
Phylogenesis of the Functional 1-Aminocyclopropane-1-Carboxylate Oxidase of Fungi and Plants
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Bioinformatics Analysis
2.3. RT-PCR
2.4. Construction of Mushroom ACO Expression Vectors
2.5. Mushroom ACO Expression and Purification
2.6. ACO Enzyme Activity Assay
3. Results
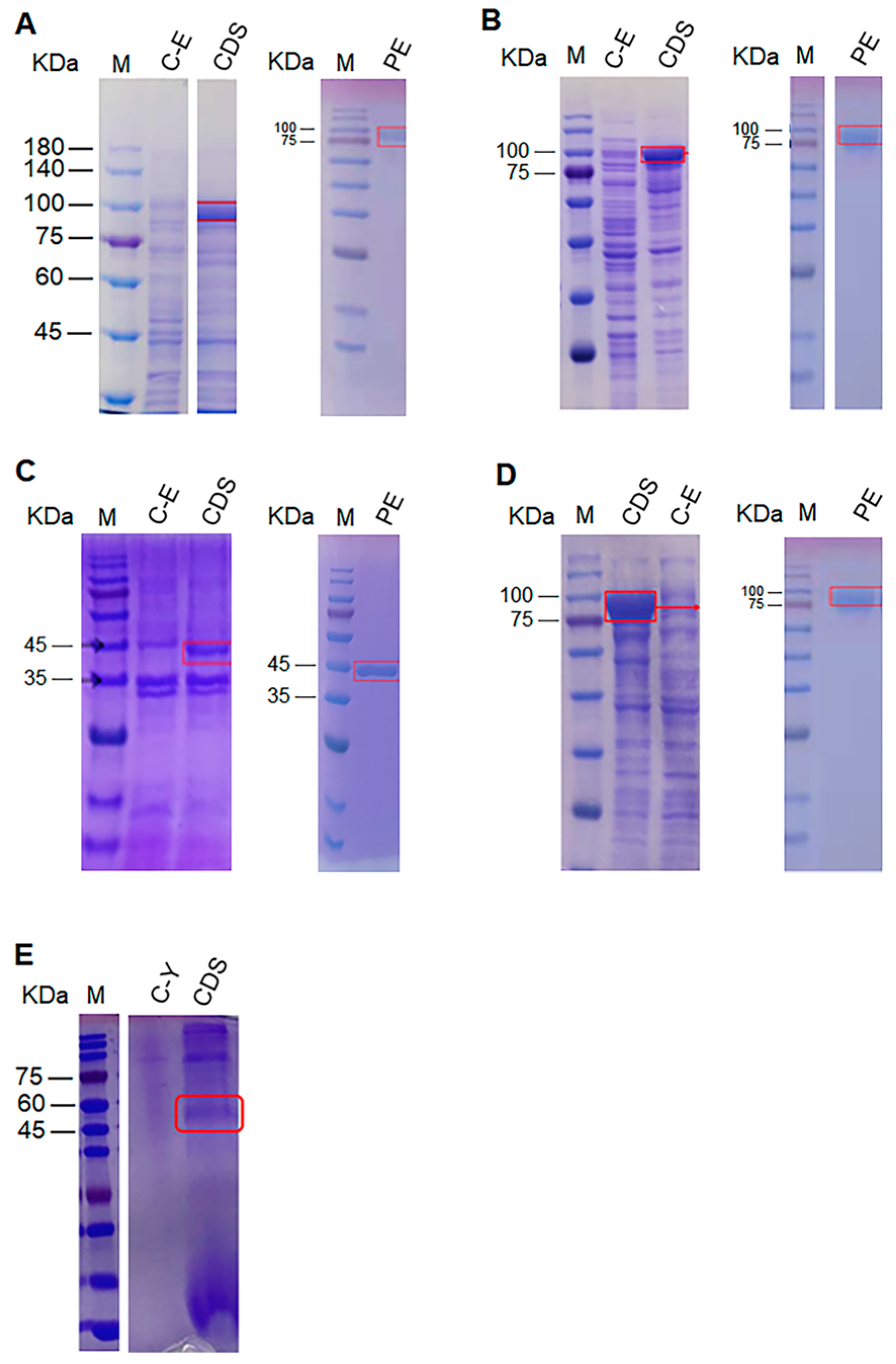
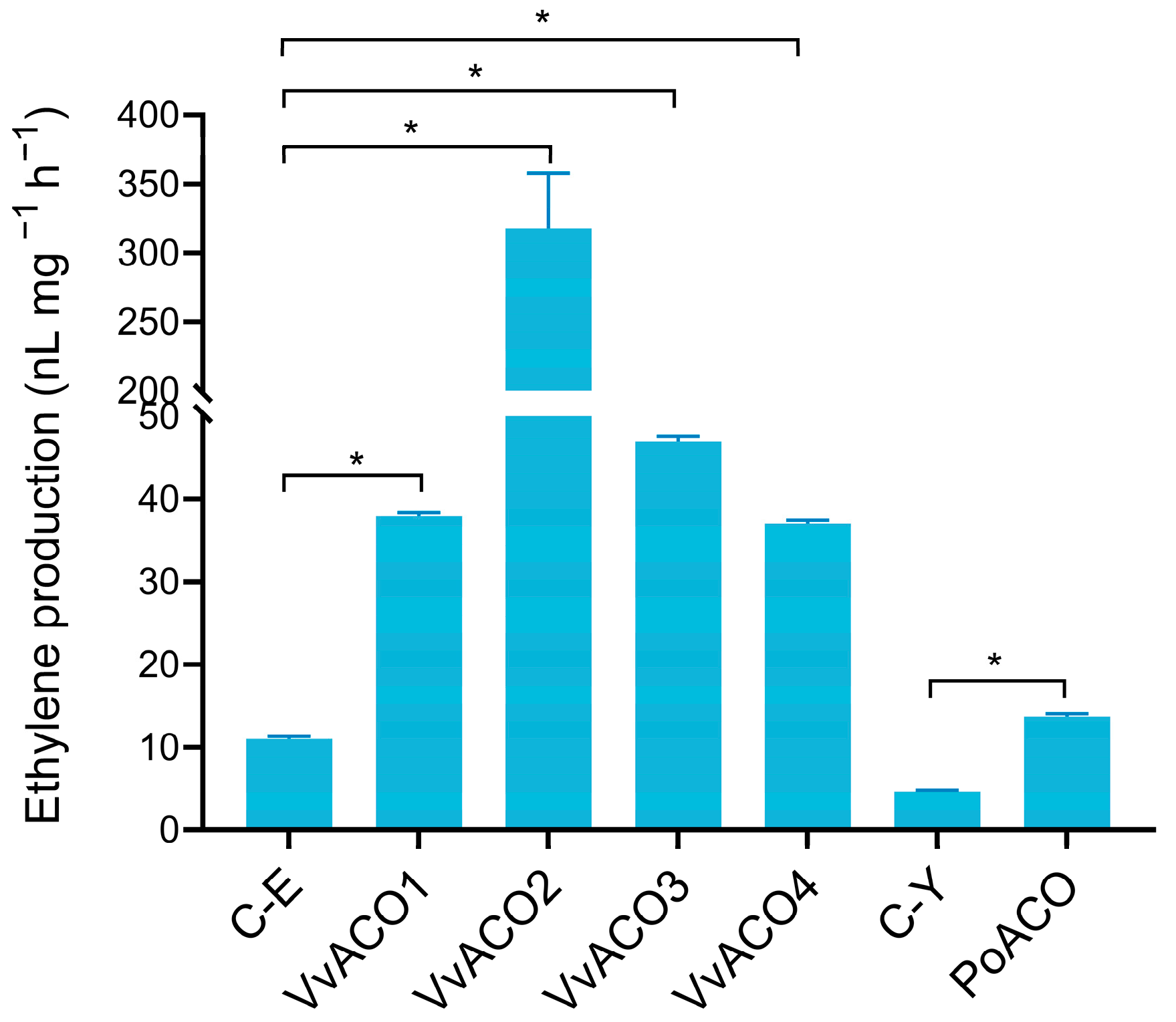
3.1. Cloning, Expression and Enzyme Activities of ACO Proteins from Straw Mushrooms and Oyster Mushrooms
3.2. Residue Analysis of Functional Fungi, Slime Mold and Plant ACOs
3.3. Phylogenetic Analysis of Microbial and Plant ACOs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poel, B.V.D.; Cooper, E.D.; Delwiche, C.F.; Chang, C. An evolutionary perspective on the plant hormone ethylene. In Ethylene in Plants; Wen, C.K., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 109–134. [Google Scholar]
- Kende, H. Ethylene biosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 283–307. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.-C. Plant growth promotion under water: Decrease of waterlogging-induced ACC and ethylene levels by ACC deaminase-producing bacteria. Front. Microbiol. 2018, 9, 1096. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Ogawa, T.; Ishihara, K.; Fujii, T.; Nagahama, K.; Omata, T.; Inoue, Y.; Tanase, S.; Morino, Y. Molecular cloning in Escherichia coli, expression, and nucleotide sequence of the gene for the ethylene-forming enzyme of Pseudomonas syringae pv. phaseolicola PK2. Biochem. Biophys. Res. Commun. 1992, 188, 826–832. [Google Scholar] [CrossRef]
- Tao, L.; Dong, H.J.; Chen, X.; Chen, S.F.; Wang, T.H. Expression of ethylene-forming enzyme (EFE) of Pseudomonas syringae pv. glycinea in Trichoderma viride. Appl. Microbiol. Biotechnol. 2008, 80, 573. [Google Scholar] [CrossRef]
- Martinez, S.; Fellner, M.; Herr, C.Q.; Ritchie, A.; Hu, J.; Hausinger, R.P. Structures and mechanisms of the non-heme Fe(II)-and 2-oxoglutarate-dependent ethylene-forming enzyme: Substrate binding creates a twist. J. Am. Chem. Soc. 2017, 139, 11980–11988. [Google Scholar] [CrossRef]
