
Proteomic Response of Paracoccidioides brasiliensis Exposed to the Antifungal 4-Methoxynaphthalene-N-acylhydrazone Reveals Alteration in Metabolism
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis of AOS
2.2. Cultivation Conditions of Paracoccidioides spp.
2.3. Determination of Inhibitory and Fungicidal Concentration
2.4. Cytotoxicity Assay
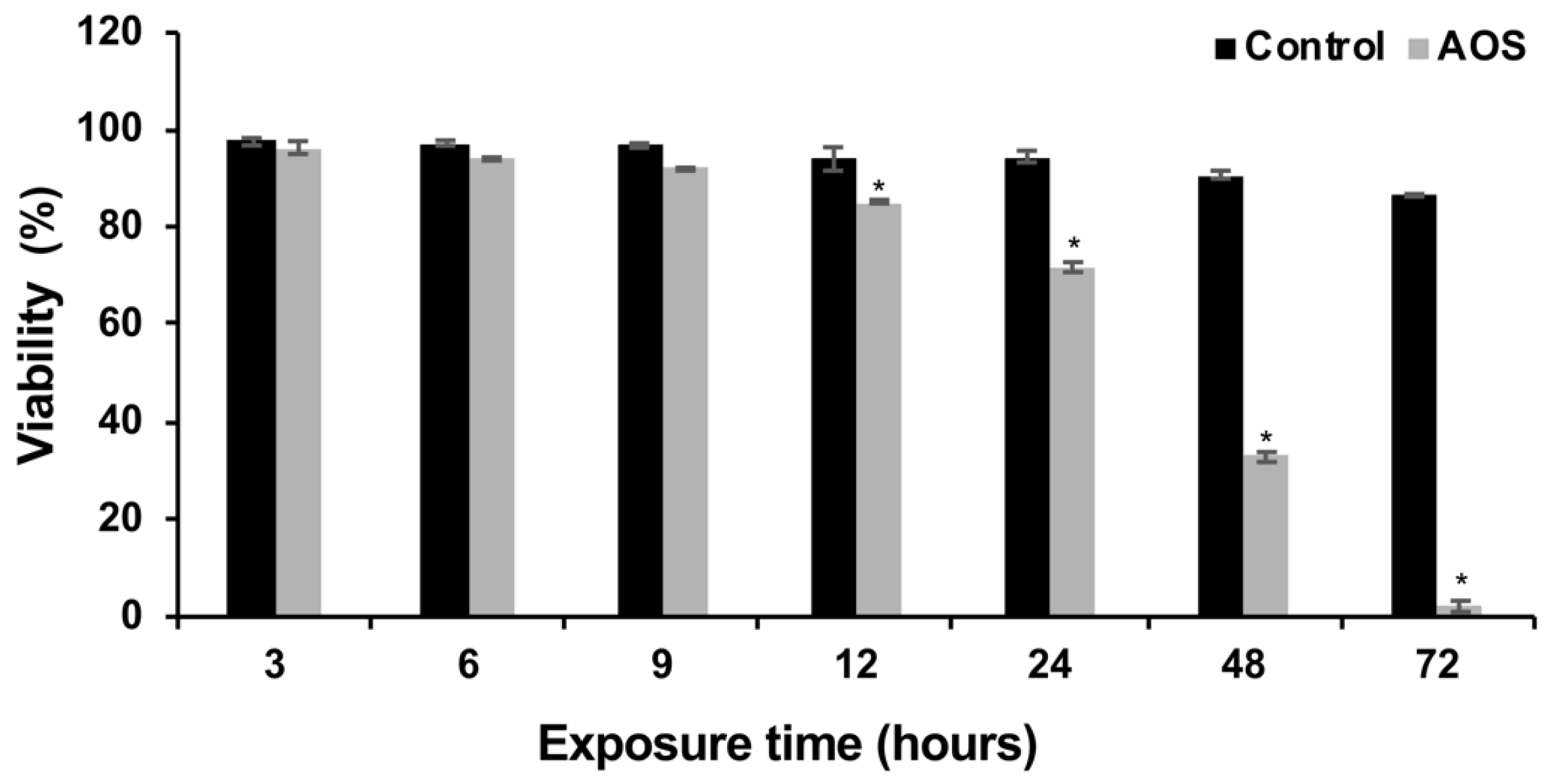
2.5. Temporal Cell Viability
2.6. Proteome of P. brasiliensis in the Presence of AOS
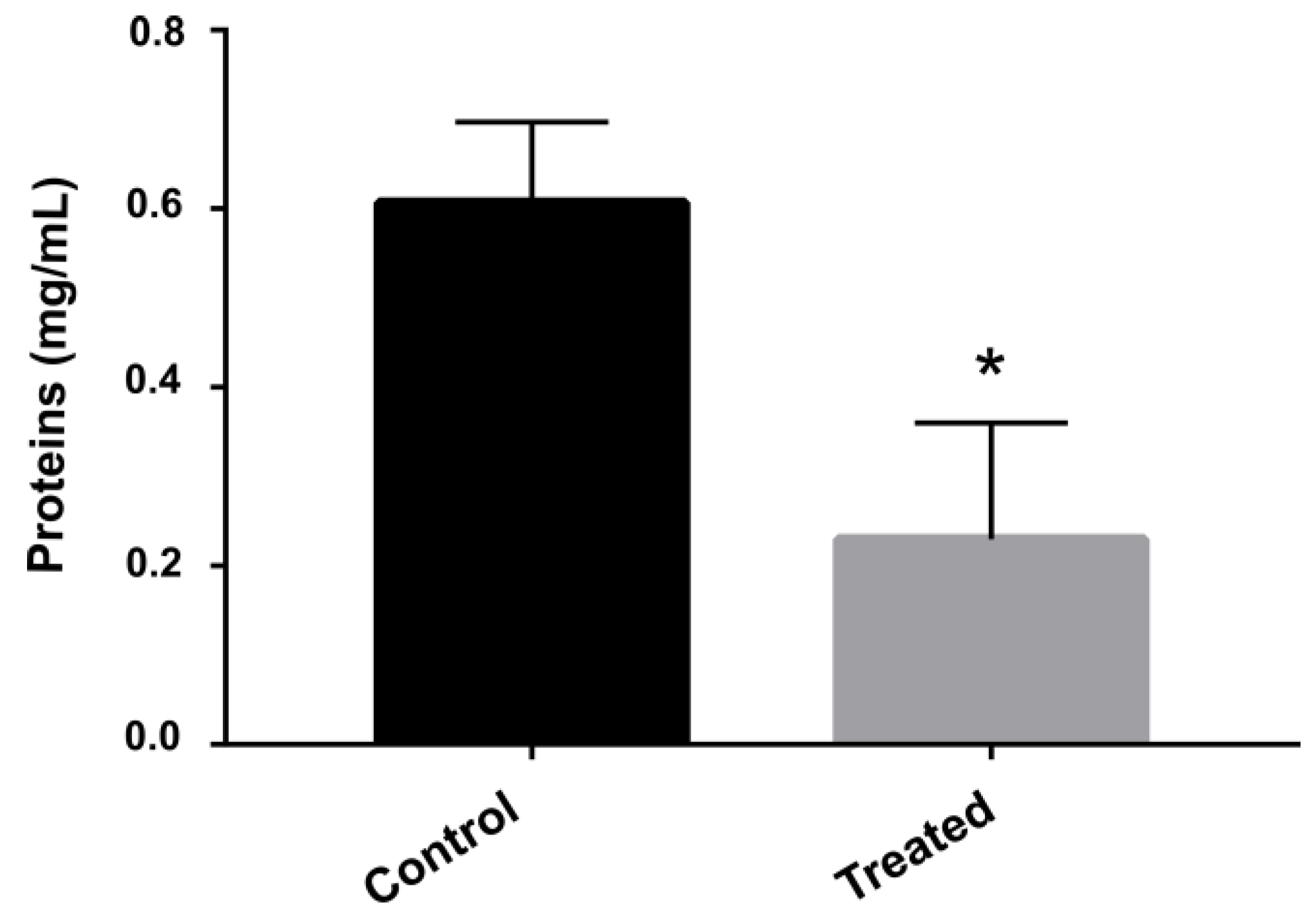
2.7. Protein Dosage
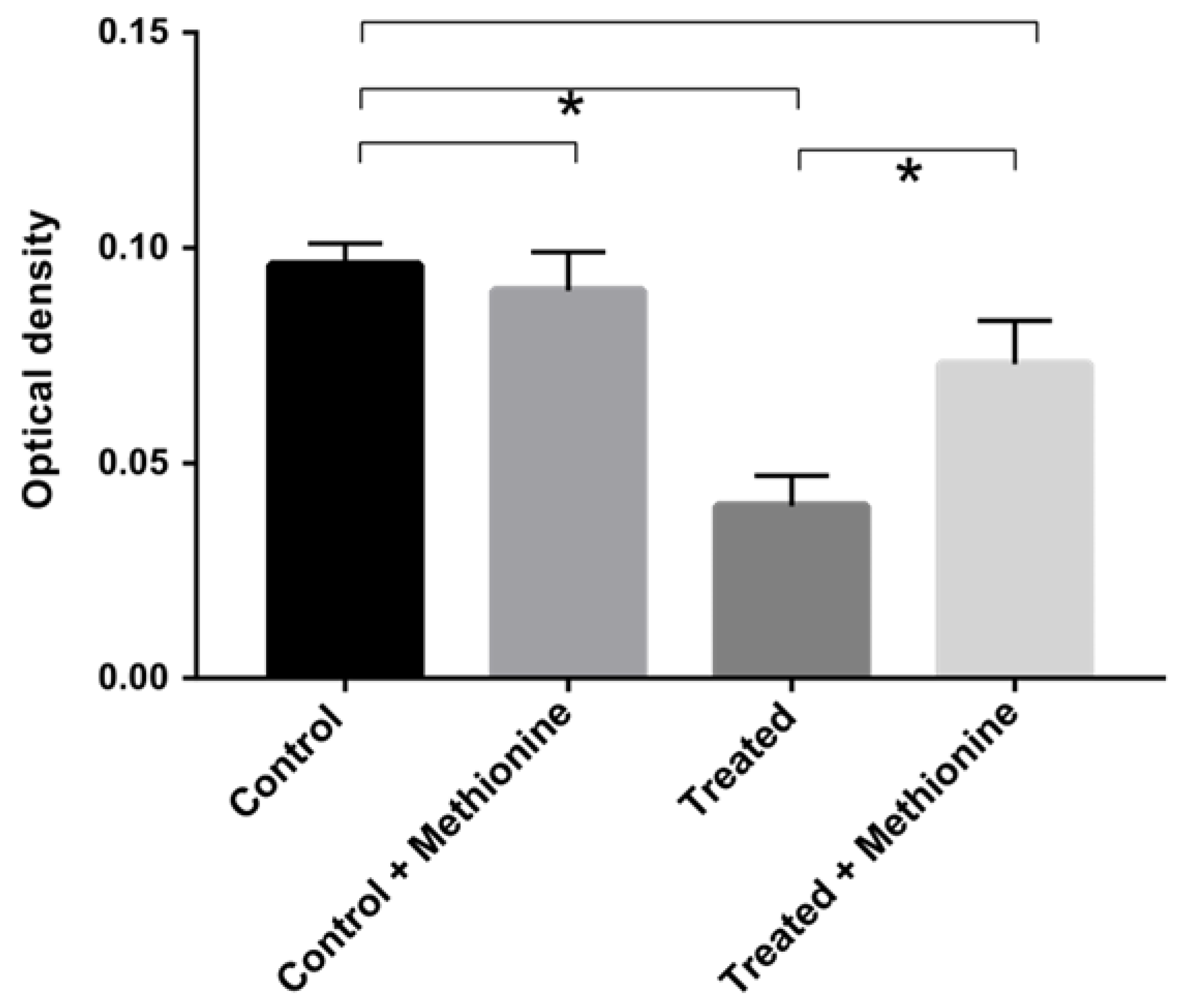
2.8. Growth Assay with Methionine Supplementation
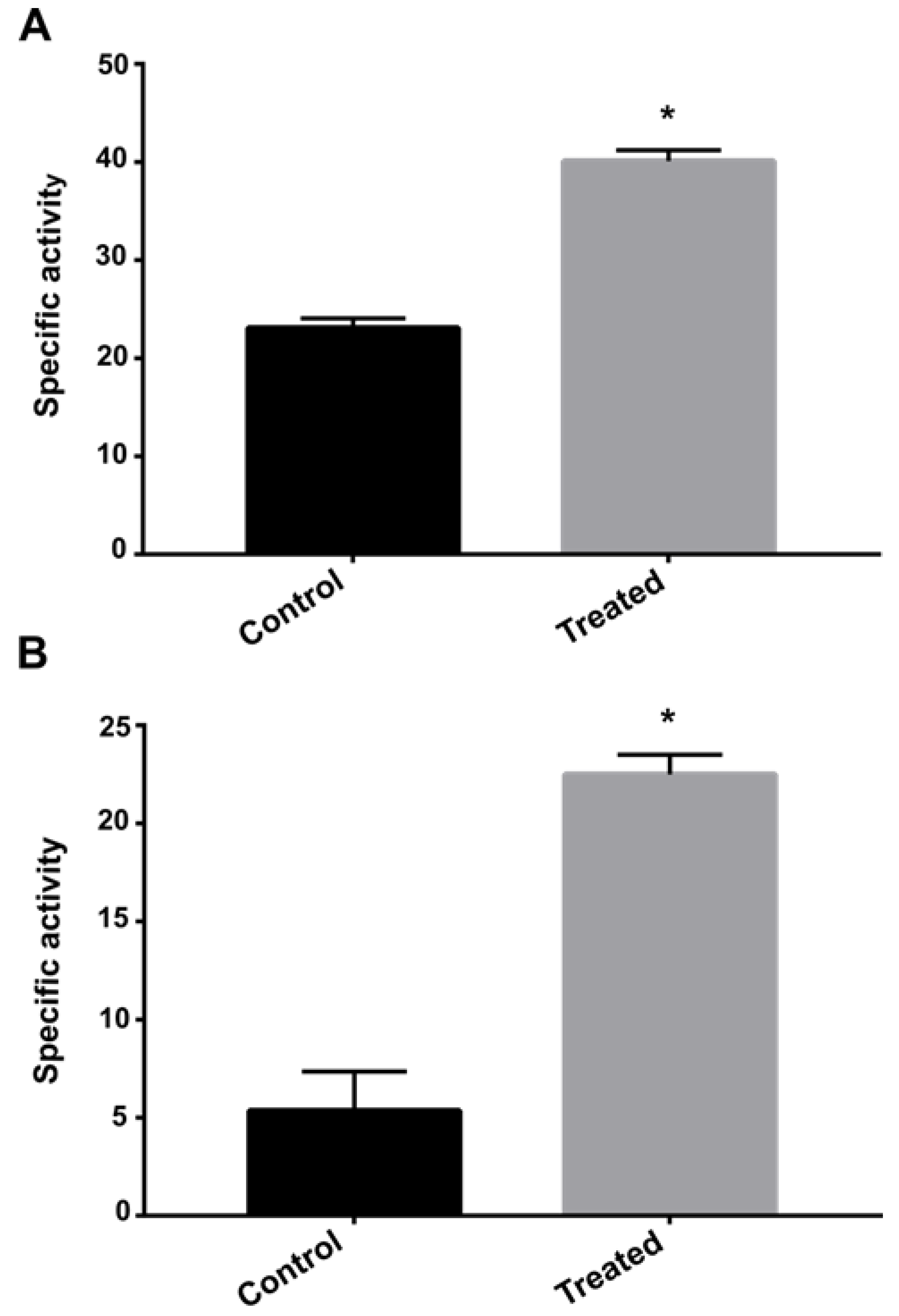
2.9. Enzyme Activity Inhibition Assay
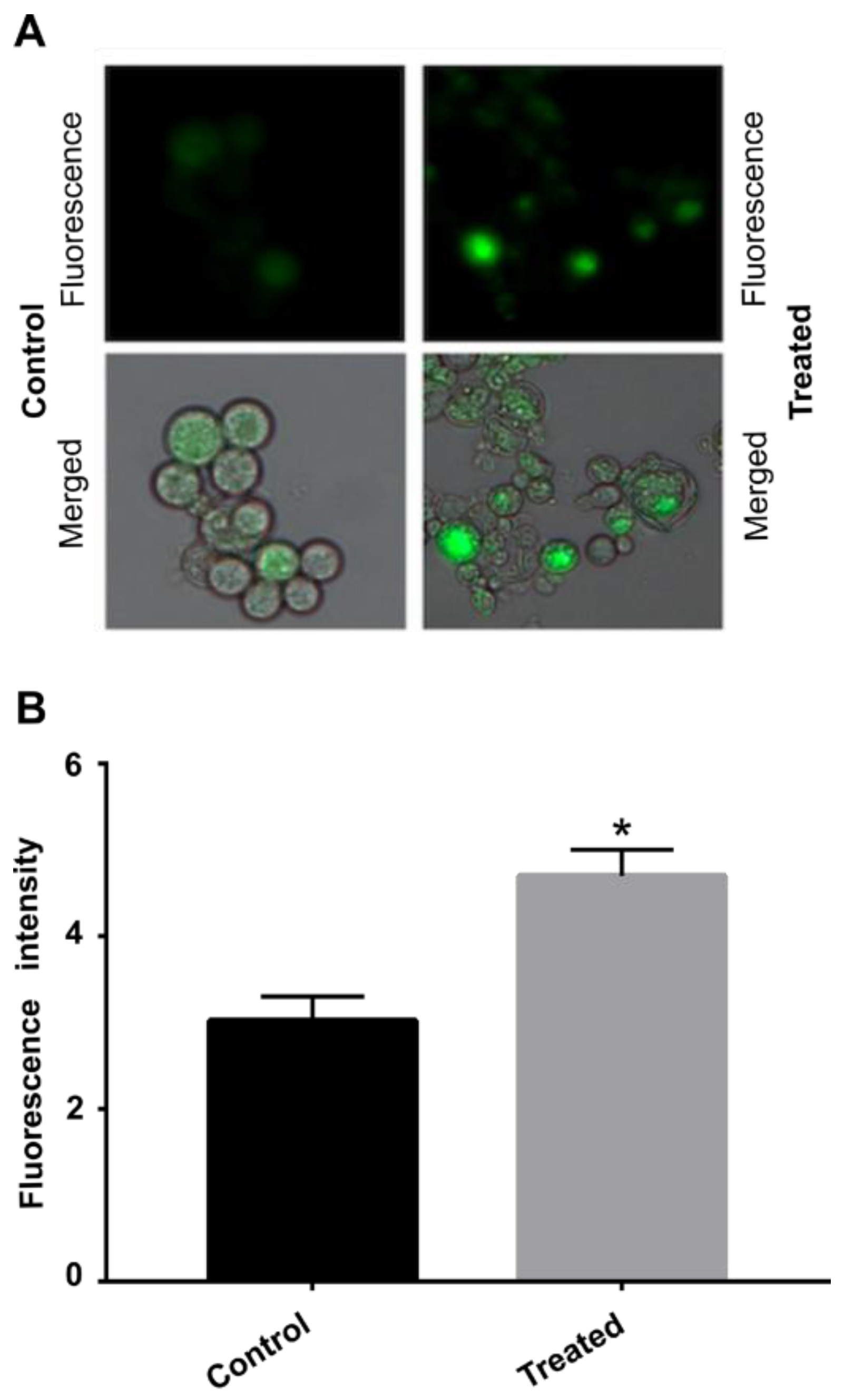
2.10. Quantification Assay of Reactive Oxygen Species
3. Results
3.1. Antifungal and Cytotoxic Activity of AOS
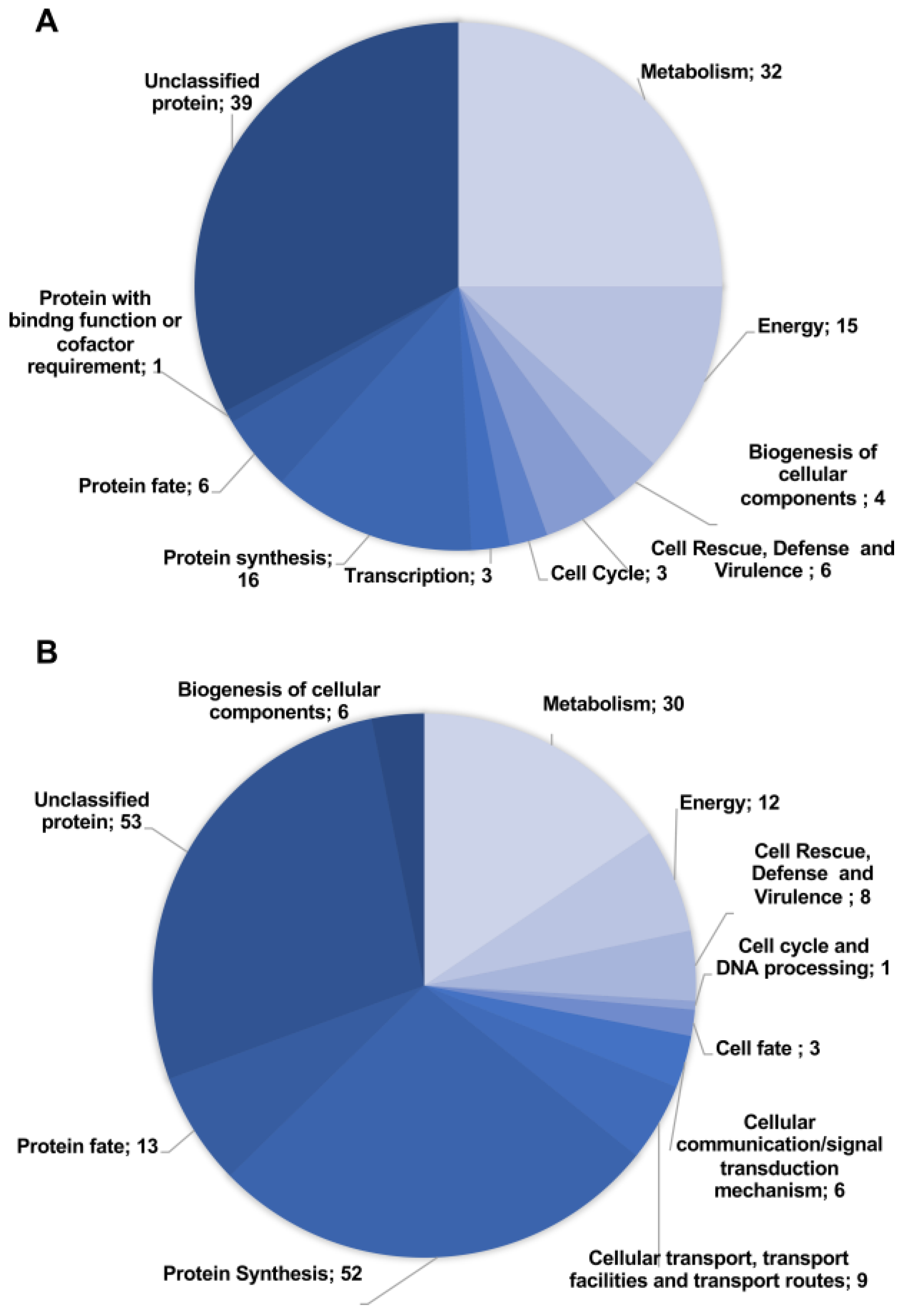
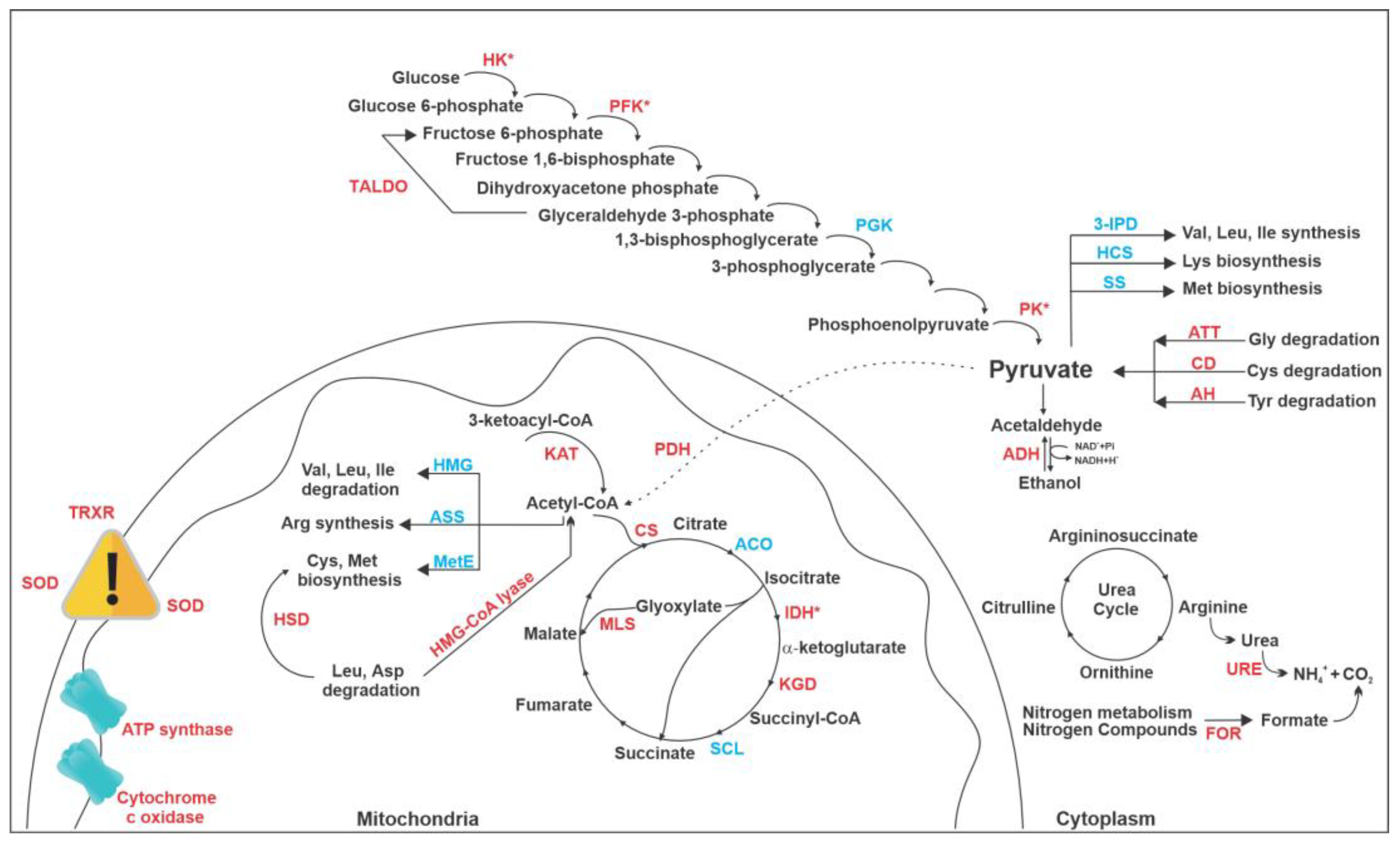
3.2. Proteomic Response of P. brasiliensis to AOS
3.3. Exposure to AOS Inhibits Protein Synthesis
3.4. Methionine Supplementation Restores Fungal Growth Exposed to AOS
3.5. Exposure to AOS Induces Nitrogen Metabolism
3.6. AOS Causes Oxidative Stress in Cells
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian Guidelines for the Clinical Management of Paracoccidioidomycosis. Rev. Da Soc. Bras. De Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, F.A.; Neves, F.F.; Mora, D.J.; Reis, T.A.D.; Sotini, D.M.; Ribeiro, B.D.M.; Andrade-Silva, L.E.; Nascentes, G.N.; Ferreira-Paim, K.; Silva-Vergara, M.L. Paracoccidioidomycosis in Brazilian Patients With and Without Human Immunodeficiency Virus Infection. Am. J. Trop. Med. Hyg. 2017, 96, 