K.S. The Saccharomyces cerevisiae mitochondrial succinate:ubiquinone oxidoreductase. Biochim. Biophys. Acta 2002, 1553, 102–116. [Google Scholar] [CrossRef]
- Yankovskaya, V.; Horsefield, R.; Tornroth, S.; Luna-Chavez, C.; Miyoshi, H.; Leger, C.; Byrne, B.; Cecchini, G.; Iwata, S. Architecture of succinate dehydrogenase and reactive oxygen species generation. Science 2003, 299, 700–704. [Google Scholar] [CrossRef]
- Gebert, N.; Gebert, M.; Oeljeklaus, S.; von der Malsburg, K.; Stroud, D.A.; Kulawiak, B.; Wirth, C.; Zahedi, R.P.; Dolezal, P.; Wiese, S.; et al. Dual function of Sdh3 in the respiratory chain and TIM22 protein translocase of the mitochondrial inner membrane. Mol. Cell 2011, 44, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Beitner, R.; Nordenberg, J. Inhibition of 6-phosphogluconate dehydrogenase (decarboxylating) by glucose 1,6-bisphosphate. Biochim. Biophys. Acta 1979, 583, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Seweryn, K.; Karkowska-Kuleta, J.; Wolak, N.; Bochenska, O.; Kedracka-Krok, S.; Kozik, A.; Rapala-Kozik, M. Kinetic and thermodynamic characterization of the interactions between the components of human plasma kinin-forming system and isolated and purified cell wall proteins of Candida albicans. Acta Biochim. Pol. 2015, 62, 825–835. [Google Scholar] [CrossRef]
- Karkowska-Kuleta, J.; Kedracka-Krok, S.; Rapala-Kozik, M.; Kamysz, W.; Bielinska, S.; Karafova, A.; Kozik, A. Molecular determinants of the interaction between human high molecular weight kininogen and Candida albicans cell wall: Identification of kininogen-binding proteins on fungal cell wall and mapping the cell wall binding regions on kininogen molecule. Peptides 2011, 32, 2488–2496. [Google Scholar] [CrossRef]
- Crowe, J.D.; Sievwright, I.K.; Auld, G.C.; Moore, N.R.; Gow, N.A.; Booth, N.A. Candida albicans binds human plasminogen: Identification of eight plasminogen-binding proteins. Mol. Microbiol. 2003, 47, 1637–1651. [Google Scholar] [CrossRef]
- Kunert, A.; Losse, J.; Gruszin, C.; Huhn, M.; Kaendler, K.; Mikkat, S.; Volke, D.; Hoffmann, R.; Jokiranta, T.S.; Seeberger, H.; et al. Immune evasion of the human pathogen Pseudomonas aeruginosa: Elongation factor Tuf is a factor H and plasminogen binding protein. J. Immunol. 2007, 179, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Jong, A.Y.; Chen, S.H.M.; Stins, M.F.; Kim, K.S.; Tuan, T.L.; Huang, S.H. Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J. Med. Microbiol. 2003, 52 Pt 8, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Gozalbo, D.; Gil-Navarro, I.; Azorin, I.; Renau-Piqueras, J.; Martinez, J.P.; Gil, M.L. The cell wall-associated glyceraldehyde-3-phosphate dehydrogenase of Candida albicans is also a fibronectin and laminin binding protein. Infect. Immun. 1998, 66, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; De Micheli, M.; Coleman, S.T.; Sanglard, D.; Moye-Rowley, W.S. Analysis of the oxidative stress regulation of the Candida albicans transcription factor, Cap1p. Mol. Microbiol. 2000, 36, 618–629. [Google Scholar] [CrossRef]
- Gil-Bona, A.; Amador-Garcia, A.; Gil, C.; Monteoliva, L. The external face of Candida albicans: A proteomic view of the cell surface and the extracellular environment. J. Proteom. 2018, 180, 70–79. [Google Scholar] [CrossRef]
