
Exploring Potent Fungal Isolates from Sanitary Landfill Soil for In Vitro Degradation of Dibutyl Phthalate
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Enrichment and Isolation of DBP Degrading Fungi
2.3. Morphological Characterization of Isolates
2.4. DBP Biodegradation
2.5. Determination of Fungal Biomass
2.6. Determination of Proteins and Esterase Production
2.7. DBP Extraction and Analysis
2.8. Molecular Identification of SDBP4
2.9. Statistical Analysis
3. Results
3.1. Isolation of DBP Degrading Fungi
3.2. Biomass, Proteins and Esterase Production
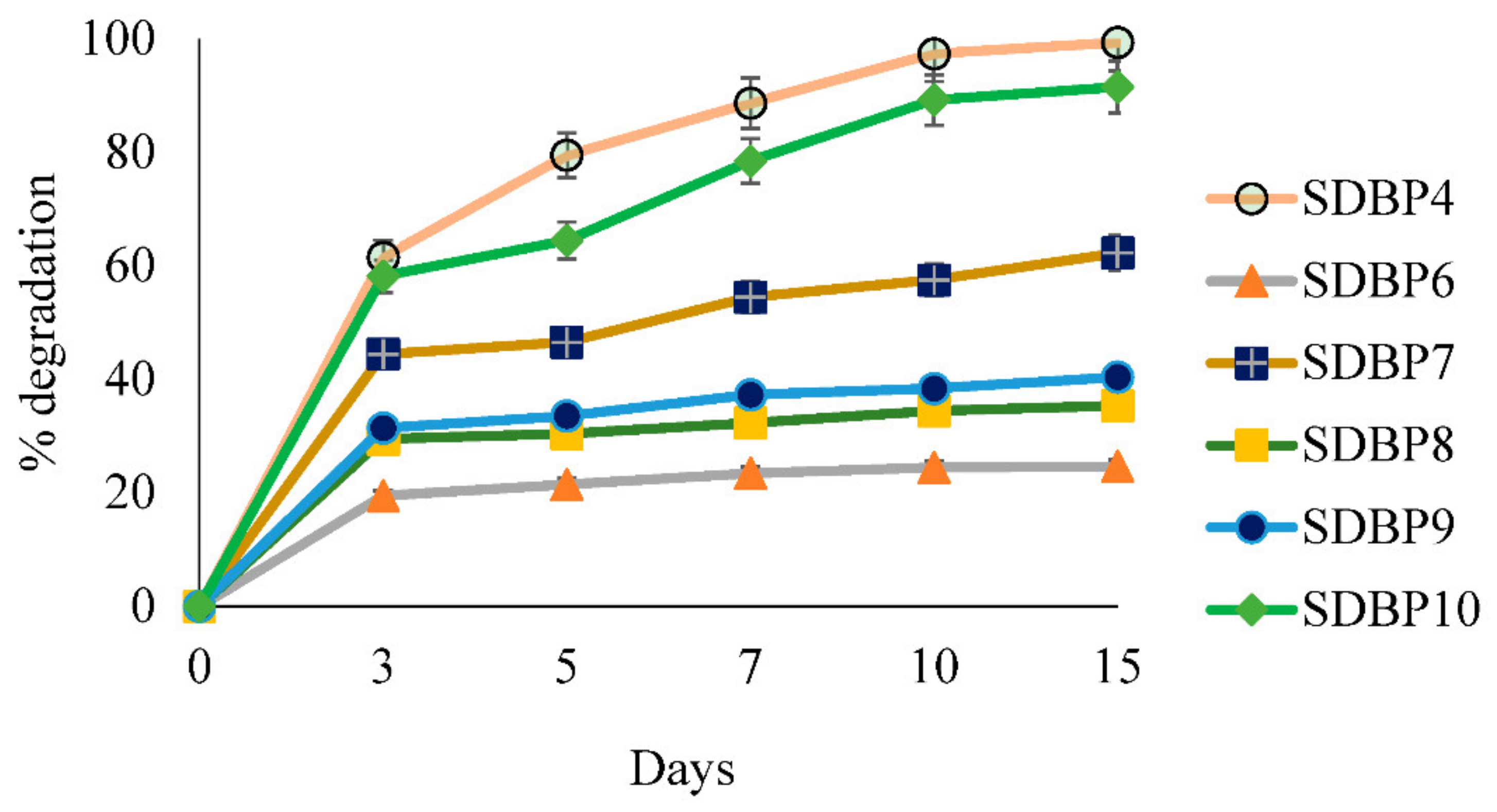
3.3. Biodegradation of DBP
3.4. Molecular Identification of SDBP4
3.5. Metabolites and Proposed Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IARC (International Agency for Research on Cancer). Some Industrial Chemicals. Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 2000; p. 77. [Google Scholar]
- Net, S.; Sempere, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Technol. 2015, 49, 4019–4035. [Google Scholar] [CrossRef] [PubMed]
- Allied Market Research Market Outlook. Available online: https://www.alliedmarketresearch.com/phthalic-anhydride-market-A12103 (accessed on 24 October 2021).