368–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, J., Jr.; Peçanha-Pietrobom, P.; Colombo, A. Paracoccidioidomycosis in Immunocompromised Patients: A Literature Review. J. Fungi 2018, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai-Yasuda, M.A. Paracoccidioidomycosis Treatment. Rev. Do Inst. De Med. Trop. De São Paulo 2015, 57, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.A.; Rice, L.B. Antifungal Agents: Mode of Action, Mechanisms of Resistance, and Correlation of These Mechanisms with Bacterial Resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [Green Version]
- do Carmo Silva, L.; de Oliveira, A.A.; de Souza, D.R.; Barbosa, K.L.B.; Freitas e Silva, K.S.; Carvalho Júnior, M.A.B.; Rocha, O.B.; Lima, R.M.; Santos, T.G.; Soares, C.M.d.A.; et al. Overview of Antifungal Drugs against Paracoccidioidomycosis: How Do We Start, Where Are We, and Where Are We Going? J. Fungi 2020, 6, 300. [Google Scholar] [CrossRef]
- Lima, R.M.; Freitas e Silva, K.S.; Silva, L.d.C.; Ribeiro, J.F.R.; Neves, B.J.; Brock, M.; Soares, C.M.d.A.; da Silva, R.A.; Pereira, M. A Structure-Based Approach for the Discovery of Inhibitors against Methylcitrate Synthase of Paracoccidioides Lutzii. J. Biomol. Struct. Dyn. 2021, 40, 9361–9373. [Google Scholar] [CrossRef]
- da Silva, L.S.; Barbosa, U.R.; Silva, L.d.C.; Soares, C.M.; Pereira, M.; da Silva, R.A. Identification of a New Antifungal Compound against Isocitrate Lyase of Paracoccidioides brasiliensis. Future Microbiol. 2019, 14, 1589–1606. [Google Scholar] [CrossRef]
- Costa, F.G.; Neto, B.R.d.S.; Gonçalves, R.L.; da Silva, R.A.; de Oliveira, C.M.A.; Kato, L.; Freitas, C.d.S.; Giannini, M.J.S.M.; da Silva, J.d.F.; Soares, C.M.d.A.; et al. Alkaloids as Inhibitors of Malate Synthase from Paracoccidioides Spp.: Receptor-Ligand Interaction-Based Virtual Screening and Molecular Docking Studies, Antifungal Activity, and the Adhesion Process. Antimicrob. Agents Chemother. 2015, 59, 5581–5594. [Google Scholar] [CrossRef] [Green Version]
