
The Adaptor Protein UvSte50 Governs Fungal Pathogenicity of Ustilaginoidea virens via the MAPK Signaling Pathway

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Construction of Gene Deletion and Complementation Mutants
2.3. Analysis of Mycelial Growth and Conidiation
2.4. Virulence and Plant Infection Assays
2.5. Stress Adaptation Assays
2.6. RT-qPCR and Transcriptome Sequencing Analysis
2.7. Generation of Green Fluorescent Protein (GFP) Fusion Cassettes
2.8. Yeast Two-Hybrid Assay
2.9. Affinity Capture–Mass Spectrometry Analysis
3. Results
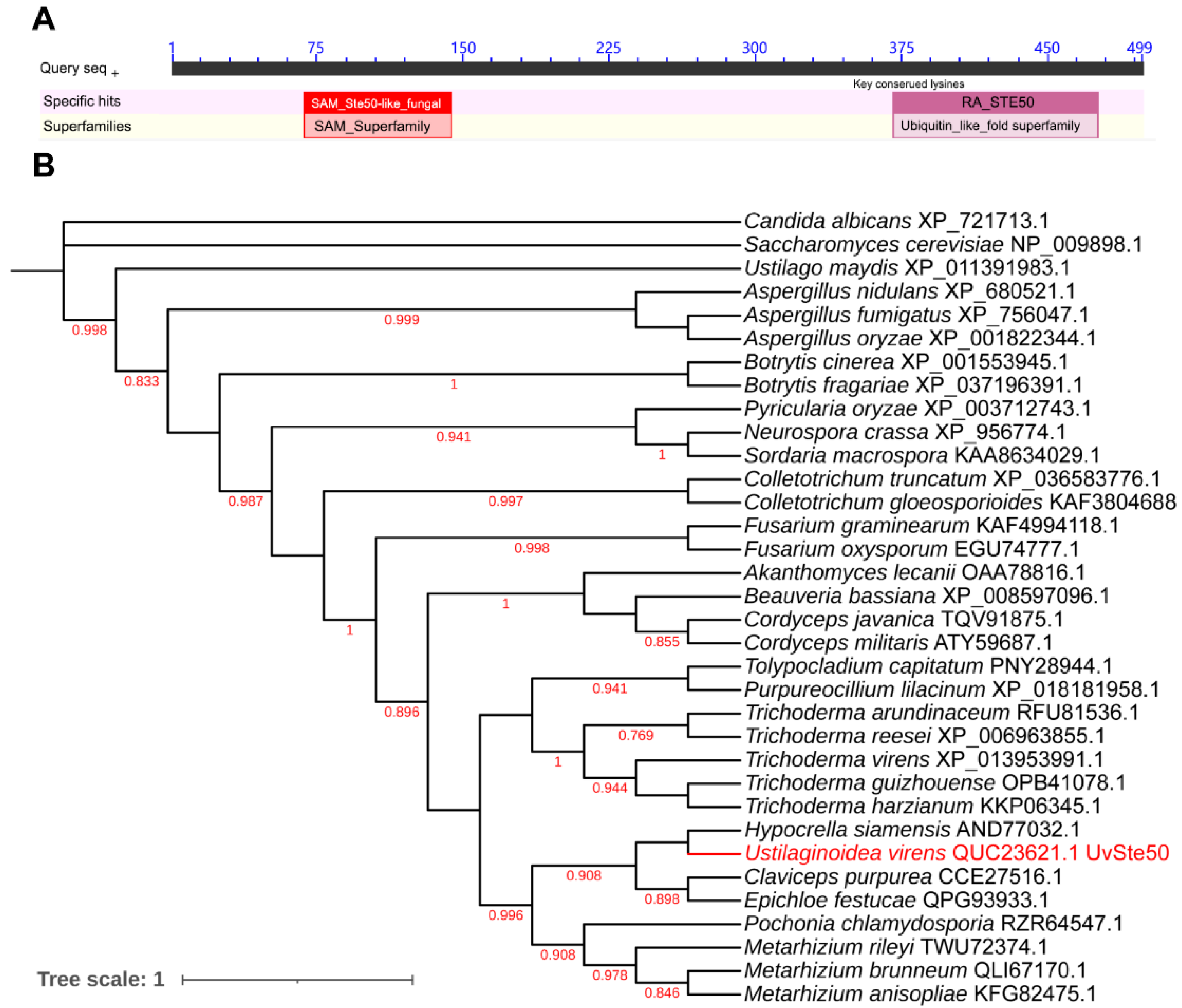
3.1. Identification of Ste50 Ortholog in U. Virens
3.2. UvSte50 Is Required for Conidiation in U. virens
3.3. UvSte50 Is Essential for Full Virulence in U. virens
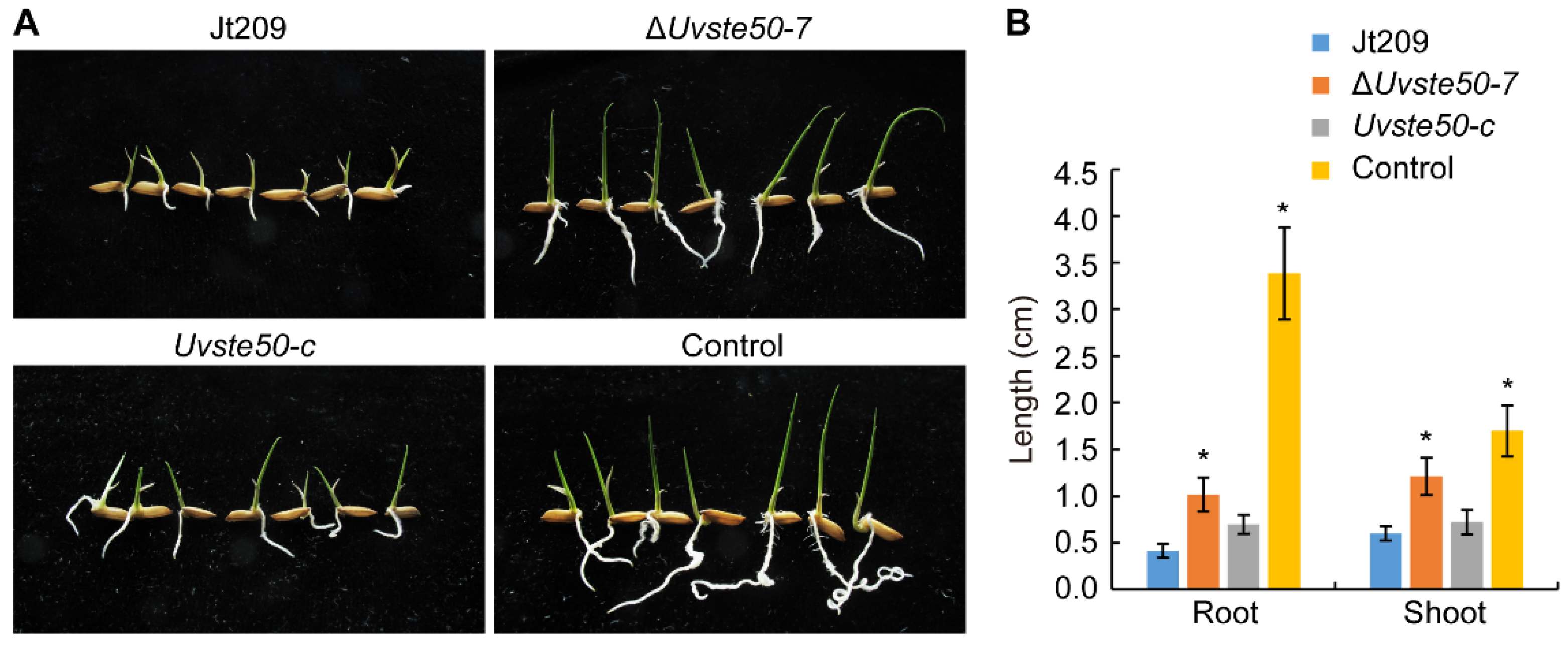
3.4. Culture Filtrates of ΔUvste50 Are Less Toxic to Rice Seed Germination
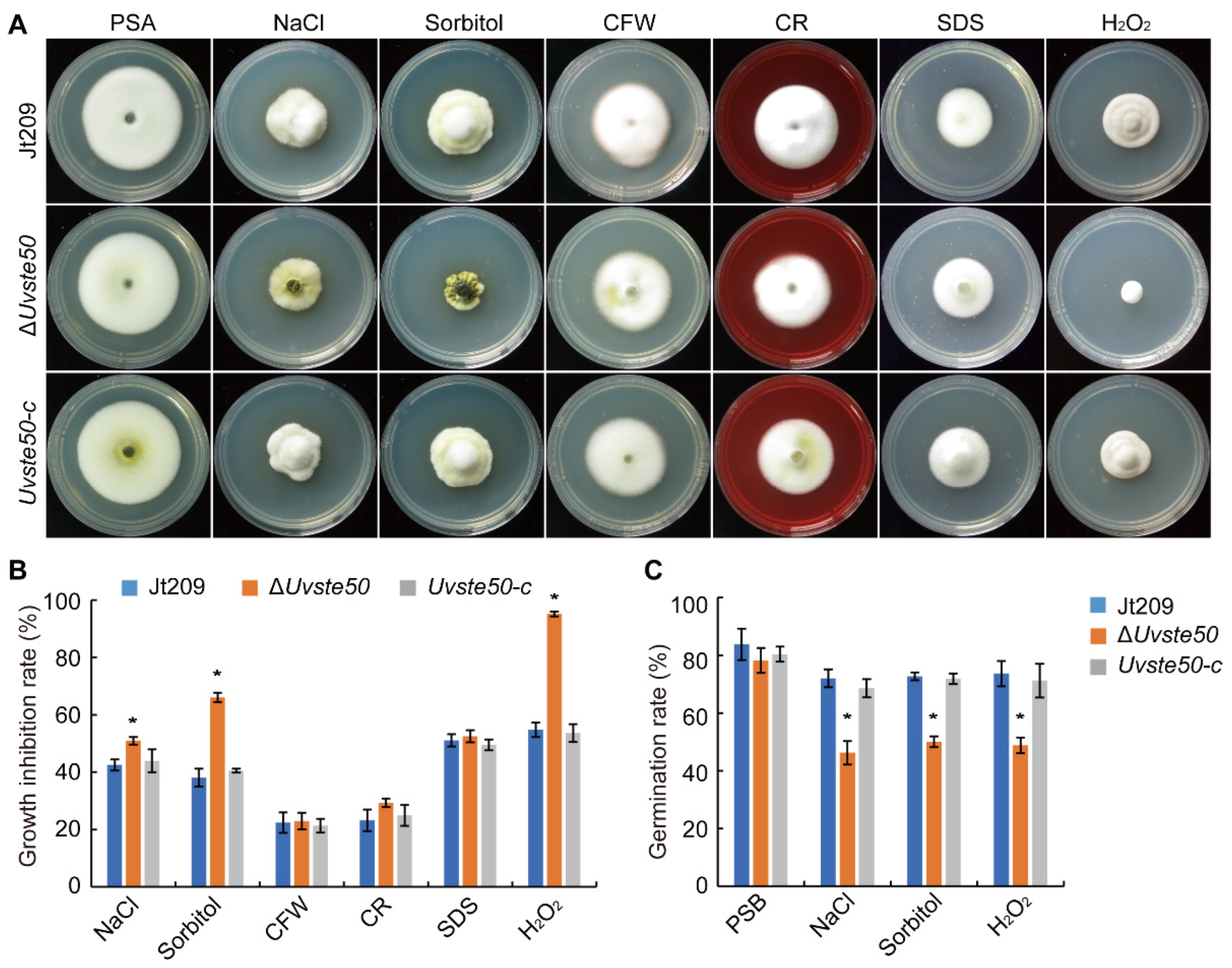
3.5. UvSTE50 Is Involved in Hyperosmotic and Oxidative Regulation in U. virens
3.6. Deletion of UvSTE50 Affects the Expression of Genes Involved in MAPK Signaling Pathways
3.7. Deletion of UvSTE50 Affects the Transcription of a Subset of Genes in U. virens
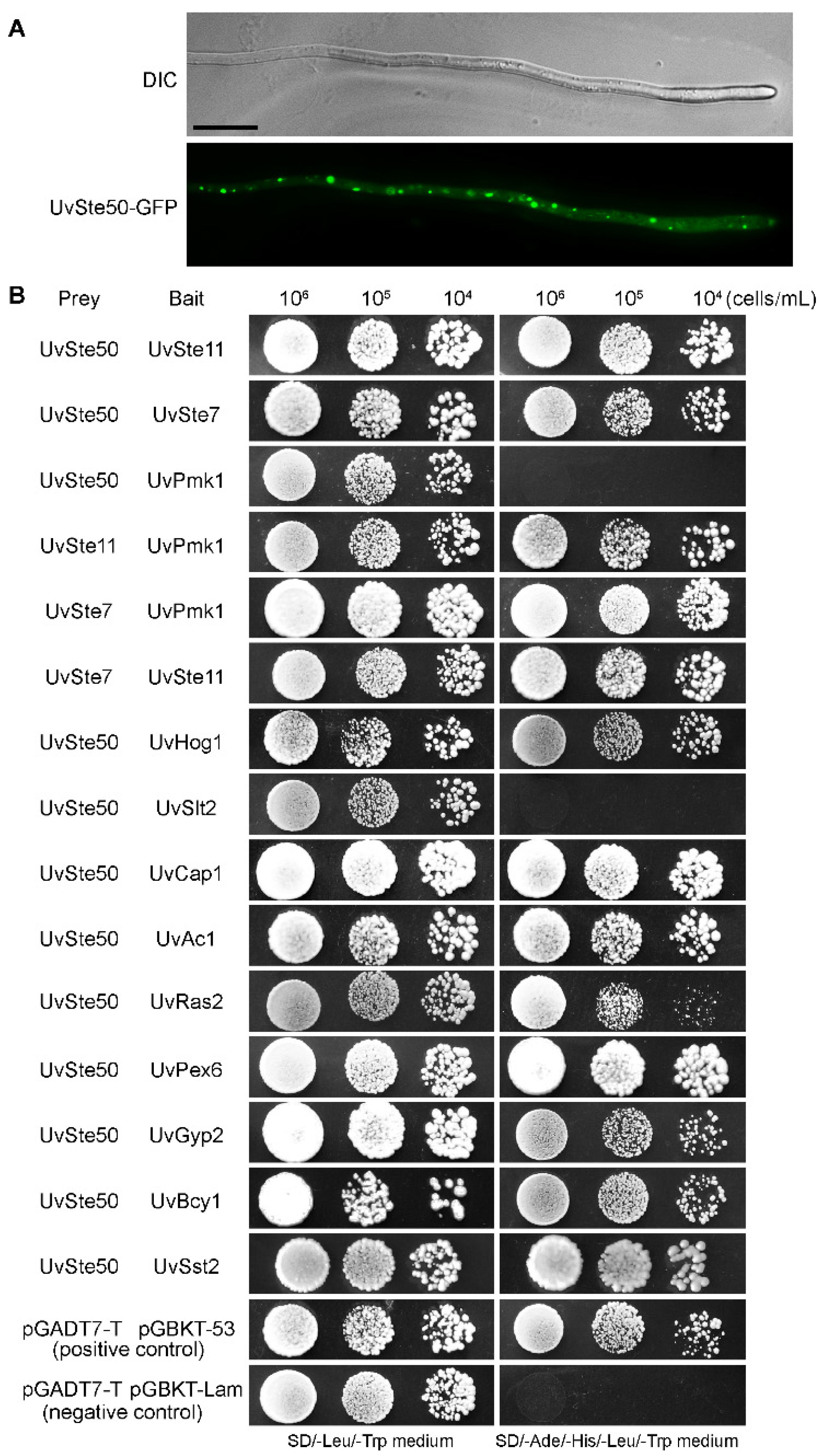
3.8. UvSte50 Was Distributed as Spots in the Cytoplasm of Hyphae
3.9. Protein–Protein Interaction Studies to Identify UvSte50-Interacting Partners
3.10. RA Domain, but Not SAM Domain, Is Essential for the Interaction of UvSte50 with UvSte7 and UvRas2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, Y.X.; Jin, J.; Hu, D.W.; Yong, M.L.; Xu, Y.; He, L.P. Elucidation of the infection process of Ustilaginoidea virens (teleomorph: Villosiclava virens) in rice spikelets. Plant Pathol. 2013, 62, 1–8. [Google Scholar] [CrossRef]
- Song, J.H.; Wei, W.; Lv, B.; Lin, Y.; Yin, W.X.; Peng, Y.L.; Schnabel, G.; Huang, J.B.; Jiang, D.H.; Luo, C.X. Rice false smut fungus hijacks the rice nutrients supply by blocking and mimicking the fertilization of rice ovary. Environ. Microbiol. 2016, 18, 3840–3849. [Google Scholar] [CrossRef]
- Sun, W.; Fan, J.; Fang, A.F.; Li, Y.J.; Tariqjaveed, M.; Li, D.Y.; Hu, D.W.; Wang, W.M. Ustilaginoidea virens: Insights into an emerging rice pathogen. Annu. Rev. Phytopathol. 2020, 58, 363–385. [Google Scholar] [CrossRef]