- Zhang, Z.; Smart, T.J.; Choi, H.; Hardy, F.; Lohans, C.T.; Abboud, M.I.; Richardson, M.S.W.; Paton, R.S.; McDonough, M.A.; Schofield, C.J. Structural and stereoelectronic insights into oxygenase-catalyzed formation of ethylene from 2-oxoglutarate. Proc. Natl. Acad. Sci. USA 2017, 114, 4667–4672. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Martinez, S.; Hausinger, R.P.; Emerson, J.P. Thermodynamics of iron (II) and substrate binding to the ethylene-forming enzyme. Biochemistry 2018, 57, 5696–5705. [Google Scholar] [CrossRef]
- Weingart, H.; Völksch, B.; Ullrich, M.S. Comparison of ethylene production by Pseudomonas syringae and Ralstonia solanacearum. Phytopathology 1999, 89, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Valls, M.; Genin, S.; Boucher, C. Integrated regulation of the type III secretion system and other virulence determinants in Ralstonia solanacearum. PLoS Pathog. 2006, 2, e82. [Google Scholar] [CrossRef] [Green Version]
- Zavřel, T.; Knoop, H.; Steuer, R.; Jones, P.R.; Červený, J.; Trtílek, M. A quantitative evaluation of ethylene production in the recombinant cyanobacterium Synechocystis sp. PCC 6803 harboring the ethylene-forming enzyme by membrane inlet mass spectrometry. Bioresour Technol. 2016, 202, 142–151. [Google Scholar] [CrossRef]
- Huang, T.C.; Chow, T.J. Ethylene production by blue-green algae. Bot. Bull. Acad. Sin. 1984, 25, 81–86. [Google Scholar]
- Hottiger, T.; Boller, T. Ethylene biosynthesis in Fusarium oxysporum f. sp. tulipae proceeds from glutamate/2-oxoglutarate and requires oxygen and ferrous ions in vivo. Arch. Microbiol. 1991, 157, 18–22. [Google Scholar] [CrossRef]
- Pazout, J.; Pazoutova, S. Ethylene is synthesised by vegetative mycelium in surface cultures of Penicillium cyclopium Westling. Can. J. Microbiol. 1989, 35, 384–387. [Google Scholar] [CrossRef]
- Fukudaa, H.; Fujiia, T.; Ogawaa, T. Preparation of a cell-free ethylene- forming system from Penicillium digitatum. Agric. Biol. Chem. 1986, 50, 977–981. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Arshad, M.; Khalid, A.; Mahmood, M.H. Substrate-dependent biosynthesis of ethylene by rhizosphere soil fungi and its influence on etiolated pea seedlings. Pedobiologia 2005, 49, 211–219. [Google Scholar] [CrossRef]
- Chagué, V.; Elad, Y.; Barakat, R.; Tudzynski, P.; Sharon, A. Ethylene biosynthesis in Botrytis cinerea. FEMS Microbiol. Ecol. 2002, 40, 143–149. [Google Scholar] [CrossRef]