- Gani, K.M.; Kazmi, A.A. Phthalate contamination in aquatic environment: A critical review of the process factors that influence their removal in conventional and advanced wastewater treatment. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1402–1439. [Google Scholar] [CrossRef]
- Meeker, J.D.; Sathyanarayana, S.; Swan, S.H. Phthalates and other additives in plastics: Human exposure and associated health outcomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2097–2113. [Google Scholar] [CrossRef] [Green Version]
- Zolfaghari, M.; Drogui, P.; Seyhi, B.; Brar, S.K.; Buelna, G.; Dubé, R. Occurrence, fate and effects of Di (2-ethylhexyl) phthalate in wastewater treatment plants: A review. Environ. Poll. 2014, 194, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Gosetti, F.; Bolfi, B.; Robotti, E.; Manfredi, M.; Binotti, M.; Ferrero, F.; Bona, G.; Marengo, E. Study of endocrine disrupting compound release from different medical devices through an on-line SPE UHPLC-MS/MS method. Anal. Chim. Acta 2018, 1042, 141–154. [Google Scholar] [CrossRef]
- Lee, Y.M.; Lee, J.E.; Choe, W.; Kim, T.; Lee, J.Y.; Kho, Y.; Choi, K.; Zoh, K.D. Distribution of phthalate esters in air, water, sediments, and fish in the Asan Lake of Korea. Environ. Int. 2019, 126, 635–643. [Google Scholar] [CrossRef]
- Wang, R.; Ji, M.; Zhai, H.; Liu, Y. Occurrence of phthalate esters and microplastics in urban secondary effluents, receiving water bodies and reclaimed water treatment processes. Sci. Total Environ. 2020, 737, 140219. [Google Scholar] [CrossRef]
- Basak, S.; Das, M.K.; Duttaroy, A.K. Plastics derived endocrine-disrupting compounds and their effects on early development. Birth. Defects Res. 2020, 112, 1308–1325. [Google Scholar] [CrossRef]
- Kong, X.; Jin, D.; Jin, S.; Wang, Z.; Yin, H.; Xu, M.; Deng, Y. Responses of bacterial community to dibutyl phthalate pollution in a soil-vegetable ecosystem. J. Hazard. Mater. 2018, 353, 142–150. [Google Scholar] [CrossRef]
- Wowkonowicz, P.; Kijeńska, M. Phthalate release in leachate from municipal landfills of central Poland. PLoS ONE 2017, 12, e0174986. [Google Scholar] [CrossRef]
- Benjamin, S.; Masai, E.; Kamimura, N.; Takahashi, K.; Anderson, R.C.; Faisal, P.A. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J. Hazard. Mater. 2017, 340, 360–383. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Wang, Z.; Chen, J.; Zhang, L.; Cai, Z.; Wu, Y.; Ma, P. Role of the necrosis factor-κB pathway in dibutyl phthalate mediated effects on human glioma cells. Toxicol. Environ. Chem. 2018, 100, 644–657. [Google Scholar] [CrossRef]
- Consumer Product Safety Improvement Act. Public Law 110–314. Available online: https://www.cpsc.gov/s3fs-public/cpsia.pdf (accessed on 24 October 2021).
