
Dispersal Limitation Controlling the Assembly of the Fungal Community in Karst Caves


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Processing and Data Processing
2.2. Statistical Analyses
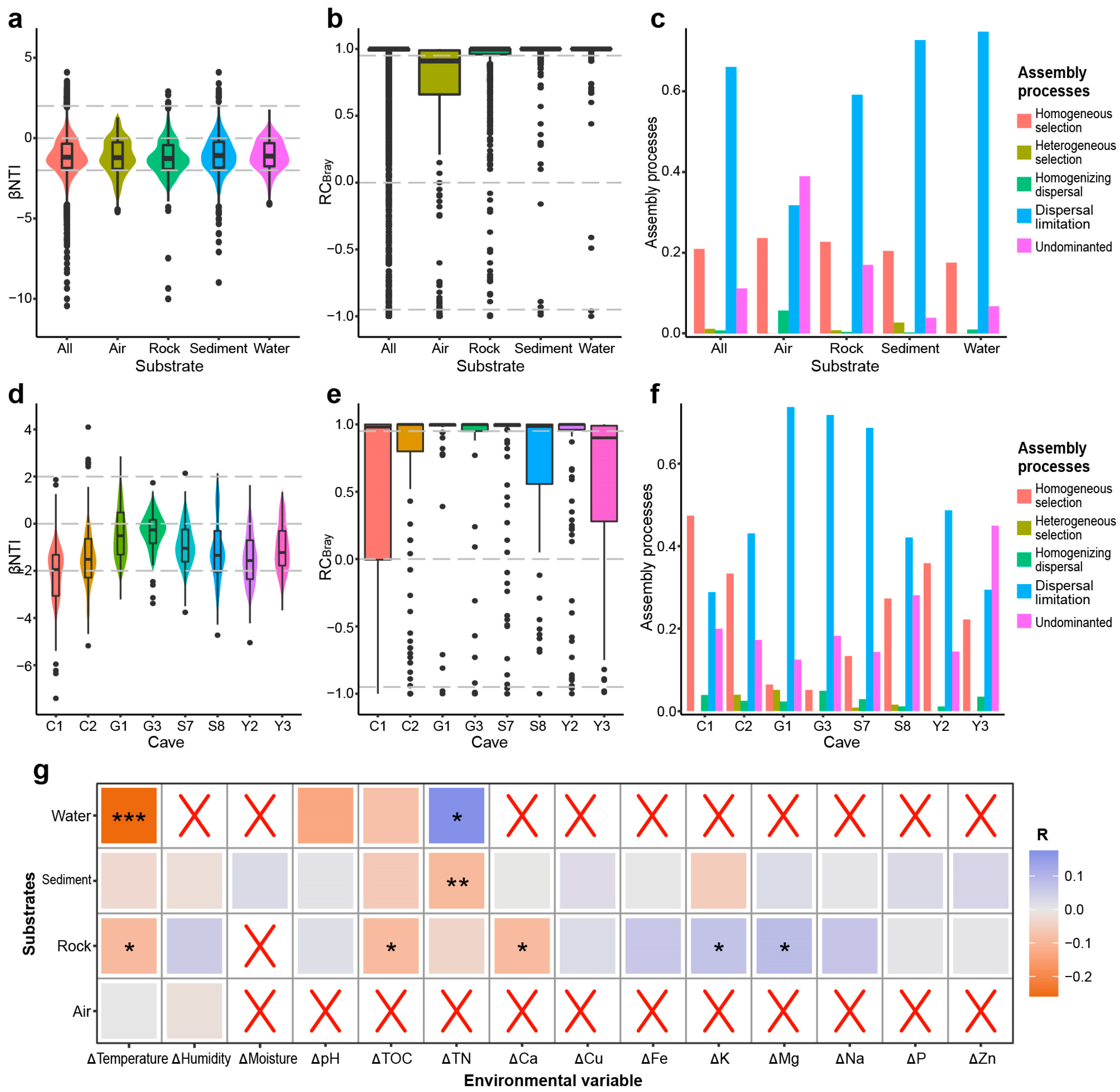
Fungal Community Assembly Patterns
3. Results
3.1. Dispersal Limitation Plays a Crucial Role in Shaping the Overall Fungal Community in Karst Caves
3.2. Environmental Selection Plays a Minor Role in the Fungal Community in Karst Caves
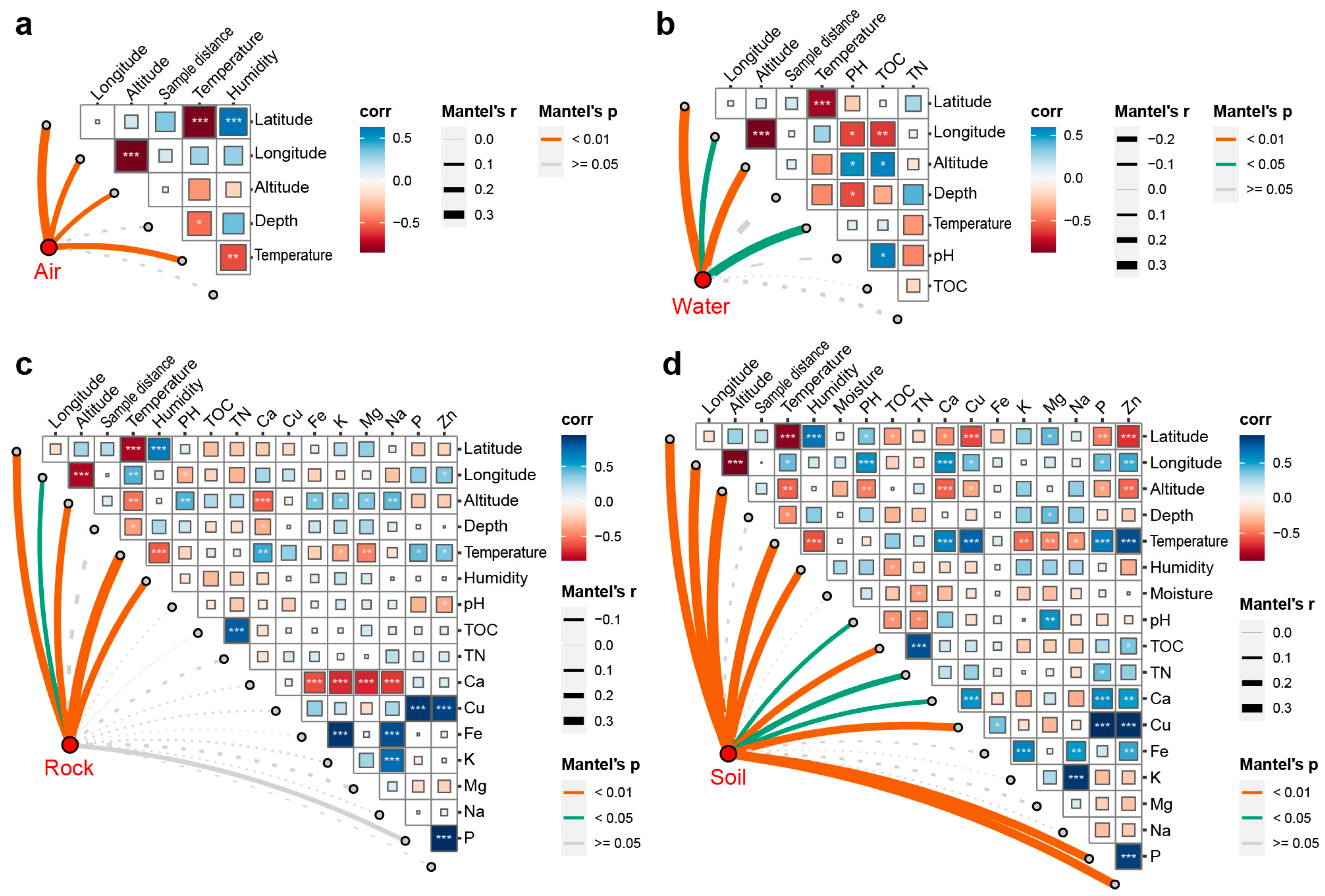
4. Discussion
4.1. Geographic Barriers between Caves May Lead to the Dispersal Limitation of the Overall Fungal Community in Karst Caves
4.2. Environmental and Spatial Factors Have a Relatively Weak Impact on the Fungal Communities in Karst Caves
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Ning, D.L. Stochastic Community Assembly: Does It Matter in Microbial Ecology. Microbiol. Mol. Biol. R 2017, 81, e00002–e00017. [Google Scholar] [CrossRef]
- Chen, W.; Ren, K.; Isabwe, A.; Chen, H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2019, 7, 138. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Pan, J.; Pan, Y.P.; Li, M. Biogeography, Assembly Patterns, Driving Factors, and Interactions of Archaeal Community in Mangrove Sediments. mSystems 2021, 6, e0138120. [Google Scholar] [CrossRef] [PubMed]
- Cheeptham, N. Cave Microbiomes: A Novel Resource for Drug Discovery; Springer: New York, NY, USA, 2012. [Google Scholar]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Northup, E.; Lavoie Diana, L.H. Geomicrobiology of Caves: A Review. Geomicrobiol. J. 2010, 18, 199–222. [Google Scholar] [CrossRef]
- Vanderwolf, K.; Malloch, D.; McAlpine, D.; Forbes, G. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Rawat, S.; Rautela, R.; Johri, B.N. Fungal world of cave eco-system. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S.K., Johri, B.N., Eds.; Springer: Singapore, 2017; pp. 99–124. [Google Scholar]
- Carmichael, S.K.; Zorn, B.T.; Santelli, C.M.; Roble, L.A.; Carmichael, M.J.; Brauer, S.L. Nutrient input influences fungal community composition and size and can stimulate manganese (II) oxidation in caves. Environ. Microbiol. Rep. 2015, 7, 592–605. [Google Scholar] [CrossRef]
- Zhang, Z.-F.; Zhou, S.-Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable mycobiota from Karst caves in China II, with descriptions of 33 new species. Fungal Divers. 2020, 106, 29–136. [Google Scholar] [CrossRef]
