
Mitochondrial Porin Is Involved in Development, Virulence, and Autophagy in Fusarium graminearum
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Construction of Gene Deletion and Complementation Strains
2.3. Plant Infection and DON Production Assays
2.4. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Assays
2.5. Mitochondrial Membrane Potential, ATP, and H2O2 Analysis
2.6. Fluorescence and Transmission Electron Microscopy Assays
2.7. Autophagy Detection
2.8. Yeast Two-Hybrid and Bimolecular Fluorescence Complementation (BiFC) Assays
3. Results
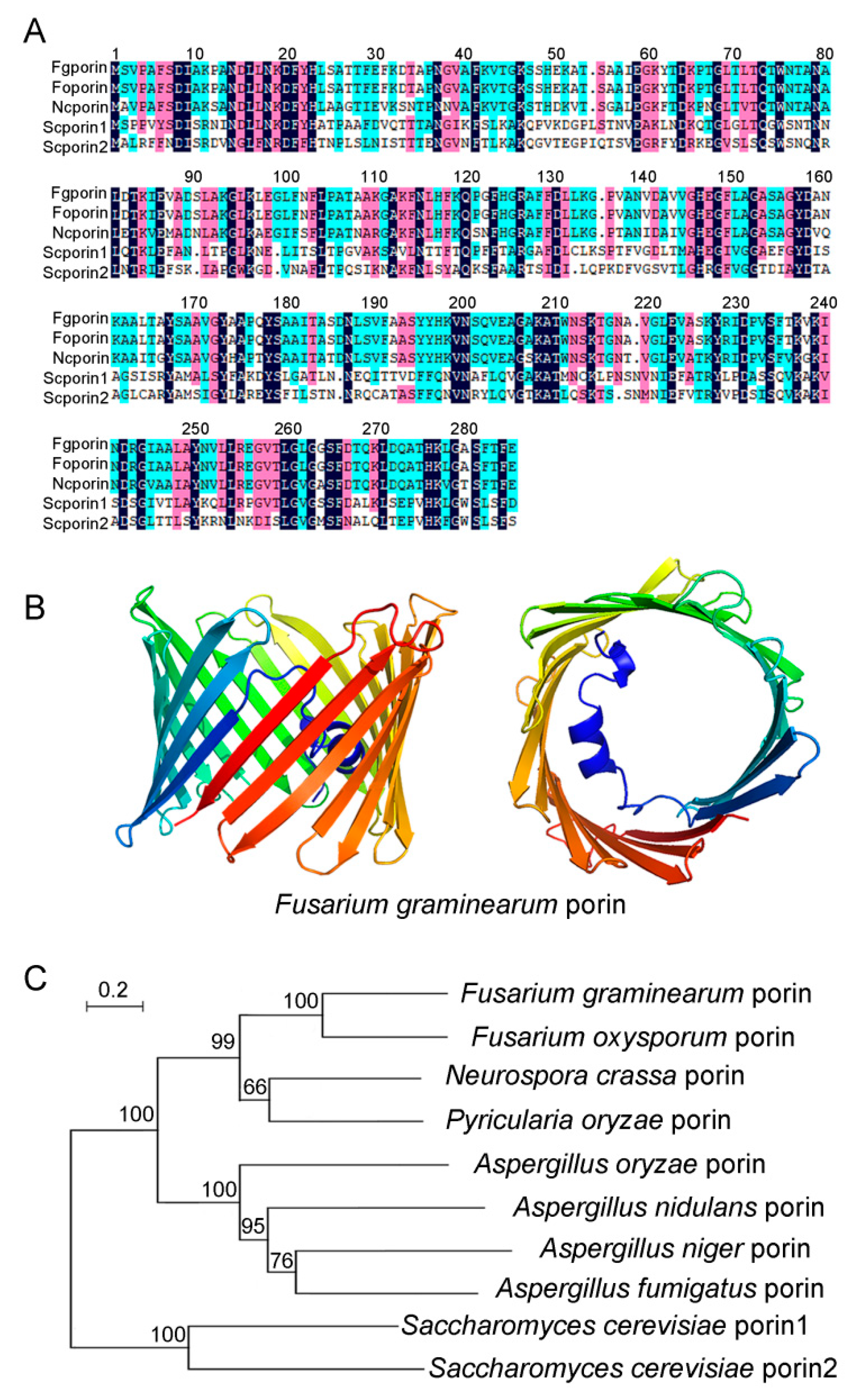
3.1. Identification of Fgporin in F. graminearum
3.2. Generation of Fgporin Deletion Mutants and Complementation Strains
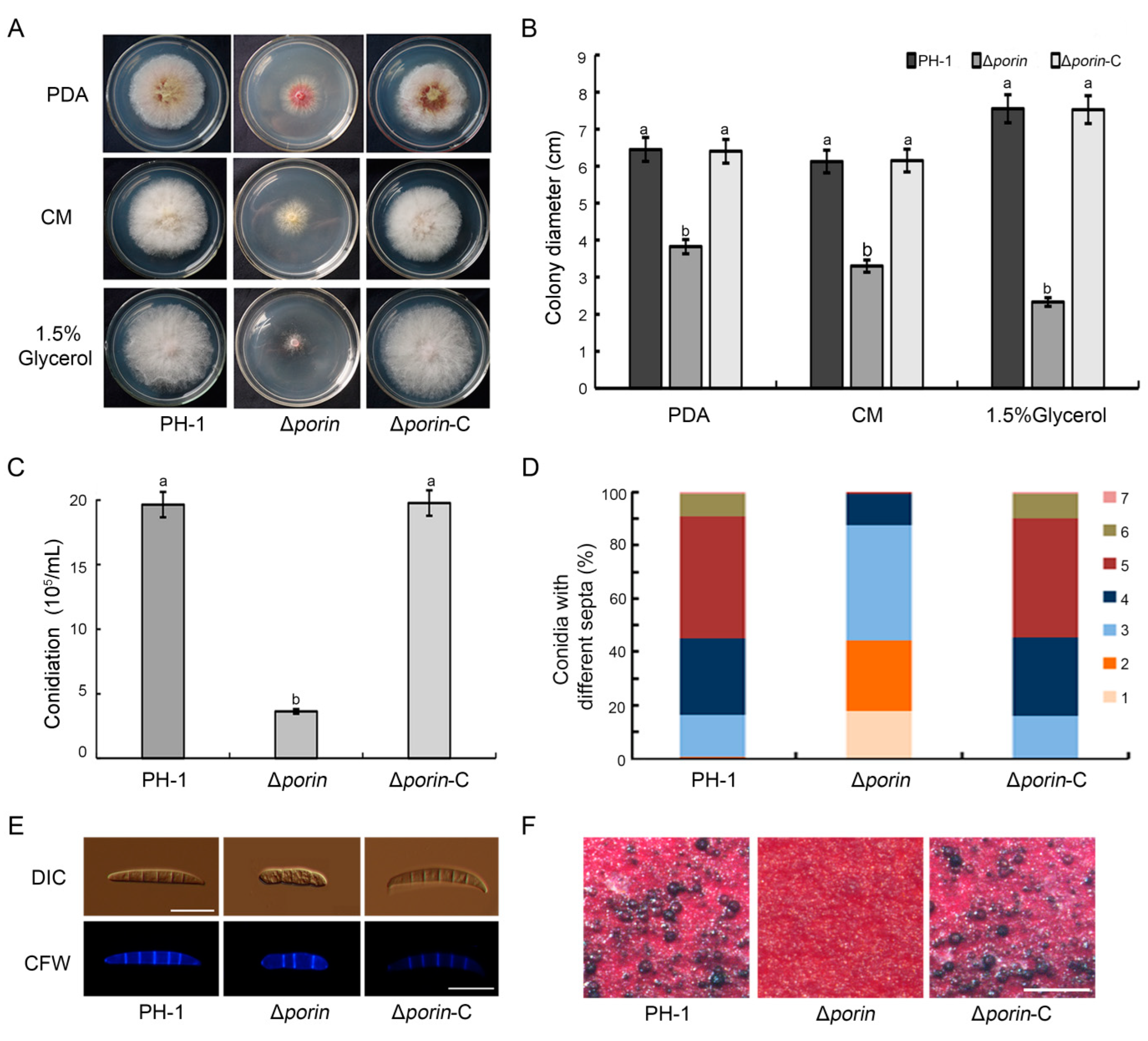
