
MoSnf5 Regulates Fungal Virulence, Growth, and Conidiation in Magnaporthe oryzae
 ,
,  , and
, and 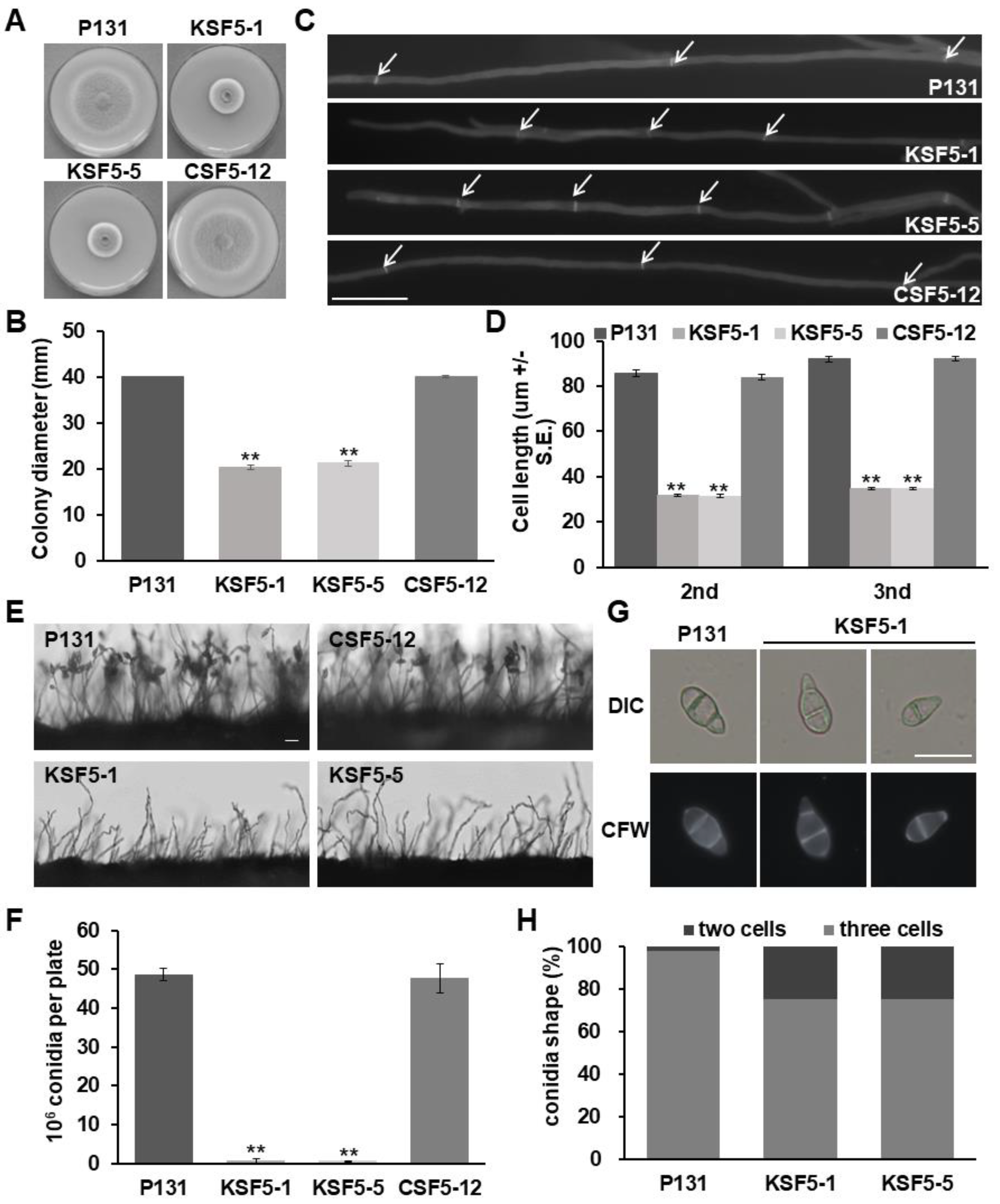{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Growth Conditions, and Transformation
2.2. Nucleic Acid Manipulation
2.3. RT-Quantitative PCR
2.4. Gene Deletion and Complementation of MoSNF5 and MoTAF14
2.5. Subcellular Localization of MoSnf5
2.6. Pathogenicity Test
2.7. Microscopy Assays
2.8. Yeast Two-Hybrid Assay
2.9. Western Blot
2.10. Pull-Down and Co-Immunoprecipitation Assay
2.11. Statistical Analysis
3. Results
3.1. MoSNF5 Is Important for Vegetative Hyphal Growth, Conidiation, and Cell Wall Integrity
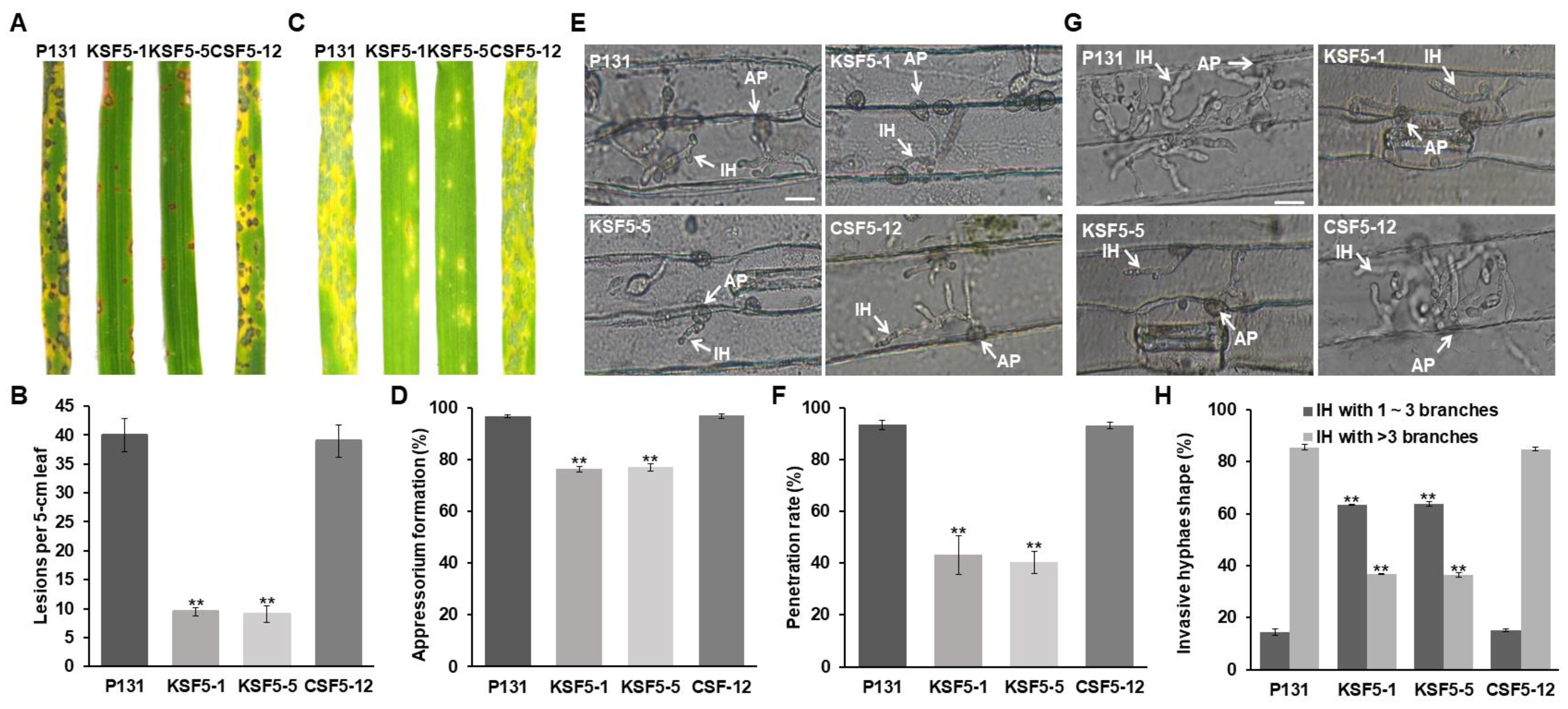
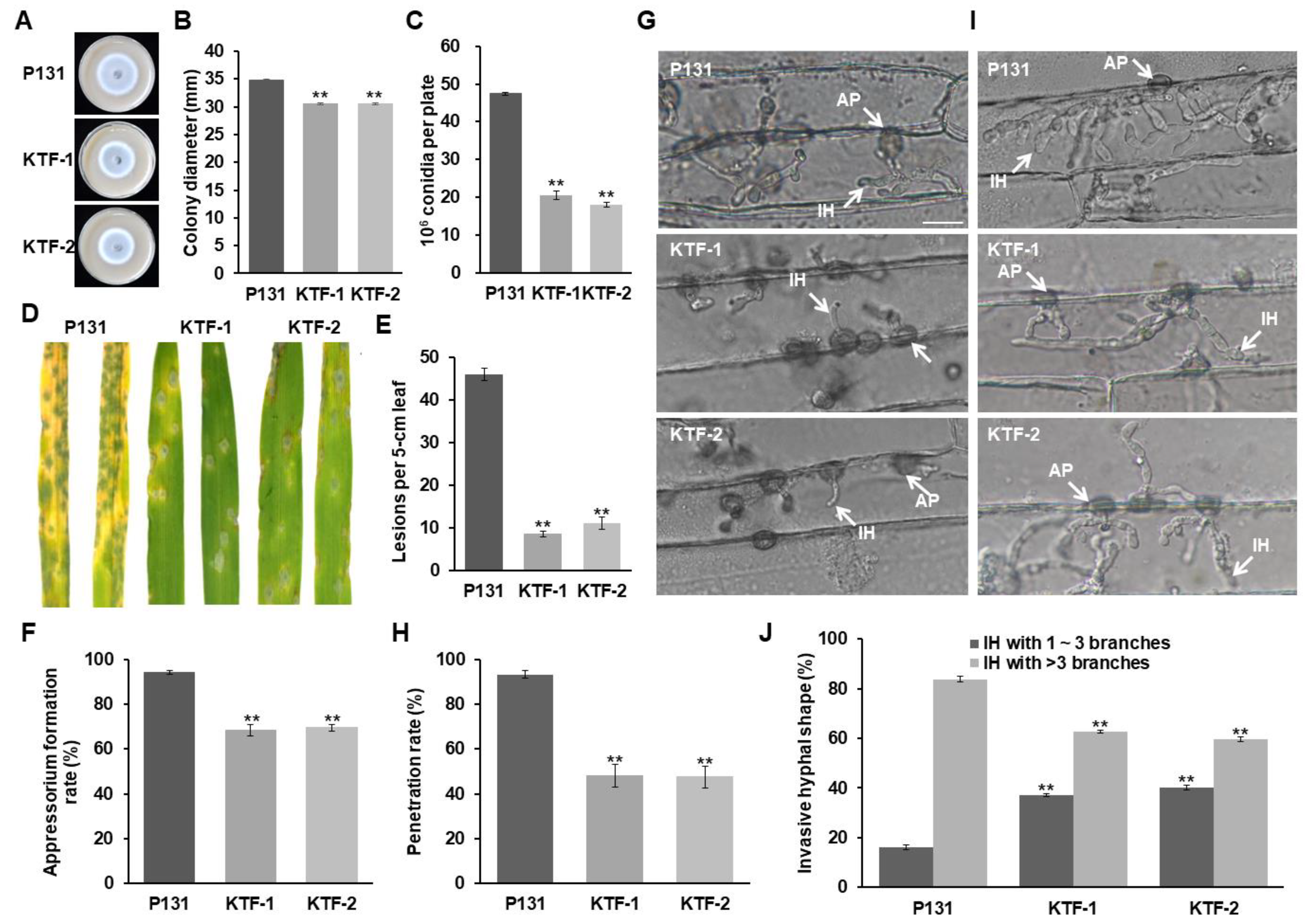
3.2. MoSNF5 Is Required for Fungal Virulence
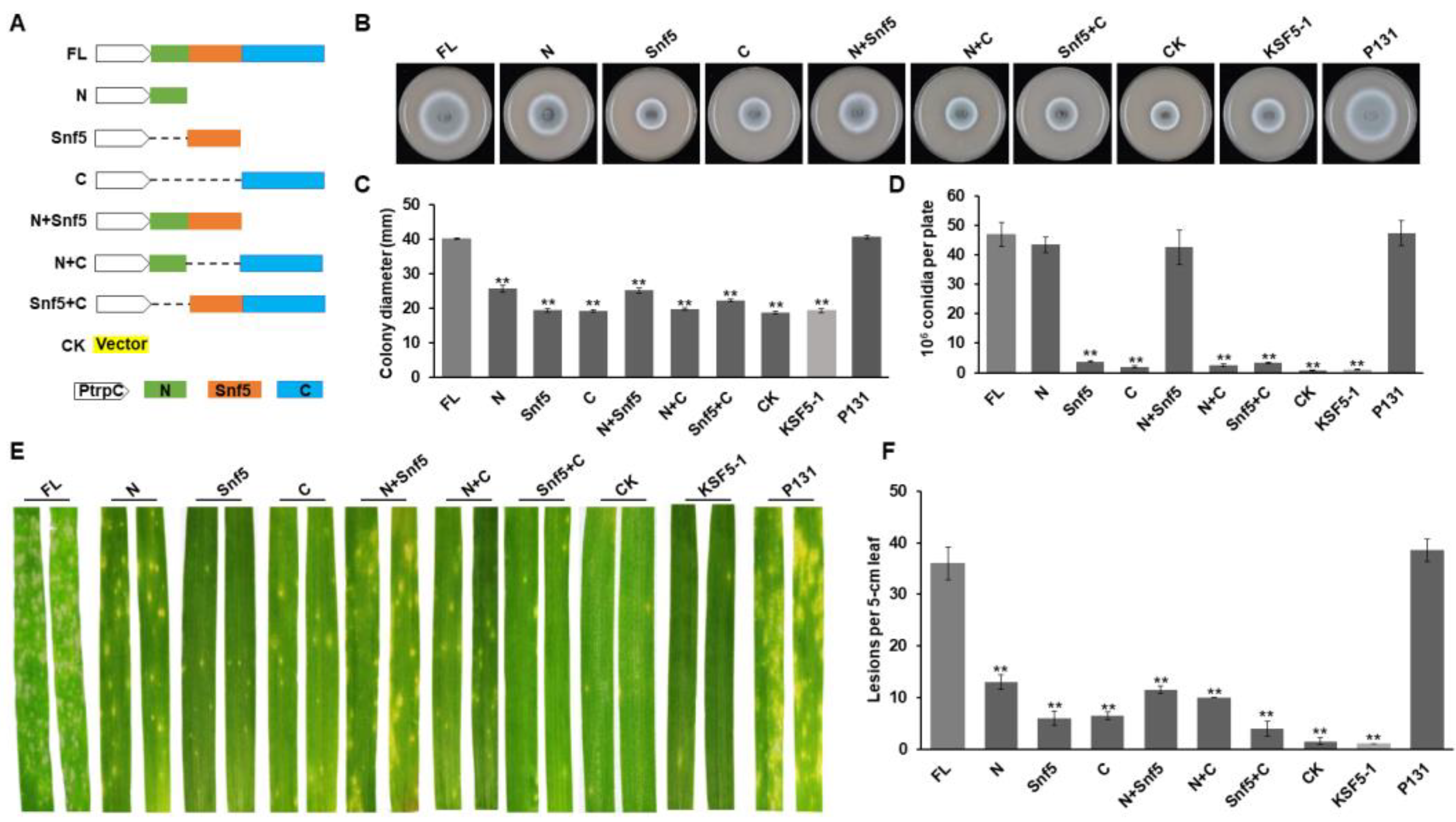
3.3. Different Regions of MoSnf5 Play Important Roles in Growth, Conidiation, and Virulence
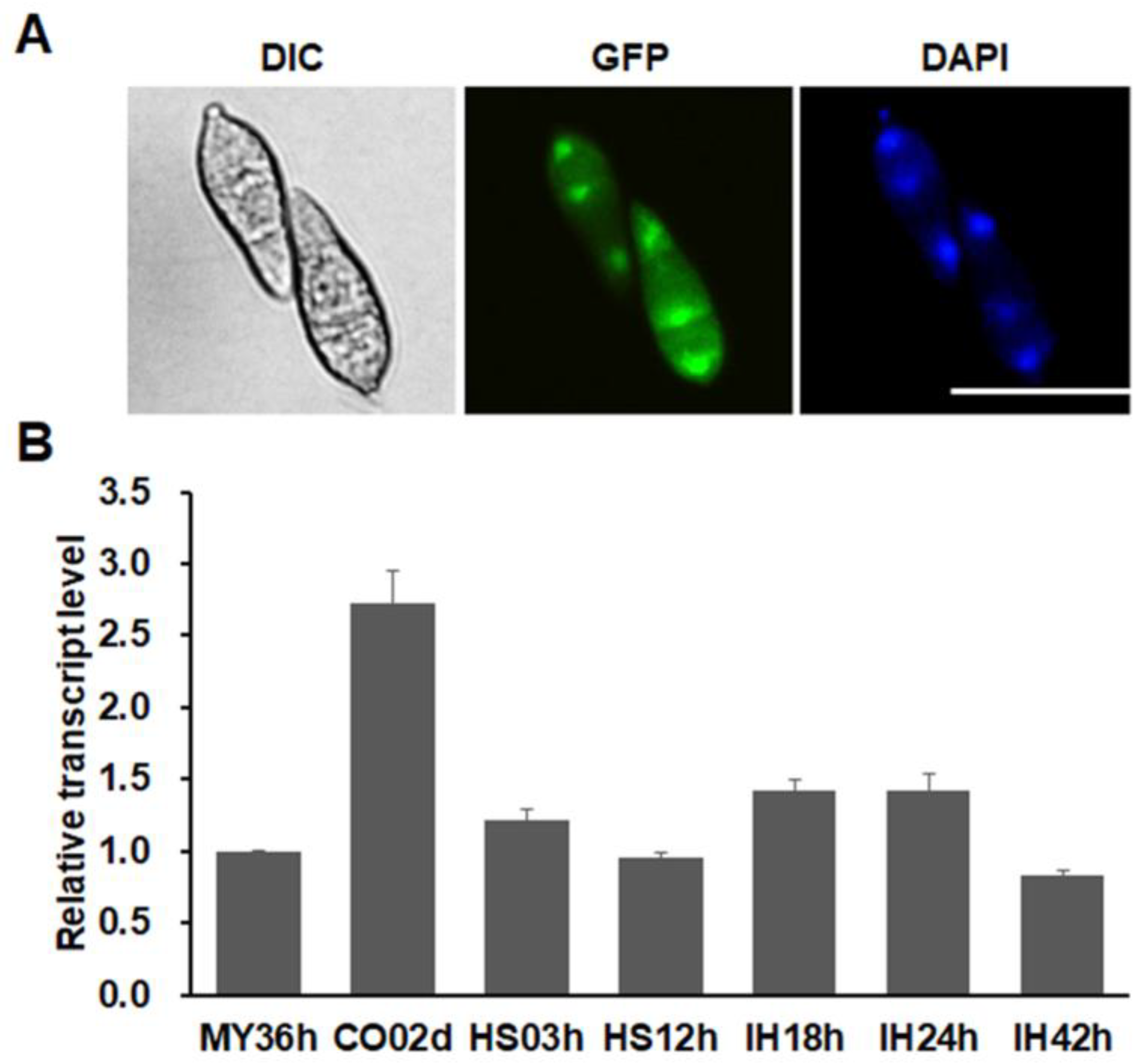
3.4. MoSnf5 Is a Nuclear Protein and Constitutively Expressed
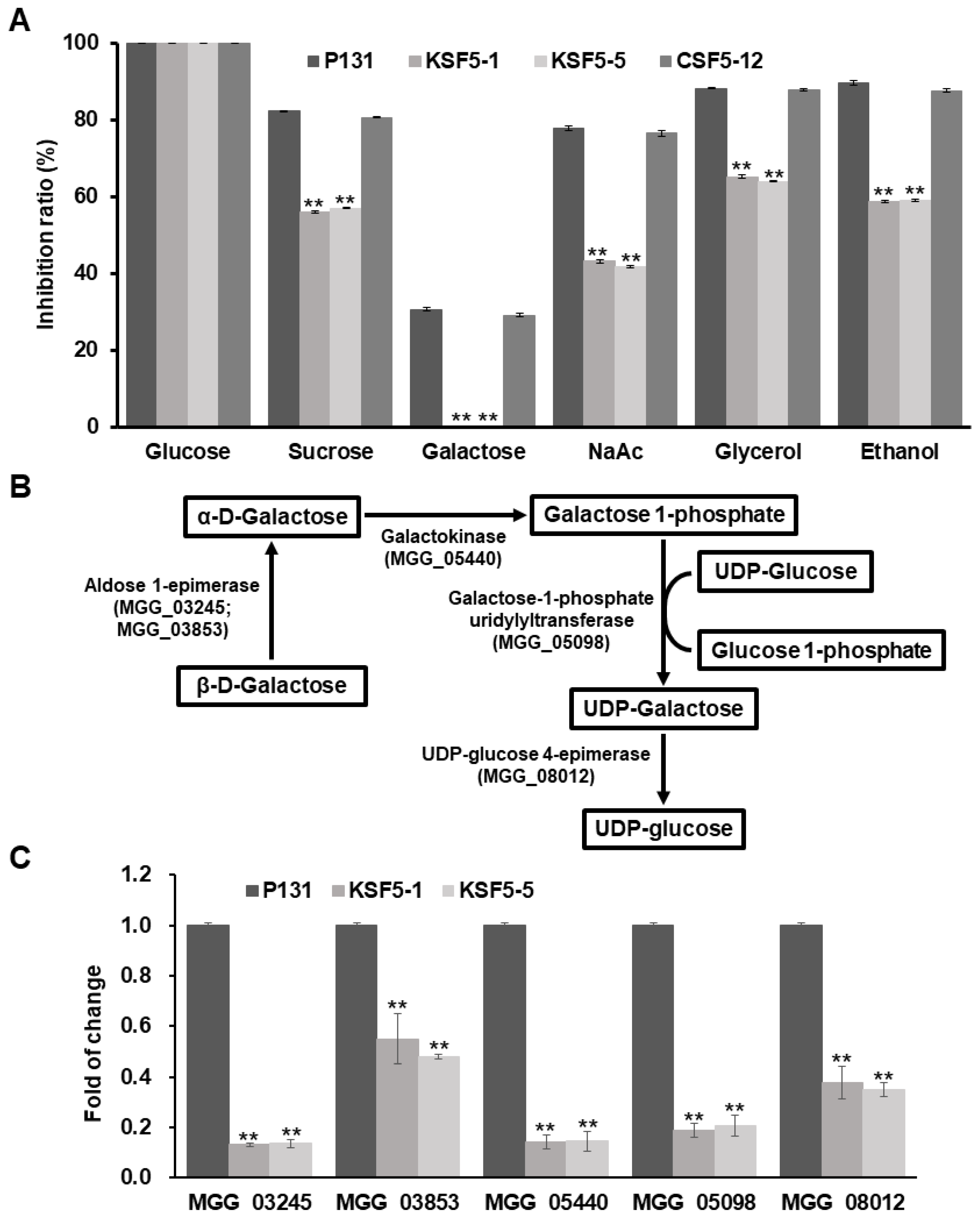
3.5. MoSNF5 Is Essential to Galactose Utilization and Regulates Expression Levels of Genes of the Leloir Pathway
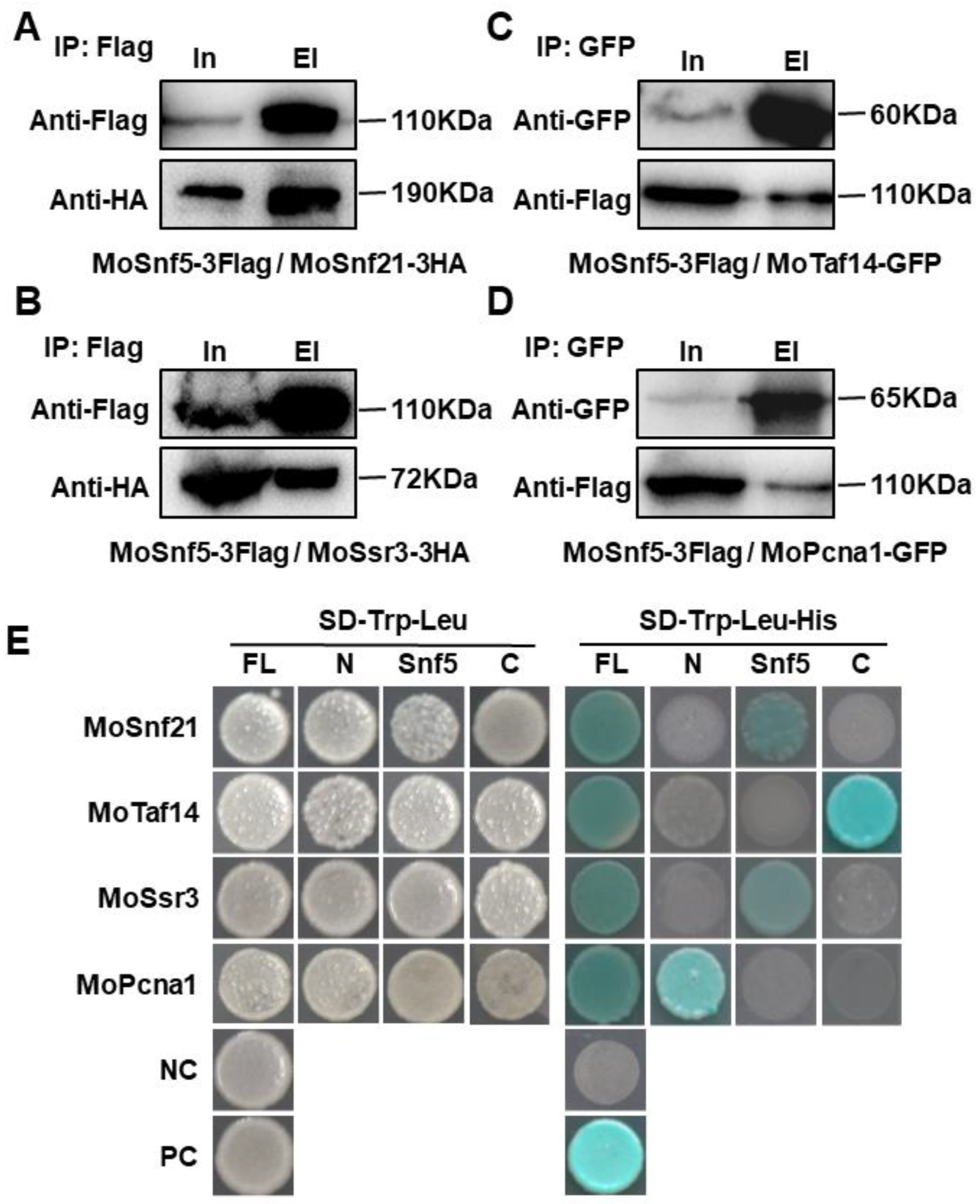
3.6. MoSnf5 Interacts with Four Proteins via Different Regions
3.7. MoSnf5-Interacting Protein MoTaf14 Is Also Required for Vegetative Hyphal Growth, Conidiation, and Virulence
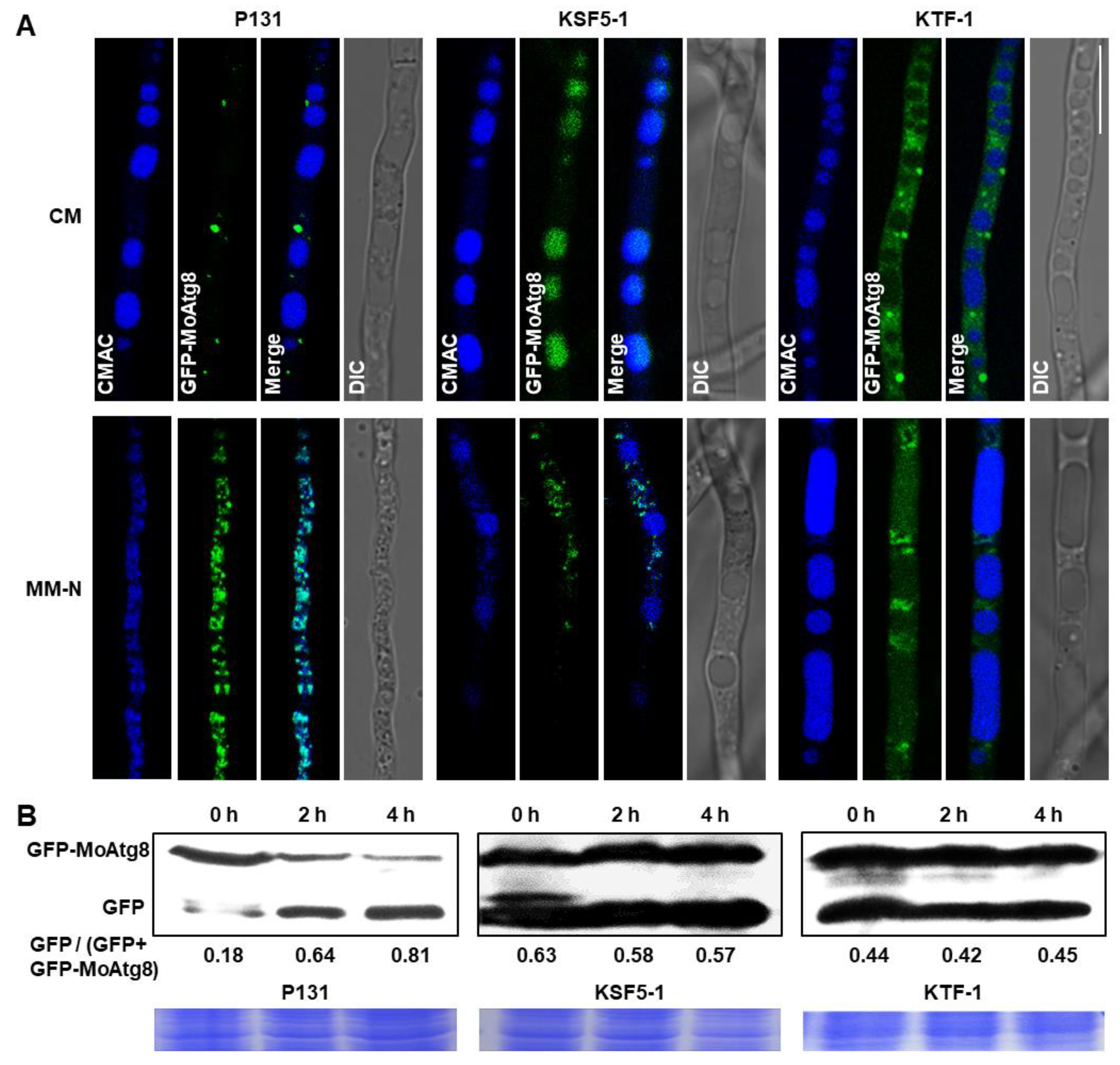
3.8. Both MoSnf5 and MoTaf14 Are Indispensable for Autophagy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ribeiro-Silva, C.; Vermeulen, W.; Lans, H. SWI/SNF: Complex complexes in genome stability and cancer. DNA Repair 2019, 77, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.L.; Dingwall, A.; Scott, M.P. Five SWI/SNF gene products are components of a large multisubunit complex required for transcriptional enhancement. Proc. Natl. Acad. Sci. USA 1994, 91, 2905–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelan, M.L.; Sif, S.; Narlikar, G.J.; Kingston, R.E. Reconstitution of a Core Chromatin Remodeling Complex from SWI/SNF Subunits. Mol. Cell 1999, 3, 247–253. [Google Scholar] [CrossRef]