- Bagatin, M.C.; Pimentel, A.L.; Biavatti, D.C.; Basso, E.A.; Kioshima, E.S.; Seixas, F.A.V.; Gauze, G.d.F. Targeting the Homoserine Dehydrogenase of Paracoccidioides Species for Treatment of Systemic Fungal Infections. Antimicrob. Agents Chemother. 2017, 61, e00165-17. [Google Scholar] [CrossRef]
- Bagatin, M.C.; FRozada, A.M.; VRodrigues, F.A.; ABueno, P.S.; Santos, J.L.; Canduri, F.; Kioshima, É.S.; VSeixas, F.A.; Basso, E.A.; Gauze, G.F. New 4-Methoxy-Naphthalene Derivatives as Promisor Antifungal Agents for Paracoccidioidomycosis Treatment. Future Microbiol. 2019, 14, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Bueno, P.S.A.; Rodrigues, F.A.V.; Santos, J.L.; Canduri, F.; Biavatti, D.C.; Pimentel, A.L.; Bagatin, M.C.; Kioshima, É.S.; de Freitas Gauze, G.; Seixas, F.A.V. New Inhibitors of Homoserine Dehydrogenase from Paracoccidioides brasiliensis Presenting Antifungal Activity. J. Mol. Model. 2019, 25, 325. [Google Scholar] [CrossRef]
- Rozada, A.M.; Rodrigues, F.A.; Sampiron, E.G.; Seixas, F.A.; Basso, E.A.; Scodro, R.B.; Kioshima, É.S.; Gauze, G.F. Novel 4-Methoxynaphthalene-N-Acylhydrazones as Potential for Paracoccidioidomycosis and Tuberculosis Co-Infection. Future Microbiol. 2019, 14, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Thota, S.; Rodrigues, D.A.; Pinheiro, P.d.S.M.; Lima, L.M.; Fraga, C.A.M.; Barreiro, E.J. N-Acylhydrazones as Drugs. Bioorganic Med. Chem. Lett. 2018, 28, 2797–2806. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Jiménez, B.E. Growth of Paracoccidioides brasiliensis Yeast Phase in a Chemically Defined Culture Medium. J. Clin. Microbiol. 1980, 12, 279–281. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.C.; Neves, B.J.; Gomes, M.N.; Melo-Filho, C.C.; Soares, C.M.; Andrade, C.H.; Pereira, M. Computer-Aided Identification of Novel Anti-Paracoccidioidomycosis Compounds. Future Microbiol. 2018, 13, 1523–1535. [Google Scholar] [CrossRef]
- Rocha, O.B.; Freitas e Silva, K.S.; de Carvalho Junior, M.A.B.; Moraes, D.; Alonso, A.; Alonso, L.; do Carmo Silva, L.; Soares, C.M.A.; Pereira, M. Proteomic Alterations in Paracoccidioides brasiliensis Caused by Exposure to Curcumin. J. Proteom. 2022, 266, 104683. [Google Scholar] [CrossRef]
- Murad, A.M.; Souza, G.H.M.F.; Garcia, J.S.; Rech, E.L. Detection and Expression Analysis of Recombinant Proteins in Plant-Derived Complex Mixtures Using NanoUPLC-MSE. J. Sep. Sci. 2011, 34, 2618–2630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Skouloubris, S.; Labigne, A.; De Reuse, H. Identification and Characterization of an Aliphatic Amidase in Helicobacter pylori. Mol. Microbiol. 1997, 25, 989–998. [Google Scholar] [CrossRef]
- Lerm, B.; Kenyon, C.; Schwartz, I.S.; Kroukamp, H.; de Witt, R.; Govender, N.P.; de Hoog, G.S.; Botha, A. First Report of Urease Activity in the Novel Systemic Fungal Pathogen Emergomyces africanus: A Comparison with the Neurotrope Cryptococcus neoformans. FEMS Yeast Res. 2017, 17, fox069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, O.B.; do Carmo Silva, L.; de Carvalho Júnior, M.A.B.; de Oliveira, A.A.; de Almeida Soares, C.M.; Pereira, M. In Vitro and in Silico Analysis Reveals Antifungal Activity and Potential Targets of Curcumin on Paracoccidioides spp. Braz. J. Microbiol. 