- Cristescu, S.M.; De Martinis, D.; Hekkert, S.L.; Parker, D.H.; Harren, F.J.M. Ethylene production by Botrytis cinerea in vitro and in tomatoes. Appl. Environ. Microbiol. 2002, 68, 5342–5350. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, D.; DeVay, J. Ethylene production and toxi genicity of methionine and its derivatives with riboflavin in cultures of Verticillium, Fusarium and Colletotrichum species exposed to light. Physiol. Plant 1984, 62, 545–552. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, T.; Shen, C.; Wang, X.; Qi, Y.; Shen, J.; Song, A.; Qiu, L.; Ai, Y. Downregulation of ethylene production increases mycelial growth and primordia formation in the button culinary-medicinal mushroom, Agaricus bisporus (Agaricomycetes). Int. J. Med. Mushrooms 2016, 18, 1131–1140. [Google Scholar] [CrossRef]
- Meng, D.; Shen, L.; Yang, R.; Zhang, X.; Sheng, J. Identification and active site analysis of the 1-aminocyclopropane-1-carboxylic acid oxidase catalysing the synthesis of ethylene in Agaricus bisporus. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 120–128. [Google Scholar] [CrossRef]
- Amagai, A.; Maeda, Y. The ethylene action in the development of cellular slime molds: An analogy to higher plants. Protoplasma 1992, 167, 159–168. [Google Scholar] [CrossRef]
- Li, D.; Mou, W.; Van de Poel, B.; Chang, C. Something old, something new: Conservation of the ethylene precursor 1-amino-cyclopropane-1-carboxylic acid as a signaling molecule. Curr Opin Plant Biol. 2022, 65, 102116. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Jimenez, P.; Robaina, R.R. Effects of ethylene on tetrasporogenesis in Pterocladiella capillacea (Rhodophyta). J. Phycol. 2012, 48, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Maillard, P.; Thepenier, C.; Gudin, C. Determination of an ethylene biosynthesis pathway in the unicellular green alga, Haematococcus pluvialis. Relationship between growth and ethylene production. J. Appl. Phycol. 1993, 5, 93–98. [Google Scholar] [CrossRef]
- Driessche, T.V.; Kevers, C.; Collet, M.; Gaspar, T. Acetabularia mediterranea and ethylene: Production in relation with development, circadian rhythms in emission, and response to external application. J. Plant Physiol. 1988, 133, 635–639. [Google Scholar] [CrossRef]
- Plettner, I.N.A.; Steinke, M.; Malin, G. Ethene (ethylene) production in the marine macroalga Ulva (Enteromorpha) intestinalis L. (Chlorophyta, Ulvophyceae): Effect of light-stress and co-production with dimethyl sulphide. Plant Cell Environ. 2005, 28, 1136–1145. [Google Scholar] [CrossRef]