- Han, Y.; Reyes, A.A.; Malik, S.; He, Y. Cryo-EM structure of SWI/SNF complex bound to a nucleosome. Nature 2020, 579, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, G.; Dong, C.; Zhao, X.; Shen, Z.; Chen, F.; Liu, B.; Long, J. Snf5 and Swi3 subcomplex formation is required for SWI/SNF complex function in yeast. Biochem. Biophys. Res. Commun. 2020, 526, 934–940. [Google Scholar] [CrossRef]
- Dastidar, R.G.; Hooda, J.; Shah, A.; Cao, T.M.; Henke, R.M.; Zhang, L. The nuclear localization of SWI/SNF proteins is subjected to oxygen regulation. Cell Biosci. 2012, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Geng, F.; Cao, Y.; Laurent, B.C. Essential roles of Snf5p in Snf-Swi chromatin remodeling in vivo. Mol. Cell. Biol. 2001, 21, 4311–4320. [Google Scholar] [CrossRef] [Green Version]
- Muchardt, C.; Sardet, C.; Bourachot, B.; Onufryk, C.; Yaniv, M. A human protein with homology to Saccharomyces cerevisiae SNF5 interacts with the potential helicase hbrm. Nucleic Acids Res. 1995, 23, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.W.; Davies, K.P.; Yung, E.; Beltran, R.J.; Yu, J.; Kalpana, G.V. c-MYC interacts with INI1/hSNF5 and requires the SWI/SNF complex for transactivation function. Nat. Genet. 1999, 22, 102–105. [Google Scholar] [CrossRef]
- Wu, D.Y.; Kalpana, G.V.; Goff, S.P.; Schubach, W.H. Epstein-Barr virus nuclear protein 2 (EBNA2) binds to a component of the human SNF-SWI complex, hSNF5/Ini1. J. Virol. 1996, 70, 6020–6028. [Google Scholar] [CrossRef]
- Neely, K.E.; Hassan, A.H.; Brown, C.E.; Howe, L.; Workman, J.L. Transcription activator interactions with multiple SWI/SNF subunits. Mol. Cell. Biol. 2002, 22, 1615–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.Y.; Fan, Y.H.; Wang, Y.L.; Zhang, S.D.; Han, C.Y. BEX2 regulates cell cycle through the interaction with INI1/hSNF5. Yi Chuan = Hered. 2012, 34, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.I.; Brittingham, G.P.; Karadeniz, Y.; Tran, K.D.; Dutta, A.; Holehouse, A.S.; Peterson, C.L.; Holt, L.J. SWI/SNF senses carbon starvation with a pH-sensitive low complexity sequence. ELife 2022, 11, e70344. [Google Scholar] [CrossRef] [PubMed]
