
CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain, Media, and Culture Conditions
2.2. Construction of Editing Plasmids and DNA Manipulation
2.3. Protoplast Preparation and Transformation
2.4. Editing Efficiency Statistics
2.5. Colony PCR
2.6. Determination of the Growth Curve of A. oryzae
2.7. Protein Expression and Validation
2.8. Determination of Lipase Enzyme Activity
2.9. Determination of α-Amylase Enzyme Activity
3. Results
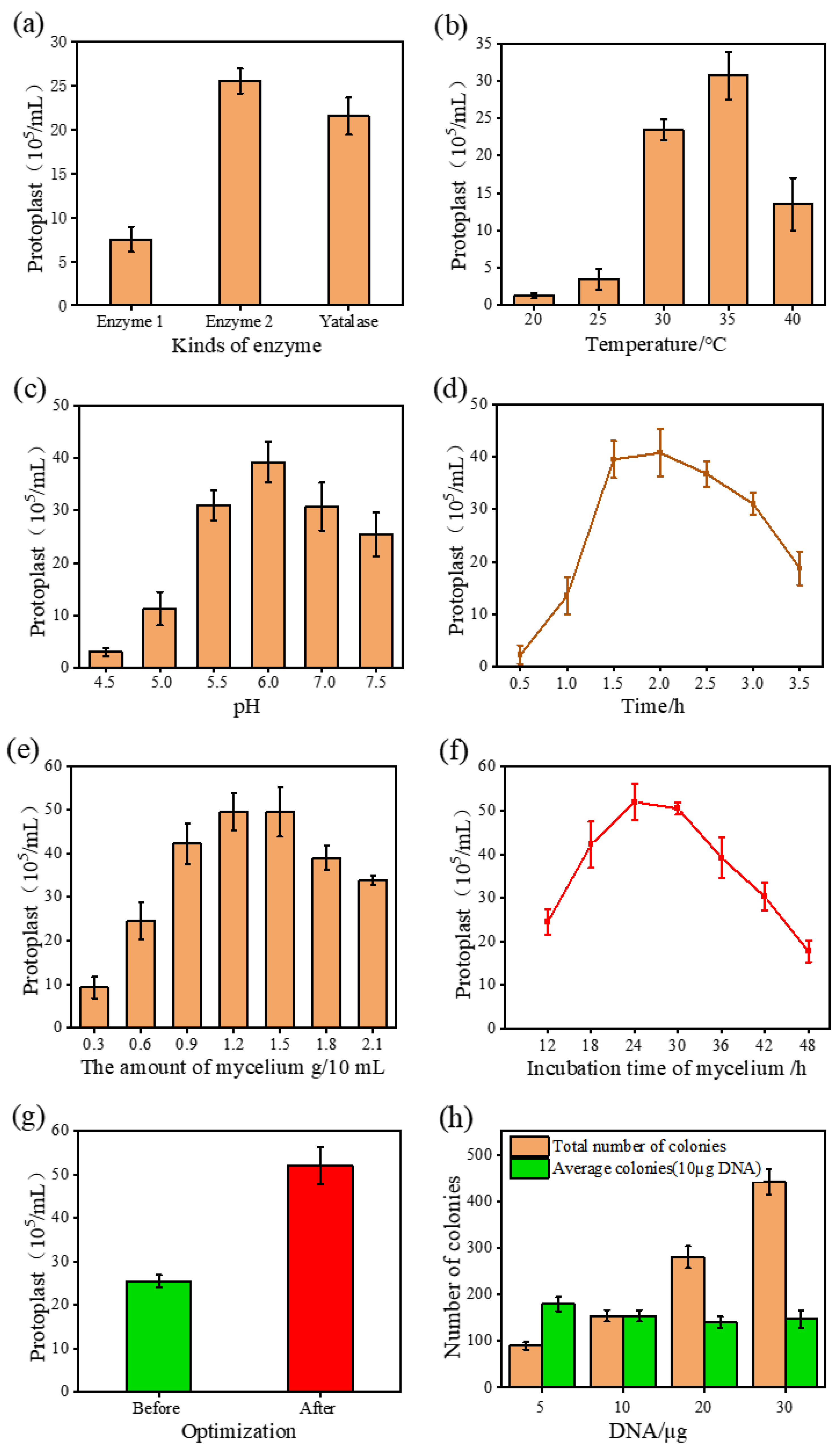
3.1. Optimization of the Genetic Transformation
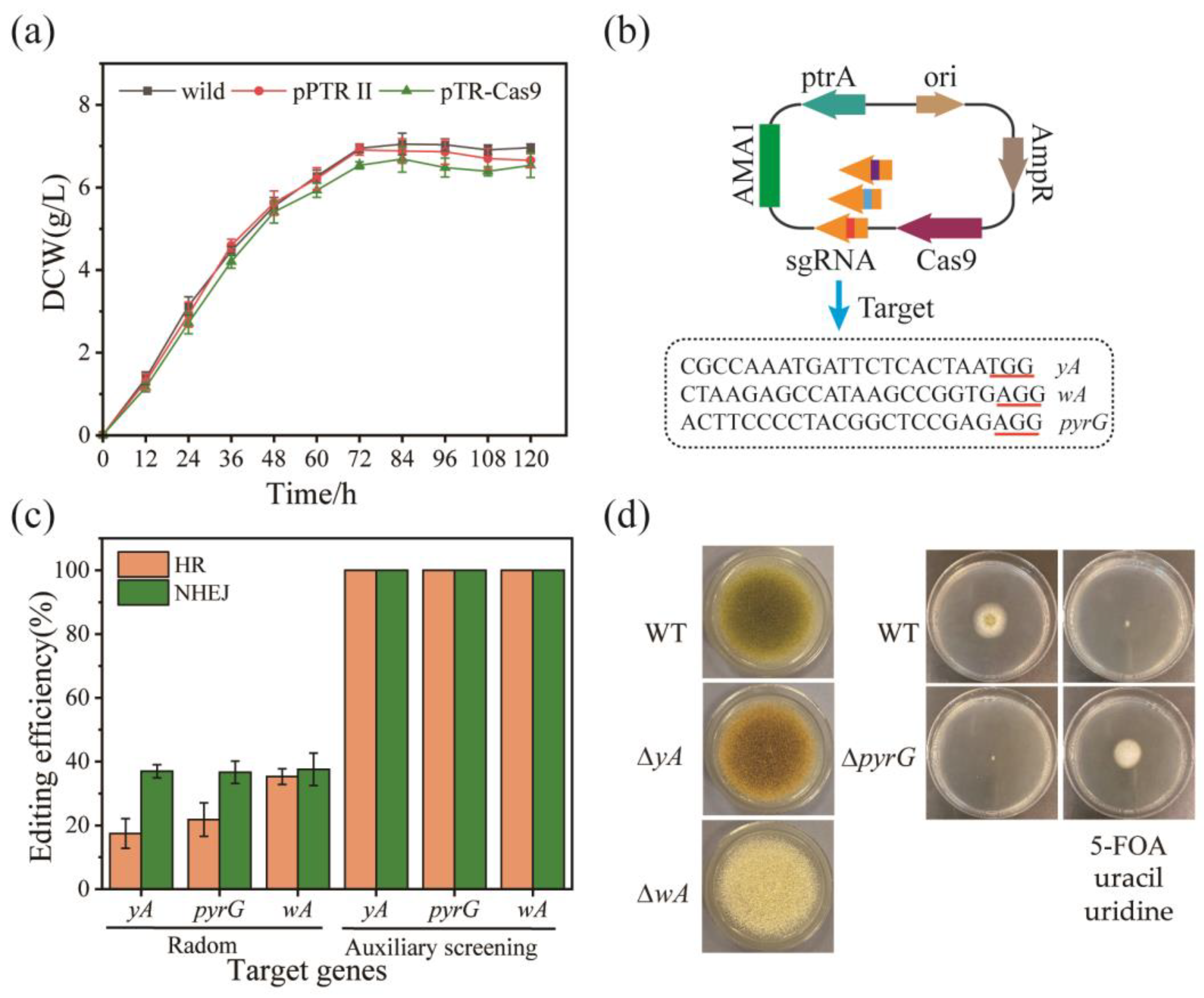
3.2. Single-Gene Editing
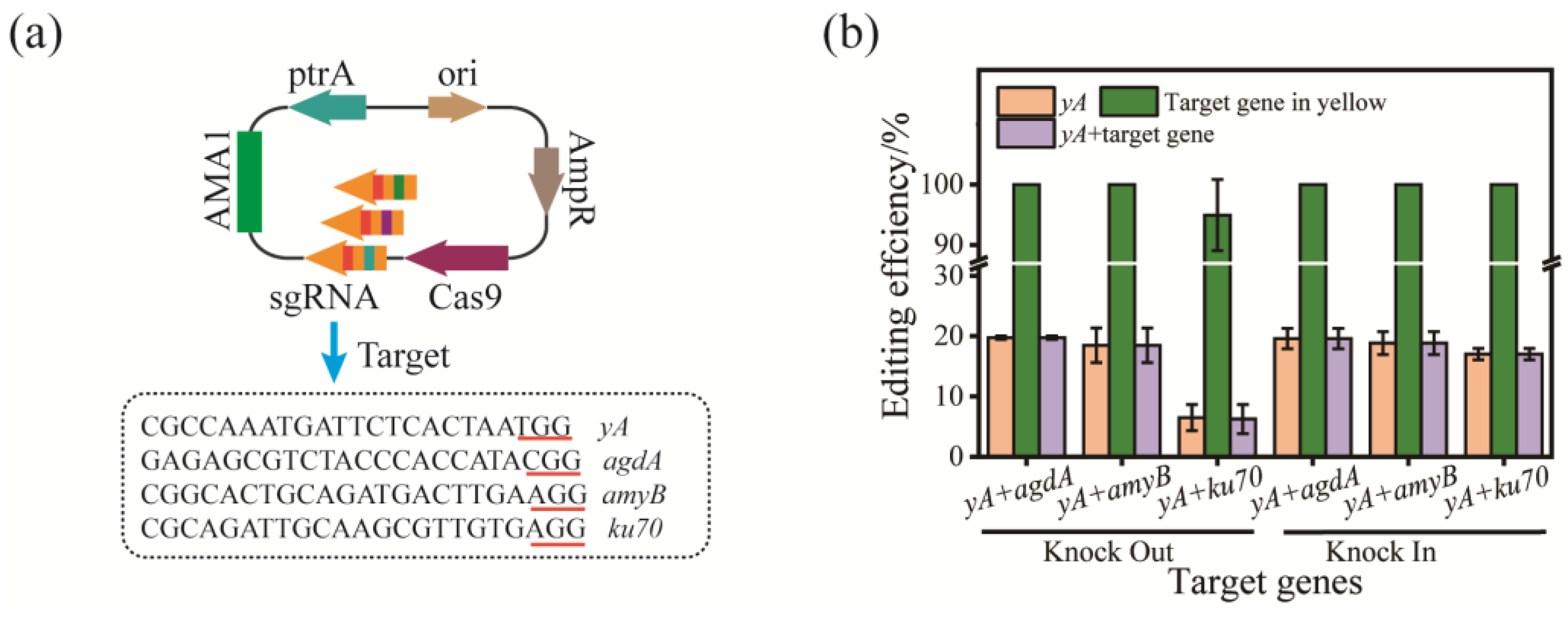
3.3. Dual Gene Editing
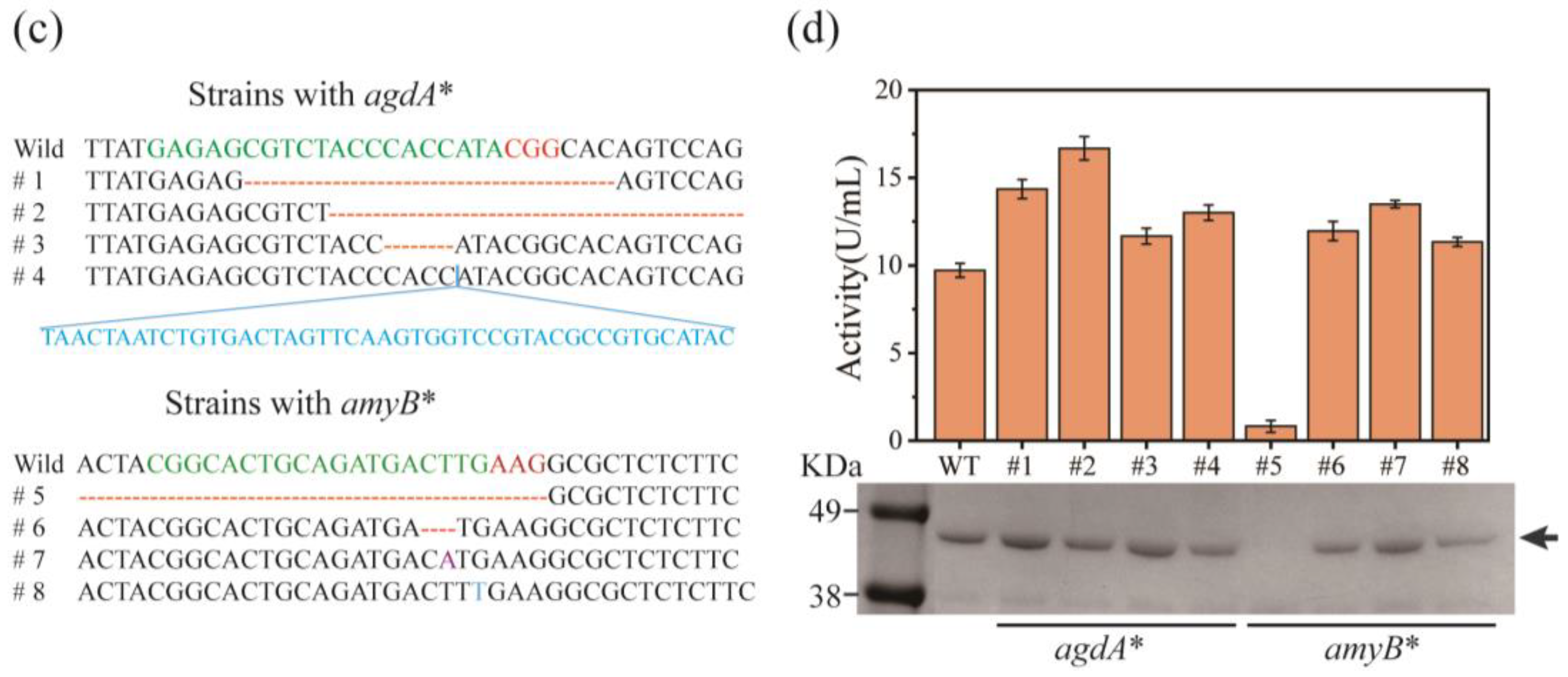
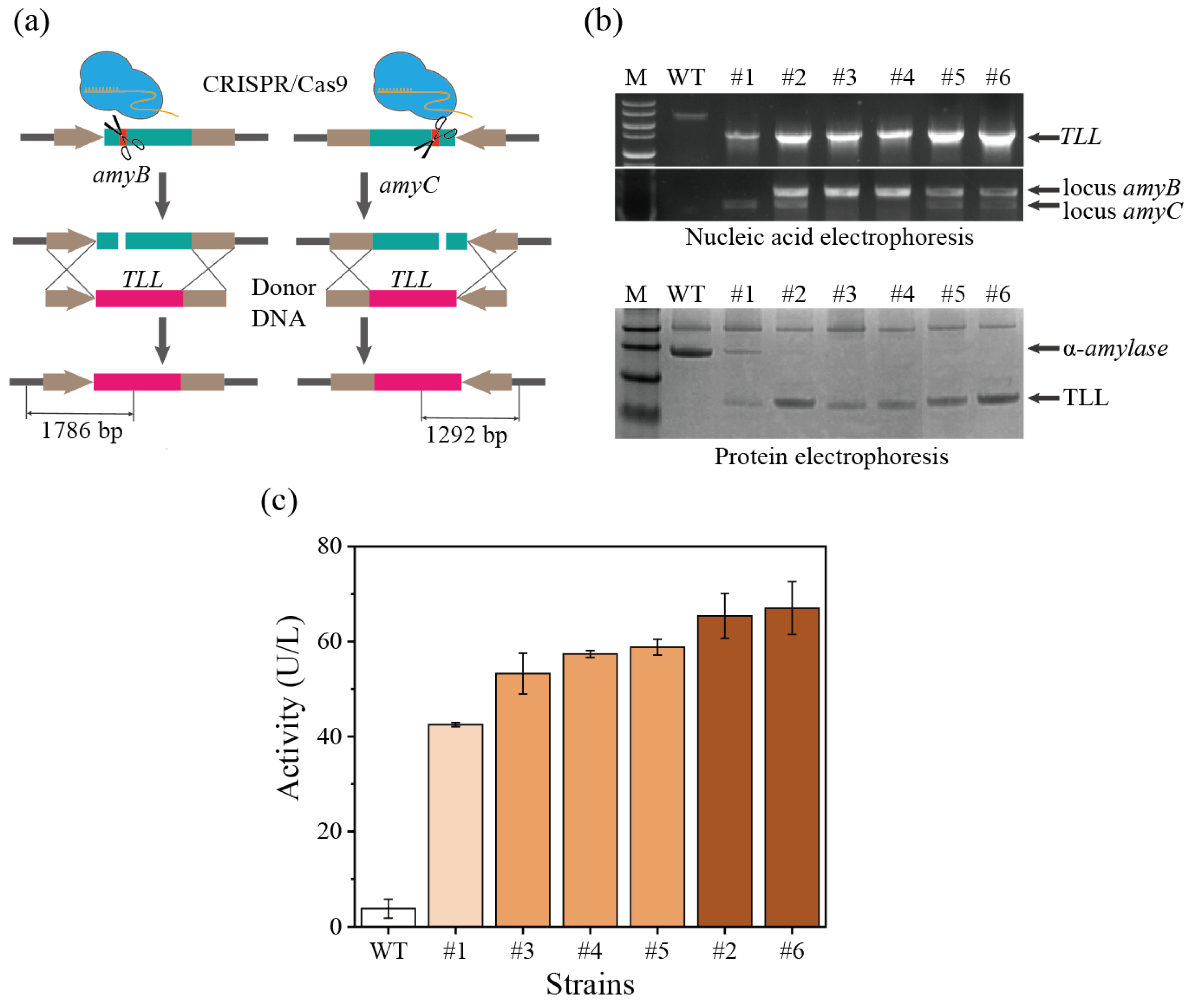
3.4. CRISPR/Cas9-Mediated Multiplexed Site Integration in A. oryzae for Heterologous Lipase Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, C.; Meng, M.; Jiang, Y.; Hou, L. Improvement of the quality of soy sauce by reducing enzyme activity in Aspergillus oryzae. Food Chem. 2019, 292, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Møller, L.L.