- Ju, C.; Van de Poel, B.; Cooper, E.D.; Thierer, J.H.; Gibbons, T.R.; Delwiche, C.F.; Chang, C. Conservation of ethylene as a plant hormone over 450 million years of evolution. Nat. Plants 2015, 1, 14004. [Google Scholar] [CrossRef]
- Rohwer, F.; Bopp, M. Ethylene synthesis in moss protonema. J. Plant Physiol. 1985, 117, 331–338. [Google Scholar] [CrossRef]
- Chernys, J.; Kende, H. Ethylene biosynthesis in Regnellidium diphyllum and Marsilea quadrifolia. Planta 1996, 200, 113–118. [Google Scholar] [CrossRef]
- Osborne, D.J.; Walters, J.; Milborrow, B.V.; Norville, A.; Stange, L.M.C. Evidence for a non-ACC ethylene biosynthesis pathway in lower plants. Phytochemistry 1996, 42, 51–60. [Google Scholar] [CrossRef]
- Uji, T.; Matsuda, R.; Takechi, K.; Takano, H.; Mizuta, H.; Takio, S. Ethylene regulation of sexual reproduction in the marine red alga Pyropia yezoensis (Rhodophyta). J. Appl. Phycol. 2016, 28, 3501–3509. [Google Scholar] [CrossRef]
- Endo, H.; Mizuta, H.; Uji, T. α-aminoisobutyric acid mimics the effect of 1-aminocyclopropane-1-carboxylic acid to promote sexual reproduction in the marine red alga Pyropia yezoensis (Rhodophyta). J. Appl. Phycol. 2021, 33, 1081–1087. [Google Scholar] [CrossRef]
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayose, A.; Kanda, A.; Kubo, Y.; Takahashi, T.; Motose, H. Distinct functions of ethylene and ACC in the basal land plant Marchantia polymorpha. Plant Cell Physiol. 2021, 62, 858–871. [Google Scholar] [CrossRef] [PubMed]
- Van de Poel, B.; Chang, C. Is losing ethylene a losing game? Mol. Plant 2022, 15, 788–790. [Google Scholar] [CrossRef]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Dilley, D.R.; Wang, Z.; Kadirjan-Kalbach, D.K.; Ververidis, F.; Beaudry, R.; Padmanabhan, K. 1-aminocyclopropane-1-carboxylic acid oxidase reaction mechanism and putative post-translational activities of the ACCO protein. AoB Plants 2013, 5, plt031. [Google Scholar] [CrossRef] [Green Version]
- Aravind, L.; Koonin, E.V. The DNA-repair protein AlkB, EGL-9, and leprecan define new families of 2-oxoglutarate- and iron-dependent dioxygenases. Genome Biol. 2001, 2, research0007.1. [Google Scholar] [CrossRef]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-carboxylic acid oxidase (ACO): The enzyme that makes the plant hormone ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Qi, Y.; Gao, Y.; Shen, J.; Qiu, L. Preliminary exploring on the mechanism of casing soil stimulating the primordium formation of Agaricus bisporus. Edible Fungi 2010, 32, 9–11. [Google Scholar]