- Luo, S.; Hoffmann, R.; Skerka, C.; Zipfel, P.F. Glycerol-3-phosphate dehydrogenase 2 is a novel factor H-, factor H-like protein 1-, and plasminogen-binding surface protein of Candida albicans. J. Infect. Dis. 2013, 207, 594–603. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Gutiérrez-Sánchez, G.; López-Romero, E.; Ruiz-Baca, E.; Villagómez-Castro, J.; Rodríguez-Sifuentes, L. Identification of Candida albicans heat shock proteins and Candida glabrata and Candida krusei enolases involved in the response to oxidative stress. Open Life Sci. 2013, 8, 337–345. [Google Scholar] [CrossRef]
- Lesiak-Markowicz, I.; Vogl, G.; Schwarzmuller, T.; Speth, C.; Lass-Florl, C.; Dierich, M.P.; Kuchler, K.; Wurzner, R. Candida albicans Hgt1p, a multifunctional evasion molecule: Complement inhibitor, CR3 analogue, and human immunodeficiency virus-binding molecule. J. Infect. Dis. 2011, 204, 802–809. [Google Scholar] [CrossRef]
- Gilmore, B.J.; Retsinas, E.M.; Lorenz, J.S.; Hostetter, M.K. An iC3b receptor on Candida albicans: Structure, function, and correlates for pathogenicity. J. Infect. Dis. 1988, 157, 38–46. [Google Scholar] [CrossRef]
- Hostetter, M.K.; Lorenz, J.S.; Preus, L.; Kendrick, K.E. The iC3b receptor on Candida albicans: Subcellular localization and modulation of receptor expression by glucose. J. Infect. Dis. 1990, 161, 761–768. [Google Scholar] [CrossRef]
- Satala, D.; Zelazna, A.; Satala, G.; Bukowski, M.; Zawrotniak, M.; Rapala-Kozik, M.; Kozik, A. Towards understanding the novel adhesin function of Candida albicans phosphoglycerate mutase at the pathogen cell surface: Some structural analysis of the interactions with human host extracellular matrix proteins. Acta Biochim. Pol. 2021, 68, 515–525. [Google Scholar] [CrossRef]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Satala, D.; Satala, G.; Karkowska-Kuleta, J.; Bukowski, M.; Kluza, A.; Rapala-Kozik, M.; Kozik, A. Structural Insights into the Interactions of Candidal Enolase with Human Vitronectin, Fibronectin and Plasminogen. Int. J. Mol. Sci. 2020, 21, 7843. [Google Scholar] [CrossRef]
- Kozik, A.; Karkowska-Kuleta, J.; Zajac, D.; Bochenska, O.; Kedracka-Krok, S.; Jankowska, U.; Rapala-Kozik, M. Fibronectin-, vitronectin- and laminin-binding proteins at the cell walls of Candida parapsilosis and Candida tropicalis pathogenic yeasts. BMC Microbiol. 2015, 15, 197. [Google Scholar] [CrossRef]
- Stie, J.; Bruni, G.; Fox, D. Surface-associated plasminogen binding of Cryptococcus neoformans promotes extracellular matrix invasion. PLoS ONE 2009, 4, e5780. [Google Scholar] [CrossRef] [PubMed]
- Wampler, J.L.; Kim, K.P.; Jaradat, Z.; Bhunia, A.K. Heat shock protein 60 acts as a receptor for the Listeria adhesion protein in Caco-2 cells. Infect. Immun. 2004, 72, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Donofrio, F.C.; Calil, A.C.A.; Miranda, E.T.; Almeida, A.M.F.; Benard, G.; Soares, C.P.; Veloso, S.N.; Soares, C.M.A.; Mendes Giannini, M.J.S. Enolase from Paracoccidioides brasiliensis: Isolation and identification as a fibronectin-binding protein. J. Med. Microbiol. 2009, 58 Pt 6, 706–713. [Google Scholar] [CrossRef]
- Barbosa, M.S.; Bao, S.N.; Andreotti, P.F.; de Faria, F.P.; Felipe, M.S.; dos Santos Feitosa, L.; Mendes-Giannini, M.J.; Soares, C.M. Glyceraldehyde-3-phosphate dehydrogenase of Paracoccidioides brasiliensis is a cell surface protein involved in fungal adhesion to extracellular matrix proteins and interaction with cells. Infect. Immun. 2006, 74, 382–389. [Google Scholar] [CrossRef]
- Pereira, L.A.; Bao, S.N.; Barbosa, M.S.; da Silva, J.L.; Felipe, M.S.; de Santana, J.M.; Mendes-Giannini, M.J.; de Almeida Soares, C.M. Analysis of the Paracoccidioides brasiliensis triosephosphate isomerase suggests the potential for adhesin function. FEMS Yeast Res. 2007, 7, 1381–1388. [Google Scholar] [CrossRef]