- Wang, B.; Liu, L.; Li, Y.J.; Zou, J.Y.; Li, D.Y.; Zhao, D.; Li, W.; Sun, W.X. Ustilaginoidin D induces hepatotoxicity and behaviour aberrations in zebrafish larvae. Toxicology 2021, 456, 152786. [Google Scholar] [CrossRef]
- Ji, F.; He, D.; Olaniran, A.O.; Mokoena, M.P.; Xu, J.; Shi, J. Occurrence, toxicity, production and detection of Fusarium mycotoxin: A review. Food Prod. Process. Nutr. 2019, 1, 6. [Google Scholar] [CrossRef]
- Kumar, A.; Pathak, H.; Bhadauria, S.; Sudan, J. Aflatoxin contamination in food crops: Causes, detection, and management: A review. Food Prod. Process. Nutr. 2021, 3, 17. [Google Scholar] [CrossRef]
- Fan, J.; Guo, X.Y.; Li, L.; Huang, F.; Sun, W.X.; Li, Y.; Huang, Y.Y.; Xu, Y.J.; Shi, J.; Lei, Y.; et al. Infection of Ustilaginoidea virens intercepts rice seed formation but activates grain-filling-related genes. J. Integr. Plant Biol. 2015, 57, 577–590. [Google Scholar] [CrossRef]
- Andrews, D.L.; Egan, J.D.; Mayorga, M.E.; Gold, S.E. The Ustilago maydis ubc4 and ubc5 genes encode members of a MAP kinase cascade required for filamentous growth. Mol. Plant Microbe Interact. 2000, 13, 781–786. [Google Scholar] [CrossRef]
- Jenczmionka, N.J.; Schafer, W. The Gpmk1 MAP kinase of Fusarium graminearum regulates the induction of specific secreted enzymes. Curr. Genet. 2005, 47, 29–36. [Google Scholar] [CrossRef]
- Li, G.T.; Zhou, X.Y.; Xu, J.R. Genetic control of infection-related development in Magnaporthe oryzae. Curr. Opin. Microbiol. 2012, 15, 678–684. [Google Scholar] [CrossRef]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [CrossRef]
- Xu, J.R. MAP kinases in fungal pathogens. Fungal Genet. Biol. 2000, 31, 137–152. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef]
- Zhao, X.H.; Xu, J.R. A highly conserved MAPK-docking site in Mst7 is essential for Pmk1 activation in Magnaporthe grisea. Mol. Microbiol. 2007, 63, 881–894. [Google Scholar] [CrossRef]
- Chen, R.E.; Thorner, J. Function and regulation in MAPK signaling pathways: Lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 1311–1340. [Google Scholar] [CrossRef]