2021, 52, 1897–1911. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.-S.; Shao, Y.-J.; Yang, Z.-T.; Liu, J.-X. Quantitative Proteomic Analysis of ER Stress Response Reveals Both Common and Specific Features in Two Contrasting Ecotypes of Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 9741. [Google Scholar] [CrossRef] [PubMed]
- Saul, M.J.; Hegewald, A.B.; Emmerich, A.C.; Ossipova, E.; Vogel, M.; Baumann, I.; Kultima, K.; Lengqivst, J.; Steinhilber, D.; Jakobsson, P.J. Mass Spectrometry-Based Proteomics Approach Characterizes the Dual Functionality of MiR-328 in Monocytes. Front. Pharmacol. 2019, 10, 640. [Google Scholar] [CrossRef] [Green Version]
- Houšť, J.; Spížek, J.; Havlíček, V. Antifungal Drugs. Metabolites 2020, 10, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, P.-W.; Chien, C.-Y.; Yeh, Y.-C.; Tung, L.; Chen, H.-F.; Chang, T.-H.; Lan, C.-Y. Candida Albicans Hom6 Is a Homoserine Dehydrogenase Involved in Protein Synthesis and Cell Adhesion. J. Microbiol. Immunol. Infect. 2017, 50, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.; Barbey, R.; Surdin-Kerjan, Y. Evolutionary Relationships between Yeast and Bacterial Homoserine Dehydrogenases. FEBS Lett. 1993, 323, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Prado, R.S.; Bailão, A.M.; Silva, L.C.; de Oliveira, C.M.A.; Marques, M.F.; Silva, L.P.; Silveira-Lacerda, E.P.; Lima, A.P.; Soares, C.M.; Pereira, M. Proteomic Profile Response of Paracoccidioides lutzii to the Antifungal Argentilactone. Front. Microbiol. 2015, 6, 616. [Google Scholar] [CrossRef]