- Eun, H.D.; Ali, S.; Jung, H.; Kim, K.; Kim, W.C. Profiling of ACC synthase gene (ACS11) expression in Arabidopsis induced by abiotic stresses. Appl. Biol. Chem. 2019, 62, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Qiu, C.; Huang, T.; Zhou, W.; Qi, Y.; Gao, Y.; Shen, J.; Qiu, L. Effect of 1-aminocyclopropane-1-carboxylic acid deaminase producing bacteria on the hyphal growth and primordium initiation of Agaricus bisporus. Fungal Ecol. 2013, 6, 110–118. [Google Scholar] [CrossRef]
- Li, T.; Zhang, J.; Gao, X.; Chen, J.; Zheng, Y.; Gao, Y.; Qiu, L. The molecular mechanism for the ethylene regulation of postharvest button mushrooms maturation and senescence. Postharvest Biol. Technol. 2019, 156, 110930. [Google Scholar] [CrossRef]
- Shui, P.R.; Zheng, X.B.; Lin, J.F.; Guo, L.Q. A rapid and efficient method for isolating high quality total RNA from edible fungi. Acta Edulis Fungi 2008, 15, 32–36. [Google Scholar]
- Furqan, B.R.N. Heterologous expression and characterization of thermostable lipase (Lk1) in Pichia pastoris GS115. Biocatal Agric Biotechnol. 2020, 23, 101448. [Google Scholar] [CrossRef]
- Zhang, C.; Shang, D.; Zhang, Y.; Gao, X.; Liu, D.; Gao, Y.; Li, Y.; Qi, Y.; Qiu, L. Two hybrid histidine kinases involved in the ethylene regulation of the mycelial growth and postharvest fruiting body maturation and senescence of Agaricus bisporus. Microbiol. Spectrum 2022, 10, 02411–02422. [Google Scholar] [CrossRef]
- Bao, D.; Gong, M.; Zheng, H.; Chen, M.; Zhang, L.; Wang, H.; Jiang, J.; Wu, L.; Zhu, Y.; Zhu, G.; et al. Sequencing and comparative analysis of the straw mushroom (Volvariella volvacea) genome. PLoS ONE 2013, 8, e58294. [Google Scholar] [CrossRef]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Vandenbussche, F.; Vriezen, W.H.; Smalle, J.; Laarhoven, L.J.J.; Harren, F.J.M.; Van Der Straeten, D. Ethylene and auxin control the Arabidopsis response to decreased light intensity. Plant Physiol. 2003, 133, 517–527. [Google Scholar] [CrossRef] [Green Version]
- Raz, V.; Ecker, J.R. Regulation of differential growth in the apical hook of Arabidopsis. Development 1999, 126, 3661–3668. [Google Scholar] [CrossRef]