- Ganster, R.W.; McCartney, R.R.; Schmidt, M.C. Identification of a calcineurin-independent pathway required for sodium ion stress response in Saccharomyces cerevisiae. Genetics 1998, 150, 31–42. [Google Scholar] [CrossRef]
- Neigeborn, L.; Carlson, M. Genes affecting the regulation of SUC2 gene expression by glucose repression in Saccharomyces cerevisiae. Genetics 1984, 108, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Navarro, F.J.; Nurse, P. A systematic screen reveals new elements acting at the G2/M cell cycle control. Genome Biol. 2012, 13, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S. Integrase interactor 1 in health and disease. Curr. Protein Pept. Sci. 2015, 16, 478–490. [Google Scholar] [CrossRef]
- Wang, X.; Lee, R.S.; Alver, B.H.; Haswell, J.R.; Wang, S.; Mieczkowski, J.; Drier, Y.; Gillespie, S.M.; Archer, T.C.; Wu, J.N.; et al. SMARCB1-mediated SWI/SNF complex function is essential for enhancer regulation. Nat. Genet. 2017, 49, 289–295. [Google Scholar] [CrossRef]
- Sévenet, N.; Sheridan, E.; Amram, D.; Schneider, P.; Handgretinger, R.; Delattre, O. Constitutional mutations of the hSNF5/INI1 gene predispose to a variety of cancers. Am. J. Hum. Genet. 1999, 65, 1342–1348. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhong, X.; Wang, C.; Wang, S.; Lin, L.; Zou, R.; Wu, Y.; Sun, N.; Sun, G.; Wen, T.; et al. SNF5 is involved in suppression of hepatocellular carcinoma progression via TGF-Beta 1 signaling. Anat. Rec. -Adv. Integr. Anat. Evol. Biol. 2016, 299, 869–877. [Google Scholar] [CrossRef]
- Zraly, C.B.; Dingwall, A.K. The chromatin remodeling and mRNA splicing functions of the Brahma (SWI/SNF) complex are mediated by the SNR1/SNF5 regulatory subunit. Nucleic Acids Res. 2012, 40, 5975–5987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzeski, J.; Podstolski, W.; Olczak, K.; Jerzmanowski, A. Identification and analysis of the Arabidopsis thaliana BSH gene, a member of the SNF5 gene family. Nucleic Acids Res. 1999, 27, 2393–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Cao, H.; Zhang, C.; Chen, F.; Liu, Y. The SNF5-type protein BUSHY regulates seed germination via the gibberellin pathway and is dependent on HUB1 in Arabidopsis. Planta 2022, 255, 34. [Google Scholar] [CrossRef] [PubMed]