- Hoyer, P.B. Reproductive toxicology: Current and future directions. Biochem. Pharmacol. 2001, 62, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Arzuaga, X.; Walker, T.; Yost, E.E.; Radke, E.G.; Hotchkiss, A.K. Use of the Adverse Outcome Pathway (AOP) framework to evaluate species concordance and human relevance of Dibutyl phthalate (DBP)-induced male reproductive toxicity. Reprod. Toxicol. 2020, 96, 445–458. [Google Scholar] [CrossRef]
- Liang, D.W.; Zhang, T.; Fang, H.H.; He, J. Phthalates biodegradation in the environment. Appl. Microbiol. Biotechnol. 2008, 80, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Kaneco, S.; Katsumata, H.; Suzuki, T.; Ohta, K. Titanium dioxide mediated photocatalytic degradation of dibutyl phthalate in aqueous solution—Kinetics, mineralization and reaction mechanism. Chem. Eng. J. 2006, 125, 59–66. [Google Scholar] [CrossRef]
- Benjamin, S.; Pradeep, S.; Josh, M.S.; Kumar, S.; Masai, E. A monograph on the remediation of hazardous phthalates. J. Hazard. Mater. 2015, 298, 58–72. [Google Scholar] [CrossRef]
- Zhu, Z.; Rao, R.; Zhao, Z.; Chen, J.; Jiang, W.; Bi, F.; Yang, Y.; Zhang, X. Research progress on removal of phthalates pollutants from environment. J. Mol. Liq. 2022, 355, 118930. [Google Scholar] [CrossRef]
- Pradeep, S.; Faseela, P.; Josh, M.K.; Balachandran, S.; Devi, R.S.; Benjamin, S. Fungal biodegradation of phthalate plasticizer in situ. Biodegradation 2013, 24, 257–267. [Google Scholar] [CrossRef]
- Córdoba-Sosa, G.; Torres, J.L.; Ahuactzin-Pérez, M.; Díaz-Godínez, G.; Díaz, R.; Sánchez, C. Growth of Pleurotus ostreatus ATCC 3526 in different concentrations of di (2-ethylhexyl) phthalate in submerged fermentation. J. Chem. Biol. Phys. Sci. 2014, 4, 96. [Google Scholar]
- Gao, J.; Chi, J. Biodegradation of phthalate acid esters by different marine microalgal species. Mar. Pollut. Bull. 2015, 99, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Niu, G.; Yang, W.; Cao, X. Di (2-ethylhexyl) phthalate biodegradation and denitrification by a Pseudoxanthomonas sp. strain. Biores. Technol. 2015, 180, 356–359. [Google Scholar] [CrossRef]
- Ahuactzin-Pérez, M.; Tlecuitl-Beristain, S.; García-Dávila, J.; González-Pérez, M.; Gutiérrez-Ruíz, M.C.; Sánchez, C. Degradation of di (2-ethyl hexyl) phthalate by Fusarium culmorum: Kinetics, enzymatic activities and biodegradation pathway based on quantum chemical modelling pathway based on quantum chemical modelling. Sci. Total Environ. 2016, 1, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Jia, Y.; Ruth, N.; Qiao, C.; Wang, J.; Zhao, B.; Yan, Y. Biodegradation of phthalic acid esters by a newly isolated Mycobacterium sp. YC-RL4 and the bioprocess with environmental samples. Environ. Sci. Pollut. Res. 2016, 23, 16609–16619. [Google Scholar] [CrossRef] [PubMed]
- Patil, N.K.; Kundapur, R.; Shouche, Y.S.; Karegoudar, T.B. Degradation of a plasticizer, di-n-butylphthalate by Delftia sp. TBKNP-05. Curr. Microbiol. 2006, 52, 225–230. [Google Scholar] [CrossRef]
- Fang, C.R.; Yao, J.; Zheng, Y.G.; Jiang, C.J.; Hu, L.F.; Wu, Y.Y.; Shen, D.S. Dibutyl phthalate degradation by Enterobacter sp. T5 isolated from municipal solid waste in landfill bioreactor. Int. Biodeterior. Biodegrad. 2010, 64, 442–446. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Y.; Liang, R.; Dai, Q.; Jin, D.; Chao, W. Biodegradation of an endocrine-disrupting chemical di-n-butyl phthalate by newly isolated Agrobacterium sp. and the biochemical pathway. Process Biochem. 2011, 46, 1090–1094. [Google Scholar] [CrossRef]