- Dong, J.G.; Olson, D.; Silverstone, A.; Yang, S.F. Sequence of a cDNA coding for a 1-aminocyclopropane-1-carboxylate oxidase homolog from apple fruit. Plant Physiol. 1992, 98, 1530–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouse, R.M.; Carraro, N. A novel phylogeny and morphological reconstruction of the PIN genes and first phylogeny of the ACC-oxidases (ACOs). Front. Plant Sci. 2014, 5, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellidou, I.; Buts, K.; Hatoum, D.; Ho, Q.T.; Johnston, J.W.; Watkins, C.B.; Schaffer, R.J.; Gapper, N.E.; Giovannoni, J.J.; Rudell, D.R.; et al. Transcriptomic events associated with internal browning of apple during postharvest storage. BMC Plant Biol. 2014, 14, 328. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Bouzayen, M.; Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. USA 1991, 88, 7434–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sell, S.; Hehl, R. A fifth member of the tomato 1-aminocyclopropane-1-carboxylic acid (ACC) oxidase gene family harbours a leucine zipper and is anaerobically induced. J. DNA Seq. Mapp. 2005, 16, 80–82. [Google Scholar] [CrossRef]
- Zhang, Z.; Mao, C.; Shi, Z.; Kou, X. The amino acid metabolic and carbohydrate metabolic pathway play important roles during salt-stress response in tomato. Front. Plant Sci. 2017, 8, 1231. [Google Scholar] [CrossRef] [Green Version]
- Gallie, D.R.; Young, T.E. The ethylene biosynthetic and perception machinery is differentially expressed during endosperm and embryo development in maize. Mol. Genet. Genomics 2004, 271, 267–281. [Google Scholar] [CrossRef]
- Chae, H.S.; Cho, Y.G.; Park, M.Y.; Lee, M.C.; Eun, M.Y.; Kang, B.G.; Kim, W.T. Hormonal cross-talk between auxin and ethylene differentially regulates the expression of two members of the 1-aminocyclopropane-1-carboxylate oxidase gene family in rice (Oryza sativa L.). Plant Cell Physiol. 2000, 41, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Iwai, T.; Miyasaka, A.; Seo, S.; Ohashi, Y. Contribution of ethylene biosynthesis for resistance to blast fungus infection in young rice plants. Plant Physiol. 2006, 142, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Hudgins, J.W.; Ralph, S.G.; Franceschi, V.R.; Bohlmann, J. Ethylene in induced conifer defense: cDNA cloning, protein expression, and cellular and subcellular localization of 1-aminocyclopropane-1-carboxylate oxidase in resin duct and phenolic parenchyma cells. Planta 2006, 224, 865–877. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, Y.; Dean, J.F. ACC oxidase genes expressed in the wood-forming tissues of loblolly pine (Pinus taeda L.) include a pair of nearly identical paralogs (NIPs). Gene 2010, 453, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Dean, J.F. Differential responses of the promoters from nearly identical paralogs of loblolly pine (Pinus taeda L.) ACC oxidase to biotic and abiotic stresses in transgenic Arabidopsis thaliana. Planta 2010, 232, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Chou, Y.; Chang, R.; Yang, S.F. Characterization of the ferrous ion binding sites of apple 1-aminocyclopropane-1-carboxylate oxidase by site-directed mutagenesis. Biochem. Biophys. Res. Commun. 1996, 700, 697–700. [Google Scholar] [CrossRef]
- Zhang, Z.; Ren, J.-S.; Clifton, I.J.; Schofield, C.J. Crystal structure and mechanistic implications of 1-aminocyclopropane-1-carboxylic acid oxidase—The ethylene-forming enzyme. Chem. Biol. 2004, 11, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Richards, T.A.; Soanes, D.M.; Foster, P.G.; Leonard, G.; Thornton, C.R.; Talbot, N.J. Phylogenomic analysis demonstrates a pattern of rare and ancient horizontal gene transfer between plants and fungi. Plant Cell 2009, 21, 1897–1911. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, D.C. Evolution of fungal wood decay. In Deterioration and Protection of Sustainable Biomaterials, ACS Symposium Series; Schultz, T.P., Goodell, B., Nicholas, D.D., Eds.; Amercian Chemical Society: Washington, DC, USA, 2014; pp. 93–112. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Nucleotide Sequence (5′–3′) | Application |
|---|---|---|
| PoACO-F | ATGCCTACGAAGGCATCTACA | Cloning of PoACO |
| PoACO-R | TTAGTTGAAATGCTTAATAACAGTTCCACGAATCC | |
| pPIC9K-ACO-F | AAGGCGAATTAATTCGCGGCCGCATGCCGACCAAAGCAAGCA | Construction for pPIC9K-PoACO |
| pPIC9K-ACO-R | GCTGAAGCTTACGTAGAATTCTTAATGATGATGATGATGATGAA- TCACTGTACCACG | |
| VvACO3-F | GGATCCATGTCCCATCTCAATTCTGCAGCC | Cloning of VvACOs and Construction for the VvACOs expression vectors |
| VvACO3-R | AAGCTTTTAATAGGCACCGACTTGCCCG | |
| VvACO1-F | GGAATTCCATATGATGGCGGCTCCTAAATCTACC | |
| VvACO1-R | GCTCTAGATTAATACCTCCCCTTCTTCTCGTC | |
| VvACO2-F | CGGGATCCATGGCTGGCTTTGCCACGT | |
| VvACO2-R | GCTCTAGATCAAGCCTCAATGCGCTCAG | |
| VvACO4-F | CGGGATCCATGTTTATCTTAAGAACTCGGCTTC | |
| VvACO4-R | GCTCTAGATTACGAGTAGGTCACTGCCAGG |