- Finkel, J.S.; Xu, W.; Huang, D.; Hill, E.M.; Desai, J.V.; Woolford, C.A. Portrait of Candida albicans adherence regulators. PLoS Pathog. 2012, 8, e1002525. [Google Scholar] [CrossRef] [Green Version]
- Burgain, A.; Pic, É.; Markey, L.; Tebbji, F.; Kumamoto, C.A.; Sellam, A. A novel genetic circuitry governing hypoxic metabolic flexibility, commensalism and virulence in the fungal pathogen Candida albicans. PLoS Pathog. 2019, 15, e1007823. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Iyer, P.; Herkal, A.; Abdullah, J.; Stout, A.; Free, S.J. Identification and characterization of genes required for cell-to-cell fusion in Neurospora crassa. Eukaryot. Cell 2011, 10, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Nakazawa, T.; Oka, K.; Nakahori, K.; Kamada, T. Cc.snf5, a gene encoding a putative component of the SWI/SNF chromatin remodeling complex, is essential for sexual development in the agaricomycete Coprinopsis cinerea. Fungal Genet. Biol. 2013, 50, 82–89. [Google Scholar] [CrossRef]
- Nakazawa, T.; Ando, Y.; Hata, T.; Nakahori, K. A mutation in the Cc.arp9 gene encoding a putative actin-related protein causes defects in fruiting initiation and asexual development in the agaricomycete Coprinopsis cinerea. Curr. Genet. 2016, 62, 565–574. [Google Scholar] [CrossRef]
- Xu, J.R.; Peng, Y.L.; Dickman, M.B.; Sharon, A. The dawn of fungal pathogen genomics. Annu. Rev. Phytopathol. 2006, 44, 337–366. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- de Jong, J.C.; McCormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol generates turgor in rice blast. Nature 1997, 389, 244. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, J.; Wilson, R.A. Cells in cells: Morphogenetic and metabolic strategies conditioning rice infection by the blast fungus Magnaporthe oryzae. Protoplasma 2014, 251, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Martin-Urdiroz, M.; Oses-Ruiz, M.; Ryder, L.S.; Talbot, N.J. Investigating the biology of plant infection by the rice blast fungus Magnaporthe oryzae. Fungal Genet. Biol. 2016, 90, 61–68. [Google Scholar] [CrossRef]