- Garcia-Carnero, L.C.; Salinas-Marin, R.; Lozoya-Perez, N.E.; Wrobel, K.; Wrobel, K.; Martinez-Duncker, I.; Nino-Vega, G.A.; Mora-Montes, H.M. The Heat Shock Protein 60 and Pap1 Participate in the Sporothrixschenckii-Host Interaction. J. Fungi. 2021, 7, 960. [Google Scholar] [CrossRef]
- Lau, S.K.; Tse, H.; Chan, J.S.; Zhou, A.C.; Curreem, S.O.; Lau, C.C.; Yuen, K.Y.; Woo, P.C. Proteome profiling of the dimorphic fungus Penicillium marneffei extracellular proteins and identification of glyceraldehyde-3-phosphate dehydrogenase as an important adhesion factor for conidial attachment. FEBS J. 2013, 280, 6613–6626. [Google Scholar] [CrossRef] [PubMed]
- Satala, D.; Bras, G.; Kozik, A.; Rapala-Kozik, M.; Karkowska-Kuleta, J. More than Just Protein Degradation: The Regulatory Roles and Moonlighting Functions of Extracellular Proteases Produced by Fungi Pathogenic for Humans. J. Fungi. 2023, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- Lipke, P.N.; Ragonis-Bachar, P. Sticking to the Subject: Multifunctionality in Microbial Adhesins. J. Fungi. 2023, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, T.; Mittal, N.; Janga, S.C.; Gerber, A.P. A screen for RNA-binding proteins in yeast indicates dual functions for many enzymes. PLoS ONE 2010, 5, e15499. [Google Scholar] [CrossRef] [PubMed]
- Tsvetanova, N.G.; Klass, D.M.; Salzman, J.; Brown, P.O. Proteome-wide search reveals unexpected RNA-binding proteins in Saccharomyces cerevisiae. PLoS ONE 2010, 5, e12671. [Google Scholar] [CrossRef]
- Kramer, K.; Sachsenberg, T.; Beckmann, B.M.; Qamar, S.; Boon, K.L.; Hentze, M.W.; Kohlbacher, O.; Urlaub, H. Photo-cross-linking and high-resolution mass spectrometry for assignment of RNA-binding sites in RNA-binding proteins. Nat. Methods 2014, 11, 1064–1070. [Google Scholar] [CrossRef]
- Beckmann, B.M.; Horos, R.; Fischer, B.; Castello, A.; Eichelbaum, K.; Alleaume, A.M.; Schwarzl, T.; Curk, T.; Foehr, S.; Huber, W.; et al. The RNA-binding proteomes from yeast to man harbour conserved enigmRBPs. Nat. Commun. 2015, 6, 10127. [Google Scholar] [CrossRef] [PubMed]
- Matia-Gonzalez, A.M.; Laing, E.E.; Gerber, A.P. Conserved mRNA-binding proteomes in eukaryotic organisms. Nat. Struct. Mol. Biol. 2015, 22, 1027–1033. [Google Scholar] [CrossRef]
- Shchepachev, V.; Bresson, S.; Spanos, C.; Petfalski, E.; Fischer, L.; Rappsilber, J.; Tollervey, D. Defining the RNA interactome by total RNA-associated protein purification. Mol. Syst. Biol. 2019, 15, e8689. [Google Scholar] [CrossRef]
- Curtis, N.J.; Jeffery, C.J. The expanding world of metabolic enzymes moonlighting as RNA binding proteins. Biochem. Soc. Trans. 2021, 49, 1099–1108. [Google Scholar] [CrossRef]
- Caudron-Herger, M.; Jansen, R.E.; Wassmer, E.; Diederichs, S. RBP2GO: A comprehensive pan-species database on RNA-binding proteins, their interactions and functions. Nucleic Acids Res. 2020, 49, D425–D436. [Google Scholar] [CrossRef] [PubMed]
- Molero, C.; Petrenyi, K.; Gonzalez, A.; Carmona, M.; Gelis, S.; Abrie, J.A.; Strauss, E.; Ramos, J.; Dombradi, V.; Hidalgo, E.; et al. The Schizosaccharomyces pombe fusion gene hal3 encodes three distinct activities. Mol. Microbiol. 2013, 90, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, C.J. Intracellular/surface moonlighting proteins that aid in the attachment of gut microbiota to the host. AIMS Microbiol. 2019, 5, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Kainulainen, V.; Korhonen, T.K. Dancing to another tune-adhesive moonlighting proteins in bacteria. Biology 2014, 3, 178–204. [Google Scholar] [CrossRef]