- Preedanon, S.; Suetrong, S.; Shrihom, S.; Somrithipol, S.; Kobmoo, N.; Saengkaewsuk, S.; Srikitikulchai, P.; Klaysuban, A.; Nuankaew, S.; Chuaseeharonnachai, C.; et al. Eight novel cave fungi in Thailand’s Satun Geopark. Fungal Syst. Evol. 2023, 12, 1–30. [Google Scholar] [CrossRef]
- Ogórek, R.; Pusz, W.; Matkowski, K.; Pląskowska, E. Assessment of Abundance and Species Composition of Filamentous Fungi in the Underground Rzeczka Complex in Sowie Mountains (Lower Silesia, Poland). Geomicrobiol. J. 2014, 31, 900–906. [Google Scholar] [CrossRef]
- Ogórek, R.; Pusz, W.; Zagożdżon, P.P.; Kozak, B.; Bujak, H. Abundance and Diversity of Psychrotolerant Cultivable Mycobiota in Winter of a Former Aluminous Shale Mine. Geomicrobiol. J. 2017, 34, 823–833. [Google Scholar] [CrossRef]
- Pusz, W.; Ogórek, R.; Uklańska-Pusz, C.; Zagożdżon, P. Speleomycological research in underground Osówka complex in Sowie Mountains (Lower Silesia, Poland). Int. J. Speleol. 2014, 43, 27–34. [Google Scholar] [CrossRef]
- Vaughan-Martini, A.; Angelini, P.; Zacchi, L. The influence of human and animal visitation on the yeast ecology of three Italian caverns. Ann. Microbiol. 2000, 50, 133–140. [Google Scholar]
- Kuzmina, L.Y.; Galimzianova, N.F.; Abdullin, S.R.; Ryabova, A.S. Microbiota of the Kinderlinskaya cave (South Urals, Russia). Microbiology 2012, 81, 251–258. [Google Scholar] [CrossRef]
- Cloutier, M.L.C.; Carmichael, S.K.; Carson, M.A.; Madritch, M.D.; Bräuer, S.L. Carbon quantity and quality drives variation in cave microbial communities and regulates Mn(II) oxidation. Biogeochemistry 2017, 134, 77–94. [Google Scholar] [CrossRef]
- Pfendler, S.; Karimi, B.; Maron, P.A.; Ciadamidaro, L.; Valot, B.; Bousta, F.; Alaoui-Sosse, L.; Alaoui-Sosse, B.; Aleya, L. Biofilm biodiversity in French and Swiss show caves using the metabarcoding approach: First data. Sci. Total Environ. 2018, 615, 1207–1217. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Cai, L. Substrate and spatial variables are major determinants of fungal community in karst caves in Southwest China. J. Biogeogr. 2019, 46, 1504–1518. [Google Scholar] [CrossRef]
- Ogórek, R.; Pusz, W.; Lejman, A.; Uklańska-Pusz, C. Microclimate Effects on Number and Distribution of Fungi in the Włodarz Underground Complex in the Owl Mountains (Góry Sowie), Poland. J. Cave Karst Stud. 2014, 76, 146–153. [Google Scholar] [CrossRef]