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wösten, H.A.B. Filamentous fungi for the production of enzymes, chemicals and materials. Curr. Opin. Biotechnol. 2019, 59, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, J.; Zhang, G.; Liu, S.; Zhou, J.; Du, G.; Chen, J. Recent advances in the development of Aspergillus for protein production. Bioresour. Technol. 2022, 348, 126768. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Liu, G.; Ji, R.; Shi, K.; Song, P.; Ren, L.; Huang, H.; Ji, X. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef]
- Jiang, D.; Zhu, W.; Wang, Y.; Sun, C.; Zhang, K.; Yang, J. Molecular tools for functional genomics in filamentous fungi: Recent advances and new strategies. Biotechnol. Adv. 2013, 31, 1562–1574. [Google Scholar] [CrossRef]
- Maruyama, J.-i.; Nakajima, H.; Kitamoto, K. Visualization of nuclei in Aspergillus oryzae with EGFP and analysis of the number of nuclei in each conidium by FACS. Biosci. Biotechnol. Biochem. 2001, 65, 1504–1510. [Google Scholar] [CrossRef]
- Maruyama, J. Genome editing technology and its application potentials in the industrial filamentous fungus Aspergillus oryzae. J. Fungi 2021, 7, 638. [Google Scholar] [CrossRef]