- Jeffery, C. Intracellular proteins moonlighting as bacterial adhesion factors. AIMS Microbiol. 2018, 4, 362–376. [Google Scholar] [CrossRef]
- Raymond, B.B.; Djordjevic, S. Exploitation of plasmin(ogen) by bacterial pathogens of veterinary significance. Vet. Microbiol. 2015, 178, 1–13. [Google Scholar] [CrossRef]
- Collen, D.; Verstraete, M. Molecular biology of human plasminogen. II. Metabolism in physiological and some pathological conditions in man. Thromb. Diath. Haemorrh. 1975, 34, 403–408. [Google Scholar]
- Dano, K.; Andreasen, P.A.; Grondahl-Hansen, J.; Kristensen, P.; Nielsen, L.S.; Skriver, L. Plasminogen activators, tissue degradation, and cancer. Adv. Cancer Res. 1985, 44, 139–266. [Google Scholar]
- Arvizu-Rubio, V.J.; Garcia-Carnero, L.C.; Mora-Montes, H.M. Moonlighting proteins in medically relevant fungi. PeerJ 2022, 10, e14001. [Google Scholar] [CrossRef]
- Satala, D.; Karkowska-Kuleta, J.; Zelazna, A.; Rapala-Kozik, M.; Kozik, A. Moonlighting Proteins at the Candidal Cell Surface. Microorganisms 2020, 8, 1046. [Google Scholar] [CrossRef]
- Wang, W.; Jeffery, C.J. An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol. Biosyst. 2016, 12, 1420–1431. [Google Scholar] [CrossRef]
- Jeffery, C.J. Protein species and moonlighting proteins: Very small changes in a protein’s covalent structure can change its biochemical function. J. Proteom. 2016, 134, 19–24. [Google Scholar] [CrossRef]
- Miura, N.; Ueda, M. Evaluation of Unconventional Protein Secretion by Saccharomyces cerevisiae and other Fungi. Cells 2018, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Cantu, A.; Ascencio, D.; Herrera-Basurto, S.; Xu, J.; Roguev, A.; Krogan, N.J.; DeLuna, A. Protein Moonlighting Revealed by Noncatalytic Phenotypes of Yeast Enzymes. Genetics 2018, 208, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Detomasi, T.C.; Rico-Ramirez, A.M.; Sayler, R.I.; Goncalves, A.P.; Marletta, M.A.; Glass, N.L. A moonlighting function of a chitin polysaccharide monooxygenase, CWR-1, in Neurospora crassa allorecognition. Elife 2022, 11, e80459. [Google Scholar] [CrossRef] [PubMed]
- Flores, C.L.; Arino, J.; Gancedo, C. The N-Acetylglucosamine Kinase from Yarrowia lipolytica Is a Moonlighting Protein. Int. J. Mol. Sci. 2021, 22, 13109. [Google Scholar] [CrossRef] [PubMed]
- Amblee, V.; Jeffery, C.J. Physical Features of Intracellular Proteins that Moonlight on the Cell Surface. PLoS ONE 2015, 10, e0130575. [Google Scholar] [CrossRef]
- Jeffery, C. The Use of Proteomics Studies in Identifying Moonlighting Proteins. Methods Mol. Biol. 2019, 1871, 437–443. [Google Scholar]
- Nunez-Beltran, A.; Lopez-Romero, E.; Cuellar-Cruz, M. Identification of proteins involved in the adhesionof Candida species to different medical devices. Microb. Pathog. 2017, 107, 293–303. [Google Scholar] [CrossRef]


{kind=link}
| Protein Name | One Function | Another Function | Reference Number |
|---|---|---|---|
| Interacting with DNA or affecting DNA metabolism | |||
| Emericella nidulans | |||
| I-AniI | mRNA maturase | Homing endonuclease | [3,4] |
| Kluyveromyces lactis | |||
| Galactokinase | Galactokinase | Transcriptional activator | [5,6] |
| Tuber melanosporum | |||
| Ask1 | Component of | Binds DNA, | [7] |
| DASH complex | transcriptional regulator | ||
| Phosphoadenosine | Thioredoxin | Transcription factor | [8] |
| phosphosulfate reductase | |||
| Saccharomyces cerevisiae | |||
| Acetohydroxyacid | Acetohydroxyacid | Maintains mitochondrial | [9] |