- Peng, Y.L.; Shishiyama, J. Temporal sequence of cytological events in rice leaves infected with Pyricularia oryzae. Can. J. Bot. -Rev. Can. De Bot. 1988, 66, 730–735. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Sun, J.; Kang, Z.; Ding, S.; Xu, J.R.; Peng, Y.L. A novel protein Com1 is required for normal conidium morphology and full virulence in Magnaporthe oryzae. Molecular Plant-Microbe Interact. 2010, 23, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Park, G.; Bruno, K.S.; Staiger, C.J.; Talbot, N.J.; Xu, J.R. Independent genetic mechanisms mediate turgor generation and penetration peg formation during plant infection in the rice blast fungus. Mol. Microbiol. 2004, 53, 1695–1707. [Google Scholar] [CrossRef]
- Gao, X.; Yin, C.; Liu, X.; Peng, J.; Chen, D.; Shi, W.; Zhao, W.; Yang, J.; Peng, Y.L. A glycine-rich protein MoGrp1 functions as a novel splicing factor to regulate fungal virulence and growth in Magnaporthe oryzae. Phytopathol. Res. 2019, 1, 1–15. [Google Scholar] [CrossRef]
- Chen, X.L.; Yang, J.; Peng, Y.L. Large-scale insertional mutagenesis in Magnaporthe oryzae by Agrobacterium tumefaciens-mediated transformation. Methods Mol. Biol. 2011, 722, 213–224. [Google Scholar] [CrossRef]
- Xu, J.R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Kong, L.A.; Yang, J.; Li, G.T.; Qi, L.L.; Zhang, Y.J.; Wang, C.F.; Zhao, W.S.; Xu, J.R.; Peng, Y.L. Different chitin synthase genes are required for various developmental and plant infection processes in the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2012, 8, e1002526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Pan, X.; Chen, D.; Yin, C.; Peng, J.; Shi, W.; Qi, L.; Wang, R.; Zhao, W.; Zhang, Z.; et al. Prp19-associated splicing factor Cwf15 regulates fungal virulence and development in the rice blast fungus. Environ. Microbiol. 2021, 23, 5901–5916. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Shi, T.; Yang, J.; Shi, W.; Gao, X.; Chen, D.; Xu, X.; Xu, J.R.; Talbot, N.J.; Peng, Y.L. N-glycosylation of effector proteins by an α-1,3-mannosyltransferase is required for the rice blast fungus to evade host innate immunity. Plant Cell 2014, 26, 1360–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohneda, M.; Arioka, M.; Nakajima, H.; Kitamoto, K. Visualization of vacuoles in Aspergillus oryzae by expression of CPY–EGFP. Fungal Genet. Biol. 2002, 2002, 29–38. [Google Scholar] [CrossRef]
- Zhou, W.; Shi, W.; Xu, X.W.; Li, Z.G.; Yin, C.F.; Peng, J.B.; Pan, S.; Chen, X.L.; Zhao, W.S.; Zhang, Y.; et al. Glutamate synthase MoGlt1-mediated glutamate homeostasis is important for autophagy, virulence and conidiation in the rice blast fungus. Mol. Plant Pathol. 2018, 19, 564–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Li, G.; Lin, C.; He, C. Conidiophore stalk-less1 encodes a putative zinc-finger protein involved in the early stage of conidiation and mycelial infection in Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2009, 22, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.X.; Leung, H. Genetic analysis of sporulation in Magnaporthe grisea by chemical and insertional mutagenesis. Mol. Plant-Microbe Interact. 1995, 8, 949–959. [Google Scholar] [CrossRef]
- Odenbach, D.; Breth, B.; Thines, E.; Weber, R.W.; Anke, H.; Foster, A.J. The transcription factor Con7p is a central regulator of infection-related morphogenesis in the rice blast fungus Magnaporthe grisea. Mol. Microbiol. 2007, 64, 293–307. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.Y.; Kim, K.S.; Rho, H.S.; Chi, M.H.; Choi, J.; Park, J.; Kong, S.; Park, J.; Goh, J.; et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae. PLoS Genet. 2009, 5, e1000757. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Fukada, J.; Moriwaki, A.; Fujikawa, T.; Ohashi, M.; Hibi, T.; Hayashi, N. Mstu1, an APSES transcription factor, is required for appressorium-mediated infection in Magnaporthe grisea. Biosci. Biotechnol. Biochem. 2009, 73, 1779–1786. [Google Scholar] [CrossRef]