- McCarty, N.S.; Graham, A.E.; Studená, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation. Nat. Commun. 2020, 11, 1281. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Xu, J.; Zuo, Y.; Ye, C.; Jiang, L.; Feng, L.; Huang, L.; Xu, Z.; Lian, J. Synthetic Biology Toolkit for Marker-Less Integration of Multigene Pathways into Pichia pastoris via CRISPR/Cas9. ACS Synth. Biol. 2022, 11, 623–633. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, L.; Xu, Y.; Jiang, W.; Shi, T.; Sun, X.; Huang, H. Recent advances in the application of multiplex genome editing in Saccharomyces cerevisiae. Appl. Microbiol. 2021, 105, 3873–3882. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Coleman, J.J. Progress and challenges: Development and implementation of CRISPR/Cas9 technology in filamentous fungi. Comput. Struct. Biotechnol. J. 2019, 17, 761–769. [Google Scholar] [CrossRef]
- Song, R.; Zhai, Q.; Sun, L.; Huang, E.; Zhang, Y.; Zhu, Y.; Guo, Q.; Tian, Y.; Zhao, B.; Lu, H. CRISPR/Cas9 genome editing technology in filamentous fungi: Progress and perspective. Appl. Microbiol. 2019, 103, 6919–6932. [Google Scholar] [CrossRef] [Green Version]