- Prasad, B.; Suresh, S. Biodegradation of dimethyl phthalate, diethyl phthalate, dibutyl phthalate and their mixture by Variovorax sp. Int. J. Environ. Sci. Dev. 2012, 3, 283. [Google Scholar] [CrossRef]
- Hu, J.; Yang, Q.; Wang, J.L. Biodegradation of di-n-butyl phthalate in sequencing batch reactor bioaugmented with Micrococcus sp. and the bacterial community analysis. Int. J. Environ. Sci. Technol. 2015, 12, 2819–2828. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Kong, X.; Liu, H.; Wang, X.; Deng, Y.; Jia, M.; Yu, X. Characterization and genomic analysis of a highly efficient dibutyl phthalate-degrading bacterium Gordonia sp. strain QH-12. Int. J. Mol. Sci. 2016, 17, 1012. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Maitra, S.S. Biodegradation of endocrine disruptor dibutyl phthalate (DBP) by a newly isolated Methylobacillus sp. V29b and the DBP degradation pathway. 3 Biotech 2016, 6, 200. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.J.; Zhang, L.S.; Fang, Y.; Zhou, Y.; Ye, B.C. Biodegradation of phthalate esters by newly isolated Rhizobium sp. LMB-1 and its biochemical pathway of di-n-butyl phthalate. J. App. Microbiol. 2016, 121, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.J.; Wan, Q.; Wang, W.F.; Wang, Y.; Feng, F.Y.; Cheng, J.J.; Yuan, J.J.; Yu, X.Y. Biodegradation of dibutyl phthalate by a novel endophytic Bacillus subtilis strain HB-T2 under in-vitro and in-vivo conditions. Environ. Technol. 2020, 28, 1917–1926. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, Y.; Jayathandar, R.S.; Dhayalan, S.; Subramaniyan, S.; Punamalai, G. Biodegradation of di-(2-ethylhexyl) phthalate by novel Rhodococcus sp. PFS1 strain isolated from paddy field soil. Arch. Microbiol. 2021, 204, 21. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, J.W.; Koo, B.W.; Kim, M.K.; Choi, D.H.; Choi, I.G. Dibutyl phthalate biodegradation by the white rot fungus, Polyporus brumalis. Biotechnol. Bioeng. 2007, 97, 1516–1522. [Google Scholar] [CrossRef]
- Ahuactzin-Pérez, M.; Tlecuitl-Beristain, S.; García-Dávila, J.; Santacruz-Juárez, E.; González-Pérez, M.; Gutiérrez-Ruíz, M.C.; Sánchez, C. Mineralization of high concentrations of the endocrine disruptor dibutyl phthalate by Fusarium culmorum. 3 Biotech 2018, 8, 42. [Google Scholar] [CrossRef]
- Ahuactzin-Pérez, M.; Tlecuitl-Beristain, S.; García-Dávila, J.; Santacruz-Juárez, E.; González-Pérez, M.; Gutiérrez-Ruíz, M.C.; Sánchez, C. Kinetics and pathway of biodegradation of dibutyl phthalate by Pleurotus ostreatus. Fungal Biol. 2018, 122, 991–997. [Google Scholar] [CrossRef]
- Carstens, L.; Cowan, A.R.; Seiwert, B.; Schlosser, D. Biotransformation of phthalate plasticizers and bisphenol A by marine-derived, freshwater, and terrestrial fungi. Front. Microbiol. 2020, 11, 317. [Google Scholar] [CrossRef]
- González-Márquez, A.; Volke-Sepulveda, T.; Díaz, R.; Sánchez, C. Enhanced esterase activity during the degradation of dibutyl phthalate by Fusarium species in liquid fermentation. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab062. [Google Scholar] [CrossRef]