- e Silva, K.S.; da S Neto, B.R.; Zambuzzi-Carvalho, P.F.; de Oliveira, C.M.; Pires, L.B.; Kato, L.; Bailão, A.M.; Parente-Rocha, J.A.; Hernández, O.; Ochoa, J.G.; et al. Response of Paracoccidioides lutzii to the Antifungal Camphene Thiosemicarbazide Determined by Proteomic Analysis. Future Microbiol. 2018, 13, 1473–1496. [Google Scholar] [CrossRef]
- Klausmeyer, P.; McCloud, T.G.; Tucker, K.D.; Cardellina, J.H.; Shoemaker, R.H. Aspirochlorine Class Compounds from Aspergillus flavus Nhibit Azole-Resistant Candida albicans. J. Nat. Prod. 2005, 68, 1300–1302. [Google Scholar] [CrossRef]
- Sun, N.; Li, D.; Zhang, Y.; Killeen, K.; Groutas, W.; Calderone, R. Repurposing an Inhibitor of Ribosomal Biogenesis with Broad Anti-Fungal Activity. Sci. Rep. 2017, 7, 17014. [Google Scholar] [CrossRef] [PubMed]
- Jastrzębowska, K.; Gabriel, I. Inhibitors of Amino Acids Biosynthesis as Antifungal Agents. Amino Acids 2015, 47, 227–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walvekar, A.S.; Laxman, S. Methionine at the Heart of Anabolism and Signaling: Perspectives from Budding Yeast. Front. Microbiol. 2019, 10, 2624. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.; Sueiro-Olivares, M.; Thornton, B.P.; Owens, R.A.; Muhamadali, H.; Fortune-Grant, R.; Thomson, D.; Thomas, R.; Hollywood, K.; Doyle, S.; et al. Targeting Methionine Synthase in a Fungal Pathogen Causes a Metabolic Imbalance That Impacts Cell Energetics, Growth, and Virulence. MBio 2020, 11, e01985-20. [Google Scholar] [CrossRef]
- Gerke, J.; Bayram, Ö.; Braus, G.H. Fungal S-Adenosylmethionine Synthetase and the Control of Development and Secondary Metabolism in Aspergillus nidulans. Fungal Genet. Biol. 2012, 49, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Karkowska-Kuleta, J.; Kozik, A. Moonlighting Proteins as Virulence Factors of Pathogenic Fungi, Parasitic Protozoa and Multicellular Parasites. Mol. Oral Microbiol. 2014, 29, 270–283. [Google Scholar] [CrossRef]