- Holden, H.M.; Rayment, I.; Thoden, J.B. Structure and function of enzymes of the Leloir pathway for galactose metabolism. J. Biol. Chem. 2003, 278, 43885–43888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Hirota, K.; Mizuno, K.; Shibata, T.; Ohta, K. Essential roles of Snf21, a Swi2/Snf2 family chromatin remodeler, in fission yeast mitosis. Genes Genet. Syst. 2008, 83, 361–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peil, K.; Jürgens, H.; Luige, J.; Kristjuhan, K.; Kristjuhan, A. Taf14 is required for the stabilization of transcription pre-initiation complex in Saccharomyces cerevisiae. Epigenet. Chromatin 2020, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Monahan, B.J.; Villén, J.; Marguerat, S.; Bähler, J.; Gygi, S.P.; Winston, F. Fission yeast SWI/SNF and RSC complexes show compositional and functional differences from budding yeast. Nat. Struct. Mol. Biol. 2008, 15, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Markin, C.J.; Xiao, W.; Spyracopoulos, L. Mechanism for recognition of polyubiquitin chains: Balancing affinity through interplay between multivalent binding and dynamics. J. Am. Chem. Soc. 2010, 132, 11247–11258. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, K.; Zhang, X.; Tang, W.; Wang, J.; Guo, M.; Zhao, Q.; Zheng, X.; Wang, P.; Zhang, Z. Two phosphodiesterase genes, PDEL and PDEH, regulate development and pathogenicity by modulating intracellular cyclic AMP levels in Magnaporthe oryzae. PLoS ONE 2011, 6, e17241. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Sun, T.; Li, N.; Peng, J.; Fu, D.; Li, W.; Li, L.; Gao, W.Q. BRG1 attenuates colonic inflammation and tumorigenesis through autophagy-dependent oxidative stress sequestration. Nat. Commun. 2019, 10, 4614. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Shen, Y.; Xiong, Y.; Wang, S.; Li, C.; Bai, J. Zhang, Y. Loss of SMARCB1 promotes autophagy and facilitates tumour progression in chordoma by transcriptionally activating ATG5. Cell Prolif. 2021, 54, e13136. [Google Scholar] [CrossRef]
- Cheong, H.; Klionsky, D.J. Biochemical methods to monitor autophagy-related processes in yeast. Methods Enzymol. 2008, 451, 1–26. [Google Scholar] [CrossRef]