- Abd El-Rahim, W.M.; Mostafa, E.M.; Moawad, H. High cell density cultivation of six fungal strains efficient in azo dye bioremediation. Biotechnol. Rep. 2016, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Keerthiraj, M.; Mandal, A.; Dutta, T.K.; Saha, S.; Dutta, A.; Singh, A.; Kundu, A. Nematicidal and molecular docking investigation of essential oils from Pogostemon cablin ecotypes against Meloidogyne incognita. Chem. Biodivers. 2021, 18, e2100320. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Xu, W.; Zhong, T.H.; He, G.Y.; Luo, Z.H. Degradation of dimethyl phthalate esters by a filamentous fungus Aspergillus versicolor isolated from deep-sea sediments. Bot. Mar. 2017, 60, 351–359. [Google Scholar] [CrossRef]
- Sheoran, O.P.; Tonk, D.S.; Kaushik, L.S.; Hasija, R.C.; Pannu, R.S. Statistical software package for agricultural research workers. In Recent Advances in Information Theory, Statistics and Computer Applications; Hooda, D.S., Hasija, R.C., Eds.; Department of Mathematics Statistics, CCS HAU: Hisar, India, 1998; pp. 139–143. [Google Scholar]
- Feng, L.; Liu, H.; Cheng, D.; Mao, X.; Wang, Y.; Wu, Z.; Wu, Q. Characterization and genome analysis of a phthalate esters-degrading strain Sphingobium yanoikuyae SHJ. BioMed Res. Int. 2018, 2018, 3917054. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xiao, H.; Tang, L.; Min, H.; Lu, Z. Biodegradation of di-n-butyl phthalate by a stable bacterial consortium, HD-1, enriched from activated sludge. Biores. Technol. 2013, 128, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.M.; Hu, R.W.; Chen, X.X.; Chen, X.B.; Lü, H.; Li, Y.W.; Li, H.; Mo, C.H.; Cai, Q.Y.; Wong, M.H. Biodegradation pathway of di-(2-ethylhexyl) phthalate by a novel Rhodococcus pyridinivorans XB and its bioaugmentation for remediation of DEHP contaminated soil. Sci. Total Environ. 2018, 640, 1121–1131. [Google Scholar] [CrossRef]
- Lee, S.M.; Koo, B.W.; Lee, S.S.; Kim, M.K.; Choi, D.H.; Hong, E.J.; Jeung, E.B.; Choi, I.G. Biodegradation of dibutyl phthalate by white rot fungi and evaluation on its estrogenic activity. Enzyme Microb. Technol. 2004, 35, 417–423. [Google Scholar] [CrossRef]
- Huang, L.; Meng, D.; Tian, Q.; Yang, S.; Deng, H.; Guan, Z.; Cai, Y.; Liao, X. Characterization of a novel carboxylesterase from Bacillus velezensis SYBC H47 and its application in degradation of phthalate esters. J. Biosci. Bioeng. 2020, 129, 588–594. [Google Scholar] [CrossRef]
- Zhao, H.M.; Du, H.; Lin, J.; Chen, X.B.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H.; Qin, H.M.; Wong, M.H. Complete degradation of the endocrine disruptor di-(2-ethylhexyl) phthalate by a novel Agromyces sp. MT-O strain and its application to bioremediation of contaminated soil. Sci. Total Environ. 2016, 562, 170–178. [Google Scholar] [CrossRef]
- Feng, N.X.; Feng, Y.X.; Liang, Q.F.; Chen, X.; Xiang, L.; Zhao, H.M.; Liu, B.L.; Cao, G.; Li, Y.W.; Li, H.; et al. Complete biodegradation of di-n-butyl phthalate (DBP) by a novel Pseudomonas sp. YJB6. Sci. Total Environ. 2021, 761, 143208. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, N.; Maitra, S.S. Comparative study on the degradation of dibutyl phthalate by two newly isolated Pseudomonas sp. V21b and Comamonas sp. 51F. Biotechnol. Rep. 2017, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.D.; Li, J.X.; Wang, Y.Y. Biochemical pathway and degradation of phthalate ester isomers by bacteria. Water Sci. Technol. 2005, 52, 241–248. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, A.T.; Ribeiro, M.A.; Pinho, C.F.; Peixoto, A.R.; Domeniconi, R.F.; Scarano, W.R. Regulatory and junctional proteins of the blood-testis barrier in human Sertoli cells are modified by monobutyl phthalate (MBP) and bisphenol A (BPA) exposure. Toxicol. In Vitro 2016, 34, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mittermeier, A.; Völkel, W.; Fromme, H. Kinetics of the phthalate metabolites mono-2-ethylhexyl phthalate (MEHP) and mono-n-butyl phthalate (MnBP) in male subjects after a single oral dose. Toxicol. Lett. 2016, 252, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Nomura, Y.; Nakagawa, M.; Ogawa, N.; Harashima, S.; Oshima, Y. Genes in PHT plasmid encoding the initial degradation pathway of phthalate in Pseudomonas putida. J. Ferment. Bioeng. 1992, 74, 333–344. [Google Scholar] [CrossRef]
- Zhao, H.M.; Du, H.; Feng, N.X.; Xiang, L.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H. Biodegradation of di-n-butylphthalate and phthalic acid by a novel Providencia sp. 2D and its stimulation in a compost-amended soil. Biol. Fertil. Soils 2016, 52, 65–76. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, J.; Ahn, J.Y.; Gu, M.B.; Moon, S.H. Enhanced degradation of an endocrine-disrupting chemical, butyl benzyl phthalate, by Fusarium oxysporum f. sp. pisi cutinase. Appl. Environ. Microbiol. 2002, 68, 4684–4688. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liang, R.; Dai, Q.; Jin, D.; Wang, Y.; Chao, W. Complete degradation of di-n-octyl phthalate by biochemical cooperation between Gordonia sp. strain JDC-2 and Arthrobacter sp. strain JDC-32 isolated from activated sludge. J. Hazard. Mater. 2010, 176, 262–268. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Colony Characteristics | Sporulation | Spore Colour |
|---|---|---|---|
| SDBP4 | Raised, grainy, filiform | Yes | Light green |
| SDBP6 | Circular, raised, entire | No | - |
| SDBP7 | Irregular, flat, filiform | Yes | Dark green |
| SDBP8 | Irregular, grainy, filiform | Yes | Dull green |
| SDBP9 | Raised, Irregular, filiform | Yes | Bright dark green |
| SDBP10 | Raised, powdery, filiform | Yes | Dull green |
| Rt (min) | Compound | Molecular Ion Peak (m/z) | Base Peak (m/z) | Fragment Ions | ||
|---|---|---|---|---|---|---|
| RIlit | next | |||||
| 7.78 | DMP | 1466 | 1465 | 194 | 163 | 194, 149, 135, 104, 77 |
| 12.33 | PA | 1836 | 1839 | 250 | 163 | 181, 149, 135, 92, 77 |
| 15.79 | DIP | 1871 | 1873 | 278 | 149 | 223, 205, 167, 160, 132, 104, 93 |
| 17.54 | DBP | 1967 | 1969 | 278 | 149 | 223, 205, 167, 160, 121, 104, 93 |
| 25.53 | BBP | 2356 | 2359 | 312 | 149 | 295, 238, 206, 178, 123, 91, 65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puranik, S.; Shukla, L.; Kundu, A.; Kamil, D.; Paul, S.; Venkadasamy, G.; Salim, R.; Singh, S.K.; Kumar, D.; Kumar, A. Exploring Potent Fungal Isolates from Sanitary Landfill Soil for In Vitro Degradation of Dibutyl Phthalate. J. Fungi 2023, 9, 125. https://doi.org/10.3390/jof9010125
Puranik S, Shukla L, Kundu A, Kamil D, Paul S, Venkadasamy G, Salim R, Singh SK, Kumar D, Kumar A. Exploring Potent Fungal Isolates from Sanitary Landfill Soil for In Vitro Degradation of Dibutyl Phthalate. Journal of Fungi. 2023; 9(1):125. https://doi.org/10.3390/jof9010125
Chicago/Turabian StylePuranik, Shriniketan, Livleen Shukla, Aditi Kundu, Deeba Kamil, Sangeeta Paul, Govindasamy Venkadasamy, Rajna Salim, Sandeep Kumar Singh, Dharmendra Kumar, and Ajay Kumar. 2023. "Exploring Potent Fungal Isolates from Sanitary Landfill Soil for In Vitro Degradation of Dibutyl Phthalate" Journal of Fungi 9, no. 1: 125. https://doi.org/10.3390/jof9010125