- Santos, L.P.A.; Assunção, L.d.P.; Lima, P.d.S.; Tristão, G.B.; Brock, M.; Borges, C.L.; Silva-Bailão, M.G.; Soares, C.M.d.A.; Bailão, A.M. Propionate Metabolism in a Human Pathogenic Fungus: Proteomic and Biochemical Analyses. IMA Fungus 2020, 11, 9. [Google Scholar] [CrossRef]
- Portis, I.G.; de Sousa Lima, P.; Paes, R.A.; Oliveira, L.N.; Pereira, C.A.; Parente-Rocha, J.A.; Pereira, M.; Nosanchuk, J.D.; de Almeida Soares, C.M. Copper Overload in Paracoccidioides lutzii Results in the Accumulation of Ergosterol and Melanin. Microbiol. Res. 2020, 239, 126524. [Google Scholar] [CrossRef]
- Yadav, M.K.; Park, S.-W.; Chae, S.-W.; Song, J.-J. Sinefungin, a Natural Nucleoside Analogue of S-Adenosylmethionine, Inhibits Streptococcus pneumoniae Biofilm Growth. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ries, L.N.A.; Beattie, S.; Cramer, R.A.; Goldman, G.H. Overview of Carbon and Nitrogen Catabolite Metabolism in the Virulence of Human Pathogenic Fungi: Overview of Carbon and Nitrogen Catabolite Metabolism. Mol. Microbiol. 2018, 107, 277–297. [Google Scholar] [CrossRef]
- Fraser, J.A.; Davis, M.A.; Hynes, M.J. The Formamidase Gene of Aspergillus nidulans: Regulation by Nitrogen Metabolite Repression and Transcriptional Interference by an Overlapping Upstream Gene. Genetics 2001, 157, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.F.A.; Navarro, M.V.; de Barros, Y.N.; Silva, R.S.; Xander, P.; Batista, W.L. Updates in Paracoccidioides Biology and Genetic Advances in Fungus Manipulation. J. Fungi 2021, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, M.; Zhang, Z. Quantitative Proteomics Reveals the Antifungal Effect of Canthin-6-One Isolated from Ailanthus Altissima against Fusarium oxysporum f. sp. Cucumerinum in Vitro. PLoS ONE 2021, 16, e0250712. [Google Scholar] [CrossRef] [PubMed]