- Katayama, T.; Nakamura, H.; Zhang, Y.; Pascal, A.; Wataru, F.; Maruyama, J. Forced recycling of an AMA1-based genome-editing plasmid allows for efficient multiple gene deletion/integration in the industrial filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2019, 85, e01896-18. [Google Scholar] [CrossRef] [Green Version]
- Nødvig, C.S.; Hoof, J.B.; Kogle, M.E.; Jarczynska, Z.D.; Lehmbeck, J.; Klitgaard, D.K.; Mortensen, U.H. Efficient oligo nucleotide mediated CRISPR-Cas9 gene editing in Aspergilli. Fungal Genet. Biol. 2018, 115, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, J.; Wang, Z.; Zhang, Y.; Shi, S.; Nielsen, J.; Liu, Z. A gRNA-tRNA array for CRISPR-Cas9 based rapid multiplexed genome editing in Saccharomyces cerevisiae. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Collias, D.; Vialetto, E.; Yu, J.; Almasi, E.; Ruettiger, A.-S.; Achmedov, T.; Strowig, T.; Beisel, C.L. Systematically attenuating DNA targeting enables CRISPR-driven editing in bacteria. bioRxiv 2022. [Google Scholar] [CrossRef]
- Li, C.; Zhou, J.; Rao, S.; Du, G.; Liu, S. Visualized multigene editing system for Aspergillus niger. ACS Synth. Biol. 2021, 10, 2607–2616. [Google Scholar] [CrossRef]