- Hamel, L.P.; Nicole, M.C.; Duplessis, S.; Ellis, B.E. Mitogen-activated protein kinase signaling in plant-interacting fungi: Distinct messages from conserved messengers. Plant Cell 2012, 24, 1327–1351. [Google Scholar] [CrossRef]
- Tang, J.T.; Bai, J.; Chen, X.Y.; Zheng, L.; Liu, H.; Huang, J.B. Two protein kinases UvPmk1 and UvCDC2 with significant functions in conidiation, stress response and pathogenicity of rice false smut fungus Ustilaginoidea virens. Curr. Genet. 2020, 66, 409–420. [Google Scholar] [CrossRef]
- Liang, Y.F.; Han, Y.; Wang, C.F.; Jiang, C.; Xu, J.R. Targeted deletion of the USTA and UvSLT2 genes efficiently in Ustilaginoidea virens with the CRISPR-Cas9 system. Front. Plant Sci. 2018, 9, 699. [Google Scholar] [CrossRef]
- Chen, X.Y.; Li, P.P.; Liu, H.; Chen, X.L.; Huang, J.B.; Luo, C.X.; Li, G.T.; Hsiang, T.; Collinge, D.B.; Zheng, L. A novel transcription factor UvCGBP1 regulates development and virulence of rice false smut fungus Ustilaginoidea virens. Virulence 2021, 12, 1563–1579. [Google Scholar] [CrossRef]
- Zheng, D.W.; Wang, Y.; Han, Y.; Xu, J.R.; Wang, C.F. UvHOG1 is important for hyphal growth and stress responses in the rice false smut fungus Ustilaginoidea virens. Sci. Rep. 2016, 6, 24824. [Google Scholar] [CrossRef]
- Ramezani-Rad, M. The role of adaptor protein Ste50-dependent regulation of the MAPKKK Ste11 in multiple signalling pathways of yeast. Curr. Genet. 2003, 43, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Poplinski, A.; Hopp, C.; Ramezani-Rad, M. Ste50 adaptor protein influences Ras/cAMP-driven stress-response and cell survival in Saccharomyces cerevisiae. Curr. Genet. 2007, 51, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Mares, C.; Lizcano, A.; Liu, Y.; Wickes, B.L. Insertional mutagenesis combined with an inducible filamentation phenotype reveals a conserved STE50 homologue in Cryptococcus neoformans that is required for monokaryotic fruiting and sexual reproduction. Mol. Microbiol. 2011, 79, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Zhao, X.H.; Snyder, A.K.; Xu, J.R.; Shah, D.M. Two mitogen-activated protein kinase signalling cascades mediate basal resistance to antifungal plant defensins in Fusarium graminearum. Cell. Microbiol. 2007, 9, 1491–1506. [Google Scholar] [CrossRef]