- Amich, J. MSphere of Influence: The Importance of Metabolism for Pathogen Adaptation to Host-Imposed Stresses. Msphere 2019, 4, e00566-19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | MIC | MFC | CC | SI |
|---|---|---|---|---|
| P01 | 6.09/1.95 | 6.09/1.95 | 4685.88/1500 | 769.43 |
| P03 | 0.75/0.24 | 0.75/0.24 | 6247.84 | |
| Pb18 | 0.75/0.24 | 0.75/0.24 | 6247.84 | |
| EPM83 | 1.5/0.48 | 1.5/0.48 | 3123.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, L.d.C.; Silva, K.S.F.e.; Rocha, O.B.; Barbosa, K.L.B.; Rozada, A.M.F.; Gauze, G.d.F.; Soares, C.M.d.A.; Pereira, M. Proteomic Response of Paracoccidioides brasiliensis Exposed to the Antifungal 4-Methoxynaphthalene-N-acylhydrazone Reveals Alteration in Metabolism. J. Fungi 2023, 9, 66. https://doi.org/10.3390/jof9010066
Silva LdC, Silva KSFe, Rocha OB, Barbosa KLB, Rozada AMF, Gauze GdF, Soares CMdA, Pereira M. Proteomic Response of Paracoccidioides brasiliensis Exposed to the Antifungal 4-Methoxynaphthalene-N-acylhydrazone Reveals Alteration in Metabolism. Journal of Fungi. 2023; 9(1):66. https://doi.org/10.3390/jof9010066
Chicago/Turabian StyleSilva, Lívia do Carmo, Kleber Santiago Freitas e Silva, Olívia Basso Rocha, Katheryne Lohany Barros Barbosa, Andrew Matheus Frederico Rozada, Gisele de Freitas Gauze, Célia Maria de Almeida Soares, and Maristela Pereira. 2023. "Proteomic Response of Paracoccidioides brasiliensis Exposed to the Antifungal 4-Methoxynaphthalene-N-acylhydrazone Reveals Alteration in Metabolism" Journal of Fungi 9, no. 1: 66. https://doi.org/10.3390/jof9010066