- Reeves, E.P.; Messina, C.; Doyle, S.; Kavanagh, K. Correlation between gliotoxin production and virulence of Aspergillus fumigatus in Galleria mellonella. Mycopathologia 2004, 158, 73–79. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, M.; Yan, X.; Zhang, G.; Xia, J.; Zeng, G.; Tian, W.; Bao, X.; Zeng, Z.; Yu, P. Expression and characterization of a novel lipase from Bacillus licheniformis NCU CS-5 for application in enhancing fatty acids flavor release for low-fat cheeses. Food Chem. 2022, 368, 130868. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, Y.; Li, Q.; Zhou, J.; Li, J.; Du, G. Growth-coupled evolution and high-throughput screening assisted rapid enhancement for amylase-producing Bacillus licheniformis. Bioresour. Technol. 2021, 337, 125467. [Google Scholar] [CrossRef]
- Jin, F.; Hu, S.; Wang, B.; Jin, L. Advances in genetic engineering technology and its application in the industrial fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef]
- Szewczyk, E.; Nayak, T.; Oakley, C.E.; Edgerton, H.; Xiong, Y.; Taheri-Talesh, N.; Osmani, S.A.; Oakley, B.R. Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 2006, 1, 3111–3120. [Google Scholar] [CrossRef]
- Seetharam, M.C.; Gargi, B.; Megha, S.; Surbhi, W.; Savita, K.; Satyendra, G.; Rita, M.; Rath, D. Determination of Cas9/dCas9 associated toxicity in microbes. bioRxiv 2019, 848135. [Google Scholar] [CrossRef] [Green Version]
- Nemoto, T.; Maruyama, J.; Kitamoto, K. Contribution ratios of amyA, amyB, amyC genes to high-level α-amylase expression in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2012, 76, 1477–1483. [Google Scholar] [CrossRef]
- Katsuya, G. Regulatory mechanisms for amylolytic gene expression in the koji mold Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2019, 83, 1385–1401. [Google Scholar]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Li, M.; Xiao, Z.; Hu, M.; Matsuda, Y.; Wang, W. Recent advances in heterologous production of natural products using Aspergillus oryzae. Synth. Biol. J. 2022, 3, 1–24. [Google Scholar] [CrossRef]
- Ntana, F.; Uffe, H.M.; Sarazin, C.; Figge, R. Aspergillus: A powerful protein production platform. Catalysts 2020, 10, 1064. [Google Scholar] [CrossRef]