- Gu, Q.; Chen, Y.; Liu, Y.; Zhang, C.Q.; Ma, Z.H. The transmembrane protein FgSho1 regulates fungal development and pathogenicity via the MAPK module Ste50-Ste11-Ste7 in Fusarium graminearum. New Phytol. 2015, 206, 315–328. [Google Scholar] [CrossRef]
- Park, G.; Xue, C.; Zhao, X.; Kim, Y.; Orbach, M.; Xu, J.R. Multiple upstream signals converge on the adaptor protein Mst50 in Magnaporthe grisea. Plant Cell 2006, 18, 2822–2835. [Google Scholar] [CrossRef]
- Mayorga, M.E.; Gold, S.E. The ubc2 gene of Ustilago maydis encodes a putative novel adaptor protein required for filamentous growth, pheromone response and virulence. Mol. Microbiol. 2001, 41, 1365–1379. [Google Scholar] [CrossRef]
- Schamber, A.; Leroch, M.; Diwo, J.; Mendgen, K.; Hahn, M. The role of mitogen-activated protein (MAP) kinase signalling components and the Ste12 transcription factor in germination and pathogenicity of Botrytis cinerea. Mol. Plant Pathol. 2010, 11, 105–119. [Google Scholar] [CrossRef]
- Yu, M.; Yu, J.; Cao, H.; Pan, X.; Song, T.; Qi, Z.; Du, Y.; Huang, S.; Liu, Y. The velvet protein UvVEA regulates conidiation and chlamydospore formation in Ustilaginoidea virens. J. Fungi 2022, 8, 479. [Google Scholar] [CrossRef]
- Yu, M.N.; Yu, J.J.; Hu, J.K.; Huang, L.; Wang, Y.H.; Yin, X.L.; Nie, Y.F.; Meng, X.K.; Wang, W.D.; Liu, Y.F. Identification of pathogenicity-related genes in the rice pathogen Ustilaginoidea virens through random insertional mutagenesis. Fungal Genet. Biol. 2015, 76, 10–19. [Google Scholar] [CrossRef]
- Cao, H.J.; Zhang, J.J.; Yong, M.L.; Yu, M.N.; Song, T.Q.; Yuv, J.J.; Pan, X.Y.; Liu, Y.F. The cyclase-associated protein UvCap1 is required for mycelial growth and pathogenicity in the rice false smut fungus. Phytopathol. Res. 2021, 3, 5. [Google Scholar] [CrossRef]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Zheng, L.; Liu, H.; Tang, J.T.; Hsiang, T.; Huang, J.B. Use of random T-DNA mutagenesis in identification of gene UvPRO1, a regulator of conidiation, stress response, and virulence in Ustilaginoidea virens. Front. Microbiol. 2016, 7, 2086. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Hai, D.; Tang, J.T.; Liu, H.; Huang, J.B.; Luo, C.X.; Hsiang, T.; Zheng, L. UvCom1 is an important regulator required for development and infection in the rice false smut fungus Ustilaginoidea virens. Phytopathology 2020, 110, 483–493. [Google Scholar] [CrossRef]