| reductoisomerase | reductoisomerase | DNA stability | |
| Arg5,6 | N-acetyl-gamma | Regulator of transcription | [10] |
| phosphate reductase/acetyl glutamate kinase | |||
| Arg82 | Kinase | Stabilizes transcription factors | [11] |
| ARG80 and MCM1 | |||
| Glycerol 3-phosphate | Glycerol 3-phosphate | Binds to Sir2 | [12] |
| Dehydrogenase | dehydrogenase | ||
| Hexokinase | Hexokinase | Binds Mig1 | [13,14] |
| Hsp60 | Chaperonin | Binds DNA | [15] |
| Lys20 | Homocitrate synthase | DNA damage repair | [16] |
| Superoxide dismutase | Superoxide dismutase | Binds DNA | [17,18] |
| Ure2 | functions in | Glutathione peroxidase, enzyme | [19,20,21,22] |
| nitrogen catabolite repression | |||
| Zuotin | Component of the | Activator of a transcription factor | [23] |
| RAC chaperone complex | |||
| Binding to RNA | |||
| Neurospora crassa | |||
| Tyrosyl tRNA synthetase | Catalyzes aminoacylation | Promotes folding of introns | [24] |
| of tyrosine | |||
| Saccharomyces cerevisiae | |||
| Isocitrate dehydrogenase 1 | Isocitrate dehydrogenase | Binds mRNA | [25] |
| Isocitrate dehydrogenase 2 | Isocitrate dehydrogenase | Binds mRNA | [25] |
| L2 Ribosomal protein | Component of ribosome | Shortens half-life of L2 mRNA | [26] |
| L32 Ribosomal protein | Component of ribosome | Inhibits the splicing of RNA | [27] |
| Leucyl-tRNA synthetase | Leucyl-tRNA synthetase | Intron splicing | [28] |
| S14 Ribosomal protein | Component of ribosome | Binds RNA, translation repressor | [29] |
| S28 Ribosomal protein | Component of ribosome | Binds, shortens half-life of mRNA | [30] |
| Protein–Protein Interactions | |||
| Hansenula poymorpha | |||
| Pyruvate carboxylase | Pyruvate carboxylase | Assembly of peroxisomal | [31,32] |
| alcohol oxidase | |||
| Pichia pastoris | |||
| Phosphofructokinase | Phosphofructokinase | Microautophagy | [33] |
| Saccharomyces cerevisiae | |||
| Arginase | Arginase | Binds and inactivates | [34] |
| ornithine transcarbamylase | |||
| Cytochrome C | Electron carrier protein | Binds apoptosis proteins | [35] |
| in electron transport chain | |||
| Enolase 1 | Enolase | Vacuole-membrane fusion | [36] |
| and protein trafficking to the vacuole | |||
| Enolase 2 | Enolase | Vacuole-membrane fusion | [36] |
| and protein trafficking to the vacuole | |||
| Fructose 1,6-bisphosphate | Aldolase | Binds vacuolar H+-ATPase | [37] |
| aldolase | and is needed for its assembly | ||
| Hal3 | Subunit of | Inhibitory subunit of | [38,39] |
| phosphopantothenoyl | protein phosphatase PPZ1 | ||
| cysteine decarboxylase | |||
| coenzyme A | |||
| Peroxiredoxin 1 | Peroxidase | Molecular chaperone | [40] |
| Peroxiredoxin 2 | Peroxidase | Chaperone | [40] |
| Rack1 | Component of ribosome | Scaffold in | [41,42] |
| signal transduction pathways | |||
| Succinate dehydrogenase | Electron transport in | Component of TIM22 complex | [43,44,45,46] |
| subunit 3 | respiratory complex II | ||
| Cell Surface | |||
| Candida albicans | |||
| 6-phosphogluconate | Oxidative decarboxylation | Binds components of contact system | [47,48,49] |
| dehydrogenase | of 6-phosphogluconate | ||
| Alcohol dehydrogenase 1 | Alcohol dehydrogenase | Binds plasminogen | [50] |
| Ef-Tu | Elongation factor | Binds plasminogen, | [48,49,51] |
| during protein synthesis | factor H, HK, FXII, and prekallikrein | ||
| Enolase | Enolase, enzyme | Binds plasminogen | [52] |
| Fructose bisphosphate | Fructose bisphosphate | Binds plasminogen | [50] |
| aldolase | aldolase | ||
| Glyceraldehyde | Glyceraldehyde | Binds plasminogen, | [53] |