- Zhang, X.; Gu, S.; Zheng, X.; Peng, S.; Li, Y.; Lin, Y.; Liang, S. A Novel and Efficient Genome Editing Tool Assisted by CRISPR-Cas12a/Cpf1 for Pichia pastoris. ACS Synth. Biol. 2021, 10, 2927–2937. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Gao, R.; Li, J.; Lin, L.; Zhao, J.; Sun, W.; Tian, C. Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering. Biotechnol. Biofuels 2017, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanka, F.; Arentshorst, M.; Cairns, T.C.; Jørgensen, T.; Ram, A.F.; Meyer, V. Highly active promoters and native secretion signals for protein production during extremely low growth rates in Aspergillus niger. Microb. Cell Fact. 2016, 15, 145. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Lu, J.; Zhang, G.; Zhou, J.; Li, J.; Du, G.; Chen, J. CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae. J. Fungi 2023, 9, 109. https://doi.org/10.3390/jof9010109
Li Q, Lu J, Zhang G, Zhou J, Li J, Du G, Chen J. CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae. Journal of Fungi. 2023; 9(1):109. https://doi.org/10.3390/jof9010109
Chicago/Turabian StyleLi, Qinghua, Jinchang Lu, Guoqiang Zhang, Jingwen Zhou, Jianghua Li, Guocheng Du, and Jian Chen. 2023. "CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae" Journal of Fungi 9, no. 1: 109. https://doi.org/10.3390/jof9010109