- Pusz, W.; Baturo-Cieśniewska, A.; Zagożdżon, P.; Ogórek, R. Mycobiota of the disused ore mine of Marcinków in Śnieżnik Masiff (western Poland). J. Mt. Sci. 2017, 14, 2448–2457. [Google Scholar] [CrossRef]
- Kokurewicz, T.; Ogorek, R.; Pusz, W.; Matkowski, K. Bats Increase the Number of Cultivable Airborne Fungi in the “Nietoperek” Bat Reserve in Western Poland. Microb. Ecol. 2016, 72, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Borzęcka, J.; Piecuch, A.; Kokurewicz, T.; Lavoie, K.H.; Ogórek, R. Greater Mouse-Eared Bats (Myotis myotis) Hibernating in the Nietoperek Bat Reserve (Poland) as a Vector of Airborne Culturable Fungi. Biology 2021, 10, 593. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Mills, D.A. Improved selection of internal transcribed spacer-specific primers enables quantitative, ultra-high-throughput profiling of fungal communities. Appl. Environ. Microbiol. 2013, 79, 2519–2526. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y. Abundant fungi adapt to broader environmental gradients than rare fungi in agricultural fields. Glob. Chang. Biol. 2020, 26, 4506–4520. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.M.; Stegen, J.C.; Kim, M.; Dong, K.; Adams, J.M.; Lee, Y.K. Soil pH mediates the balance between stochastic and deterministic assembly of bacteria. ISME J. 2018, 12, 1072–1083. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J. Geosphere: Spherical Trigonometry, R Package Version 1.5-10; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H. Vegan: Community Ecology Package, R Package Version 2.5-6; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Ning, D.; Yuan, M.; Wu, L.; Zhang, Y.; Guo, X.; Zhou, X.; Yang, Y.; Arkin, A.P.; Firestone, M.K.; Zhou, J. A quantitative framework reveals ecological drivers of grassland microbial community assembly in response to warming. Nat. Commun. 2020, 11, 4717. [Google Scholar] [CrossRef]
- Stegen, J.C.; Fredrickson, J.K.; Wilkins, M.J.; Konopka, A.E.; Nelson, W.C.; Arntzen, E.V.; Chrisler, W.B.; Chu, R.K.; Danczak, R.E.; Fansler, S.J.; et al. Groundwater-surface water mixing shifts ecological assembly processes and stimulates organic carbon turnover. Nat. Commun. 2016, 7, 11237. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, M.; Duan, C.; Cao, N.; Jia, W.; Zhao, Z.; Ding, C.; Huang, Y.; Wang, J. Contribution of stochastic processes to the microbial community assembly on field-collected microplastics. Environ. Microbiol. 2021, 23, 6707–6720. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Deng, Y.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.D.; Yang, Y.; He, Z.; Wu, L.; Stahl, D.A.; et al. Stochasticity, succession, and environmental perturbations in a fluidic ecosystem. Proc. Natl. Acad. Sci. USA 2014, 111, E836–E845. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef]