| 3-phosphate | 3-phosphate | fibronectin, and laminin | |
| dehydrogenase | dehydrogenase | ||
| Glutathione reductase | Glutathione reductase | Binds plasma proteins | [54,55] |
| Glycerol 3-phosphate | Glycerol 3-phosphate | Binds plasminogen | [56] |
| dehydrogenase | dehydrogenase | ||
| Heat shock protein Ssa2 | Chaperone | Binds a peptide in saliva | [57] |
| High-affinity glucose | Glucose transporter | Complement inhibitor | [58] |
| transporter 1 | |||
| Integrin-like protein | Inhibits opsonization | Adhesin | [59,60] |
| and phagocytosis | |||
| Peroxisomal catalase | Peroxisomal catalase | Plasminogen binding | [50] |
| Phosphoglycerate kinase | Phosphoglycerate kinase | Plasminogen binding | [50] |
| Phosphoglyceromutase | Phosphoglyceromutase | Binds plasminogen, vitronectin, and fibronectin | [50,61] |
| Phosphotransferase | Transports and | Binds to plasma proteins | [48,62] |
| phosphorylates sugars | |||
| Superoxide dismutase 3 | Removes superoxide | Binds contact system proteins | [48,49] |
| radicals | HK, FXII, and prekallikrein | ||
| Thiol-specific antioxidant | Thiol-specific antioxidant | Binds plasminogen | [50] |
| protein | protein | ||
| Transcription elongation | Transcription elongation | Binds plasminogen | [50] |
| factor | factor | ||
| Triosephosphate | Isomerase | Binds extracellular matrix proteins | [61] |
| isomerase | |||
| Heat shock protein Ssa2 | Chaperone | Binds HTN3/histatin 5 | [57] |
| Candida glabrata | |||
| Triosephosphate | Isomerase | Binds extracellular matrix proteins | [61] |
| isomerase | |||
| Candida tropicalis | |||
| Enolase | Enolase | Binds vitronectin, fibronectin, | [63,64] |
| and plasminogen | |||
| Fructose-bisphosphate | Aldolase | Binds fibronectin, | [64] |
| aldolase | vitronectin, and laminin | ||
| Cryptococcus neoformans | |||
| Glucose 6-phosphate | Glucose 6-phosphate | Binds plasminogen | [65] |
| isomerase | isomerase | ||
| Nitric oxide dioxygenase | Nitric oxide dioxygenase | Binds plasminogen | [65] |
| Phosphoglycerate kinase | Phosphoglycerate kinase | Binds plasminogen | [65] |
| Pyruvate kinase | Pyruvate kinase | Binds plasminogen | [65] |
| Subunits of ATP synthase | ATP synthesis | Binds plasminogen | [65] |
| Histoplasma capsulatum | |||
| Hsp60 chaperone | Chaperone | Adhesin | [66] |
| Paracoccidioides brasiliensis | |||
| Enolase | Enolase | Binds fibronectin | [67] |
| Glyceraldehyde | Glycerol 3-phosphate | Binds fibronectin, laminin, | [68] |
| 3-phosphate | dehydrogenase | and type I collagen | |
| dehydrogenase | |||
| Triose phosphate | Triose phosphate | Adhesin | [69] |
| isomerase | isomerase | ||
| Sporothrix schenckii | |||
| GroEL/Hsp60 | Chaperone | Binds laminin, elastin, | [70] |
| fibrinogen, and fibronectin | |||
| Talaromyces marneffei | |||
| Glyceraldehyde | Glyceraldehyde | Adhesin | [71] |
| 3-phosphate | 3-phosphate | ||
| dehydrogenase | dehydrogenase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtis, N.J.; Patel, K.J.; Rizwan, A.; Jeffery, C.J. Moonlighting Proteins: Diverse Functions Found in Fungi. J. Fungi 2023, 9, 1107. https://doi.org/10.3390/jof9111107
Curtis NJ, Patel KJ, Rizwan A, Jeffery CJ. Moonlighting Proteins: Diverse Functions Found in Fungi. Journal of Fungi. 2023; 9(11):1107. https://doi.org/10.3390/jof9111107
Chicago/Turabian StyleCurtis, Nicole J., Krupa J. Patel, Amina Rizwan, and Constance J. Jeffery. 2023. "Moonlighting Proteins: Diverse Functions Found in Fungi" Journal of Fungi 9, no. 11: 1107. https://doi.org/10.3390/jof9111107