- Xie, S.L.; Wang, X.F.; Wei, W.; Li, C.Y.; Liu, Y.; Qu, J.S.; Meng, Q.H.; Lin, Y.; Yin, W.X.; Yang, Y.N.; et al. The Bax inhibitor UvBI-1, a negative regulator of mycelial growth and conidiation, mediates stress response and is critical for pathogenicity of the rice false smut fungus Ustilaginoidea virens. Curr. Genet. 2019, 65, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Yong, M.L.; Yu, J.J.; Pan, X.Y.; Yu, M.N.; Cao, H.J.; Qi, Z.Q.; Du, Y.; Zhang, R.S.; Song, T.Q.; Yin, X.L.; et al. MAT1-1-3, a mating type gene in the Villosiclava virens, is required for fruiting bodies and sclerotia formation, asexual development and pathogenicity. Front. Microbiol. 2020, 11, 1337. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, K.; Fang, A.F.; Han, Y.Q.; Yang, J.; Xue, M.F.; Bao, J.D.; Hu, D.W.; Zhou, B.; Sun, X.Y.; et al. Specific adaptation of Ustilaginoidea virens in occupying host florets revealed by comparative and functional genomics. Nat. Commun. 2014, 5, 3849. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenfuhrer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar] [CrossRef]
- Ren, J.; Zhang, Y.; Wang, Y.; Li, C.; Bian, Z.; Zhang, X.; Liu, H.; Xu, J.-R.; Jiang, C. Deletion of all three MAP kinase genes results in severe defects in stress responses and pathogenesis in Fusarium graminearum. Stress Biol. 2022, 2, 6. [Google Scholar] [CrossRef]
- Yan, X.; Li, Y.; Yue, X.F.; Wang, C.C.; Que, Y.W.; Kong, D.D.; Ma, Z.H.; Talbot, N.J.; Wang, Z.Y. Two novel transcriptional regulators are essential for infection-related morphogenesis and pathogenicity of the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002385. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.W.; Gao, Y.X.; Yu, Z.M.; Xiao, Y.H.; Zhang, Z.G.; Zhang, H.F. The adenylate cyclase UvAc1 and phosphodiesterase UvPdeH control the intracellular cAMP level, development, and pathogenicity of the rice false smut fungus Ustilaginoidea virens. Fungal Genet. Biol. 2019, 129, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zhang, X.; Liu, H.Q.; Xu, J.R. Mitogen-activated protein kinase signaling in plant pathogenic fungi. PLoS Pathog. 2018, 14, e1006875. [Google Scholar] [CrossRef] [PubMed]