- Liu, X.H.; Chen, S.M.; Gao, H.M.; Ning, G.A.; Shi, H.B.; Wang, Y.; Dong, B.; Qi, Y.Y.; Zhang, D.M.; Lu, G.D.; et al. The small GTPase MoYpt7 is required for membrane fusion in autophagy and pathogenicity of Magnaporthe oryzae. Environ. Microbiol. 2015, 17, 4495–4510. [Google Scholar] [CrossRef]
- Wilson, C.J.; Chao, D.M.; Imbalzano, A.N.; Schnitzler, G.R.; Kingston, R.E.; Young, R.A. RNA polymerase II holoenzyme contains SWI/SNF regulators involved in chromatin remodeling. Cell 1996, 84, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, M.; Park, J.H.; Ahn, J.H.; Lee, Y.H. MoSNF1 regulates sporulation and pathogenicity in the rice blast fungus Magnaporthe Oryzae. Fungal Genet. Biol. 2008, 45, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, M.J.; Talbot, N.J. Genome-wide functional analysis reveals that infection-associated fungal autophagy is necessary for rice blast disease. Proc. Natl. Acad. Sci. USA 2009, 106, 15967–15972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.H.; Gao, H.M.; Xu, F.; Lu, J.P.; Devenish, R.J.; Lin, F.C. Autophagy vitalizes the pathogenicity of pathogenic fungi. Autophagy 2012, 8, 1415–1425. [Google Scholar] [CrossRef]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.Y.; Talbot, N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.-W.; Zhao, R.; Xu, X.-Z.; Tang, L.; Shi, W.; Chen, D.; Peng, J.-B.; Bhadauria, V.; Zhao, W.-S.; Yang, J.; et al. MoSnf5 Regulates Fungal Virulence, Growth, and Conidiation in Magnaporthe oryzae. J. Fungi 2023, 9, 18. https://doi.org/10.3390/jof9010018
Xu X-W, Zhao R, Xu X-Z, Tang L, Shi W, Chen D, Peng J-B, Bhadauria V, Zhao W-S, Yang J, et al. MoSnf5 Regulates Fungal Virulence, Growth, and Conidiation in Magnaporthe oryzae. Journal of Fungi. 2023; 9(1):18. https://doi.org/10.3390/jof9010018
Chicago/Turabian StyleXu, Xiao-Wen, Rui Zhao, Xiao-Zhou Xu, Liu Tang, Wei Shi, Deng Chen, Jun-Bo Peng, Vijai Bhadauria, Wen-Sheng Zhao, Jun Yang, and et al. 2023. "MoSnf5 Regulates Fungal Virulence, Growth, and Conidiation in Magnaporthe oryzae" Journal of Fungi 9, no. 1: 18. https://doi.org/10.3390/jof9010018