| Species | Protein Name | Protein ID | Type | Protein (aa) | Reference |
|---|---|---|---|---|---|
| Volvariella volvacea | VvACO4 | JGI 111930 | 4 | 354 | This study |
| VvACO3 | JGI 116615 | 4 | 374 | This study | |
| VvACO2 | JGI 118606 | 4 | 381 | This study | |
| VvACO1 | JGI 111142 | 4 | 353 | This study | |
| Pleurotus ostreatus | PoACO | KDQ32580 | 4 | 324 | This study |
| Agaricus bisporus | AbACO | JGI 195789 | 4 | 368 | [20] |
| Dictyostelium mucoroides | DmACO | BAF64840 | 4 | 368 | [22] |
| Arabidopsis thaliana | AtACO1 | AT2G19590.1 | 2 | 310 | [50] |
| AtACO2 | AT1G62380.1 | 1 | 320 | [51] | |
| AtACO5 | AT1G77330.1 | 3 | 307 | [50] | |
| Apple (Malus domestica) | MdACO1 | MDP0000195885 | 1 | 314 | [52] |
| MdACO6 | MDP0000025650 | 3 | 298 | [53] | |
| MdACO7 | MDP0000200896 | 2 | 305 | [54] | |
| Tomato (Solanum lycopersicum) | SlACO1 | Solyc07g049530.2.1 | 1 | 315 | [55] |
| SlACO5 | Solyc07g026650.2.1 | 2 | 301 | [56] | |
| SlACO7 | Solyc06g060070.2.1 | 3 | 314 | [57] | |
| Maize (Zea mays) | ZmACO20 | Zm00008a017510_T01 | 1 | 323 | [58] |
| ZmACO15 | Zm00008a037502_T01 | 3 | 314 | [22] | |
| Rice (Oryza sativa) | OsACO1 | LOC_Os09g27820.1 | 1 | 322 | [59] |
| OsACO6 | LOC_Os06g37590.1 | 2 | 293 | [60] | |
| OsACO7 | LOC_Os01g39860.1 | 2 | 312 | [22] | |
| OsACO4 | LOC_Os11g08380.1 | 3 | 309 | [22] | |
| Douglas fir (Pseudotsuga menziesii) | PmACO | ABF20554 | 4 | 320 | [61] |
| Loblolly pine (Pinus taeda) | PtACO1 | ADD65762 | 4 | 333 | [62,63] |
| PtACO2 | ADD65761 | 4 | 333 | [62,63] | |
| PtACO3 | ADD65760 | 4 | 323 | [62,63] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Qi, M.; Zhang, Q.; Xu, Z.; Zhang, Y.; Gao, Y.; Qi, Y.; Qiu, L.; Wang, M. Phylogenesis of the Functional 1-Aminocyclopropane-1-Carboxylate Oxidase of Fungi and Plants. J. Fungi 2023, 9, 55. https://doi.org/10.3390/jof9010055
Li Y, Qi M, Zhang Q, Xu Z, Zhang Y, Gao Y, Qi Y, Qiu L, Wang M. Phylogenesis of the Functional 1-Aminocyclopropane-1-Carboxylate Oxidase of Fungi and Plants. Journal of Fungi. 2023; 9(1):55. https://doi.org/10.3390/jof9010055
Chicago/Turabian StyleLi, Yanan, Man Qi, Qi Zhang, Zhixu Xu, Yan Zhang, Yuqian Gao, Yuancheng Qi, Liyou Qiu, and Mingdao Wang. 2023. "Phylogenesis of the Functional 1-Aminocyclopropane-1-Carboxylate Oxidase of Fungi and Plants" Journal of Fungi 9, no. 1: 55. https://doi.org/10.3390/jof9010055