- Osés-Ruiz, M.; Cruz-Mireles, N.; Martin-Urdiroz, M.; Soanes, D.M.; Eseola, A.B.; Tang, B.; Derbyshire, P.; Nielsen, M.; Cheema, J.; Were, V.; et al. Appressorium-mediated plant infection by Magnaporthe oryzae is regulated by a Pmk1-dependent hierarchical transcriptional network. Nat. Microbiol. 2021, 6, 1383–1397. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Z.; Jiang, C.; Xu, J.-R. Regulation of biotic interactions and responses to abiotic stresses by MAP kinase pathways in plant pathogenic fungi. Stress Biol. 2021, 1, 5. [Google Scholar] [CrossRef]
- Ko, Y.J.; Yu, Y.M.; Kim, G.B.; Lee, G.W.; Maeng, P.J.; Kim, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. 2009. Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways. Eukaryot. Cell 2009, 8, 1197–1217. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.W.; Kim, S.Y.; Okagaki, L.H.; Nielsen, K.; Bahn, Y.S. Ste50 adaptor protein governs sexual differentiation of Cryptococcus neoformans via the pheromone-response MAPK signaling pathway. Fungal Genet. Biol. 2011, 48, 154–165. [Google Scholar] [CrossRef]
- Rad, M.R.; Jansen, G.; Buhring, F.; Hollenberg, C.P. Ste50p is involved in regulating filamentous growth in the yeast Saccharomyces cerevisiae and associates with Ste11p. Mol. Gen. Genet. 1998, 259, 29–38. [Google Scholar] [CrossRef]
- Lee, B.N.; Elion, E.A. The MAPKKK Ste11 regulates vegetative growth through a kinase cascade of shared signaling components. Proc. Natl. Acad Sci. USA 1999, 96, 12679–12684. [Google Scholar] [CrossRef]
- O’Rourke, S.M.; Herskowitz, I. The Hog1 MAPK prevents cross talk between the HOG and pheromone response MAPK pathways in Saccharomyces cerevisiae. Genes Dev. 1998, 12, 2874–2886. [Google Scholar] [CrossRef]
- Posas, F.; Witten, E.A.; Saito, H. Requirement of STE50 for osmostress-induced activation of the STE11 mitogen-activated protein kinase kinase kinase in the high-osmolarity glycerol response pathway. Mol. Cell. Biol. 1998, 18, 5788–5796. [Google Scholar] [CrossRef] [Green Version]
- Barr, M.M.; Tu, H.; VanAelst, L.; Wigler, M. Identification of Ste4 as a potential regulator of Byr2 in the sexual response pathway of Schizosaccharomyces pombe. Mol. Cell. Biol. 1996, 16, 5597–5603. [Google Scholar] [CrossRef] [PubMed]