- Bontemps, Z.; Alonso, L.; Pommier, T.; Hugoni, M.; Moenne-Loccoz, Y. Microbial ecology of tourist Paleolithic caves. Sci. Total Environ. 2022, 816, 151492. [Google Scholar] [CrossRef]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeterior. Biodegrad. 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhao, P.; Cai, L. Origin of Cave Fungi. Front. Microbiol. 2018, 9, 1407. [Google Scholar] [CrossRef] [PubMed]
- De Waele, J.; Follesa, R. Human impact on karst- the example of Lusaka (Zambia). Int. J. Speleol. 2003, 32, 71–83. [Google Scholar] [CrossRef]
- Jacobs, A.; Msimang, D.; Venter, E. First survey of the fungi from the Bakwena Cave in South Africa suggests low human disturbance. J. Cave Karst Stud. 2017, 79, 89–94. [Google Scholar] [CrossRef]
- Mammola, S.; Di Piazza, S.; Ziotti, M.; Badino, G.; Marco, I. Human-induced Alterations of the Mycobiota in an Alpine Show Cave (Italy, SW-Alps). Acta Carsologica 2017, 46, 111–123. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Lima-Mendez, G.; Faust, K.; Henry, N.; Decelle, J.; Colin, S.; Carcillo, F.; Chaffron, S.; Ignacio-Espinosa, J.C.; Roux, S.; Vincent, F.; et al. Determinants of community structure in the global plankton interactome. Science 2015, 348, 1262073. [Google Scholar] [CrossRef]
- Wei, G.; Li, M.; Li, F.; Li, H.; Gao, Z. Distinct distribution patterns of prokaryotes between sediment and water in the Yellow River estuary. Appl. Microbiol. Biotechnol. 2016, 100, 9683–9697. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-F.; Mao, J.; Cai, L. Dispersal Limitation Controlling the Assembly of the Fungal Community in Karst Caves. J. Fungi 2023, 9, 1013. https://doi.org/10.3390/jof9101013
Zhang Z-F, Mao J, Cai L. Dispersal Limitation Controlling the Assembly of the Fungal Community in Karst Caves. Journal of Fungi. 2023; 9(10):1013. https://doi.org/10.3390/jof9101013
Chicago/Turabian StyleZhang, Zhi-Feng, Jian Mao, and Lei Cai. 2023. "Dispersal Limitation Controlling the Assembly of the Fungal Community in Karst Caves" Journal of Fungi 9, no. 10: 1013. https://doi.org/10.3390/jof9101013