- Klosterman, S.J.; Martinez-Espinoza, A.D.; Andrews, D.L.; Seay, J.R.; Gold, S.E. Ubc2, an ortholog of the yeast Ste50p adaptor, possesses a basidiomycete-specific carboxy terminal extension essential for pathogenicity independent of pheromone response. Mol. Plant Microbe Interact. 2008, 21, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.A.; Bowie, J.U. SAM domains: Uniform structure, diversity of function. Trends Biochem. Sci. 2003, 28, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Aviv, T.; Lin, Z.; Ben-Ari, G.; Smibert, C.A.; Sicheri, F. Sequence-specific recognition of RNA hairpins by the SAM domain of Vts1p. Nat. Struct. Mol. Biol. 2006, 13, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.; Domadia, P.N.; Mohanram, H.; Bhattacharjya, S. NMR structural studies of the Ste11 SAM domain in the dodecyl phosphocholine micelle. Proteins 2009, 74, 328–343. [Google Scholar] [CrossRef]
- Slaughter, B.D.; Huff, J.M.; Wiegraebe, W.; Schwartz, J.W.; Li, R. SAM Domain-based protein oligomerization observed by live-cell fluorescence fluctuation spectroscopy. PLoS ONE 2008, 3, e1931. [Google Scholar] [CrossRef]
- Bhattacharjya, S.; Xu, P.; Gingras, R.; Shaykhutdinov, R.; Wu, C.L.; Whiteway, M.; Ni, F. Solution structure of the dimeric SAM domain of MAPKKK Ste11 and its interactions with the adaptor protein Ste50 from the budding yeast: Implications for Ste11 activation and signal transmission through the Ste50-Ste11 complex. J. Mol. Biol. 2004, 344, 1071–1087. [Google Scholar] [CrossRef]
- Kwan, J.J.; Warner, N.; Maini, J.; Tung, K.W.C.; Zakaria, H.; Pawson, T.; Donaldson, L.W. Saccharomyces cerevisiae Ste50 binds the MAPKKK Ste11 through a head-to-tail SAM domain interaction. J. Mol. Biol. 2006, 356, 142–154. [Google Scholar] [CrossRef]
- Xu, G.; Jansen, G.; Thomas, D.Y.; Hollenberg, C.P.; Rad, M.R. Ste50p sustains mating pheromone-induced signal transduction in the yeast Saccharomyces cerevisiae. Mol. Microbiol. 1996, 20, 773–783. [Google Scholar] [CrossRef]
- Truckses, D.M.; Bloomekatz, J.E.; Thorner, J. The RA domain of Ste50 adaptor protein is required for delivery of Ste11 to the plasma membrane in the filamentous growth signaling pathway of the yeast Saccharomyces cerevisiae. Mol. Cell. Biol. 2006, 26, 912–928. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.L.; Jansen, G.; Zhang, L.C.; Thomas, D.Y.; Whiteway, M. Adaptor protein Ste50p links the Ste11p MEKK to the HOG pathway through plasma membrane association. Gene. Dev. 2006, 20, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Ekiel, I.; Sulea, T.; Jansen, G.; Kowalik, M.; Minailiuc, O.; Cheng, J.; Harcus, D.; Cygler, M.; Whiteway, M.; Wu, C.L. Binding the atypical RA domain of Ste50p to the unfolded Opy2p cytoplasmic tail is essential for the high-osmolarity glycerol pathway. Mol. Biol. Cell 2009, 20, 5117–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus in U. virens | Putative Function | Ortholog in S. cerevisiae |
|---|---|---|
| UV8b_04866 | MAP kinase kinase EMK1 | Ste7 |
| UV8b_06470 | MAP kinase kinase kinase Ste11 | Ste11 |
| Uv8b_04241 | stress-activated MAP kinase | Hog1 |
| Uv8b_02467 | adenylate cyclase | Ac1 |
| Uv8b_00969 | putative adenylate cyclase-associated protein | Srv2 |
| UV8b_06597 | Peroxisomal biogenesis factor 6 | Pex6 |
| UV8b_04168 | GTPase activating protein (Gyp2) | Gyp2 |
| UV8b_04860 | cAMP-dependent protein kinase regulatory subunit | Bcy1 |
| UV8b_04229 | regulator of G protein signaling pathway | Sst2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, H.; Gong, H.; Song, T.; Yu, M.; Pan, X.; Yu, J.; Qi, Z.; Du, Y.; Liu, Y. The Adaptor Protein UvSte50 Governs Fungal Pathogenicity of Ustilaginoidea virens via the MAPK Signaling Pathway. J. Fungi 2022, 8, 954. https://doi.org/10.3390/jof8090954
Cao H, Gong H, Song T, Yu M, Pan X, Yu J, Qi Z, Du Y, Liu Y. The Adaptor Protein UvSte50 Governs Fungal Pathogenicity of Ustilaginoidea virens via the MAPK Signaling Pathway. Journal of Fungi. 2022; 8(9):954. https://doi.org/10.3390/jof8090954
Chicago/Turabian StyleCao, Huijuan, Hao Gong, Tianqiao Song, Mina Yu, Xiayan Pan, Junjie Yu, Zhongqiang Qi, Yan Du, and Yongfeng Liu. 2022. "The Adaptor Protein UvSte50 Governs Fungal Pathogenicity of Ustilaginoidea virens via the MAPK Signaling Pathway" Journal of Fungi 8, no. 9: 954. https://doi.org/10.3390/jof8090954