3.3. The Δporin Mutant Is Defective in Mycelial Growth, Conidiation, and Sexual Reproduction
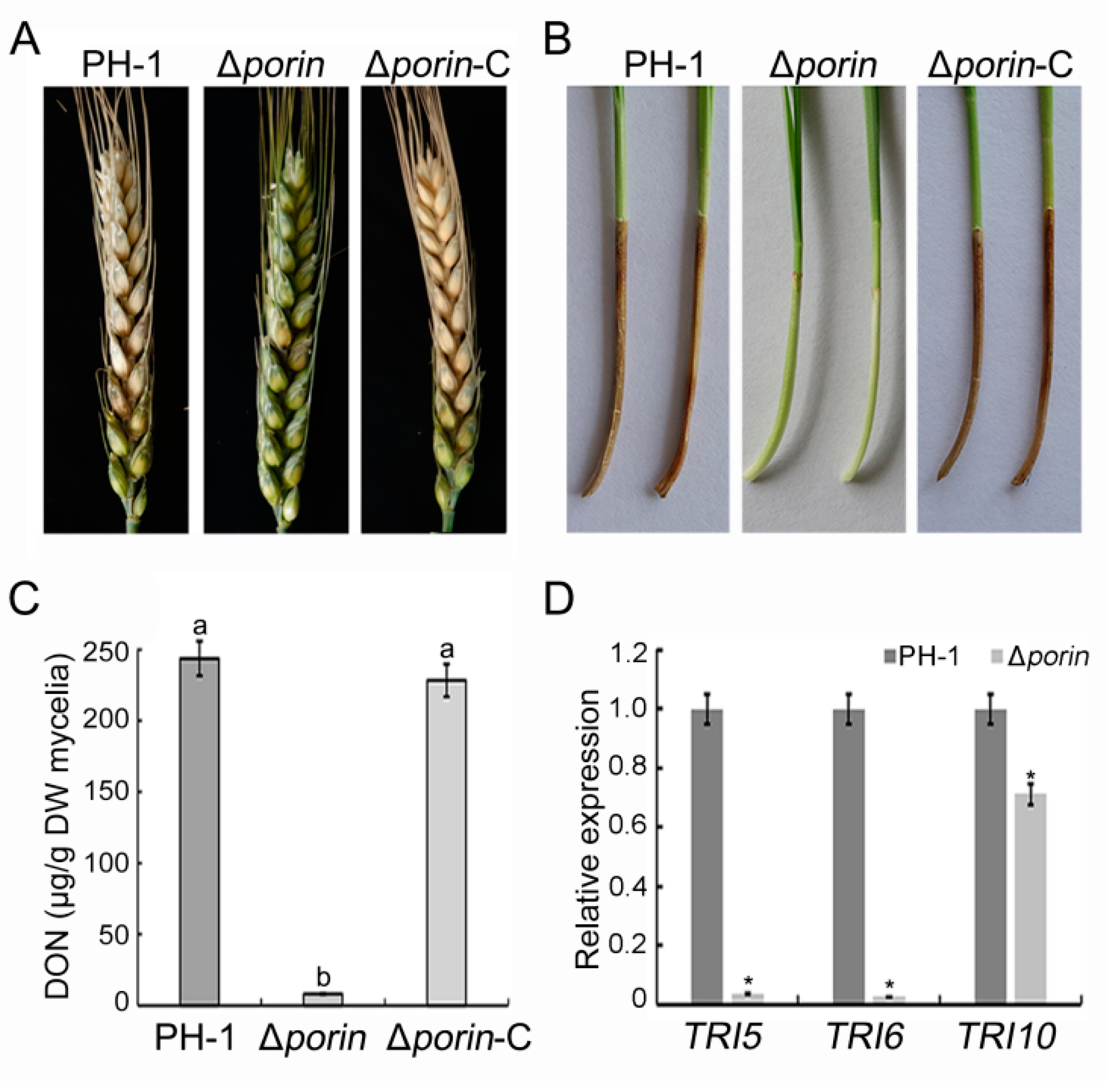
3.4. Δporin Strain Reduces Virulence and DON Production
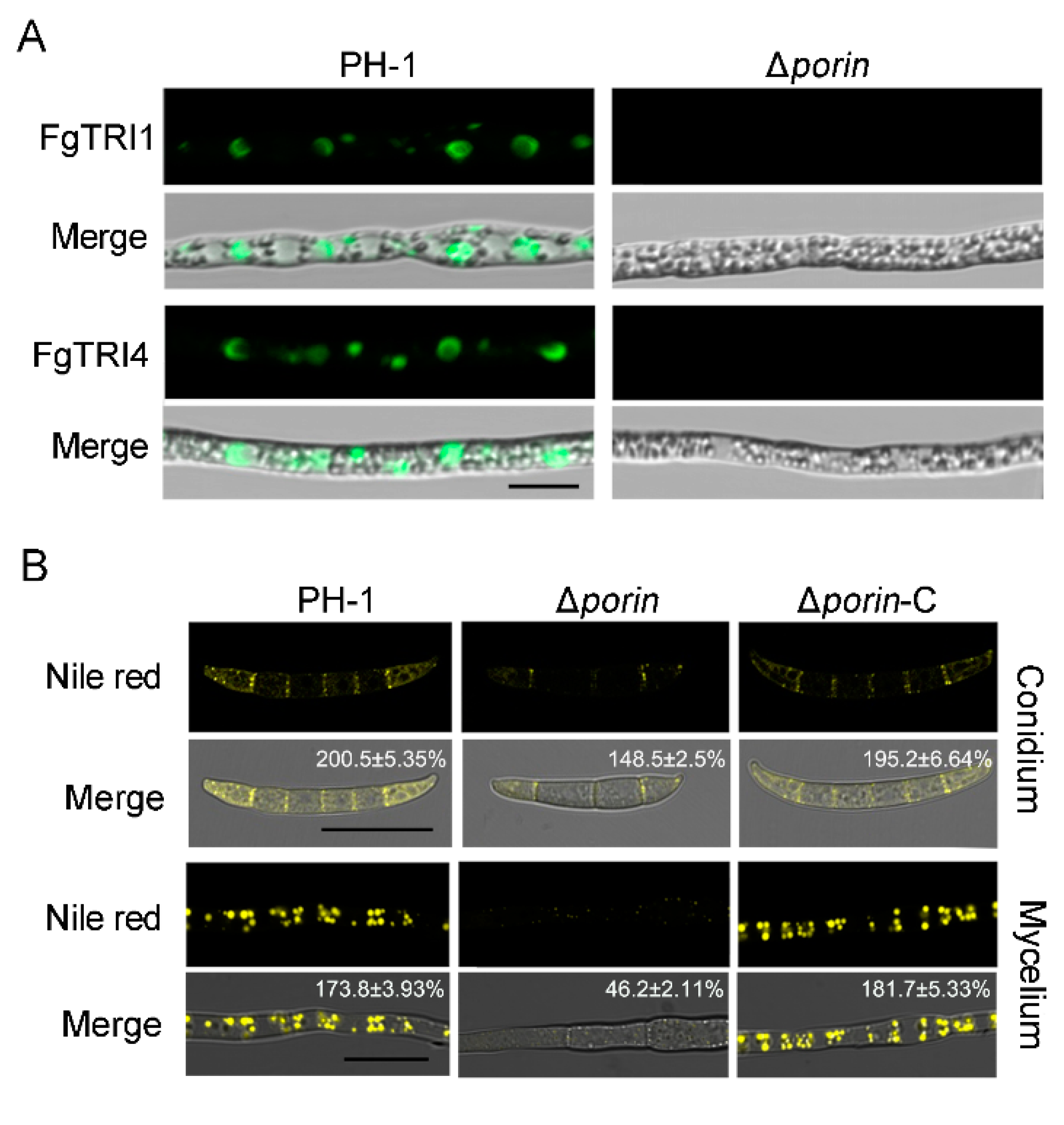
3.5. Fgporin Is Involved in the Formation of Toxisomes and Lipid Droplets
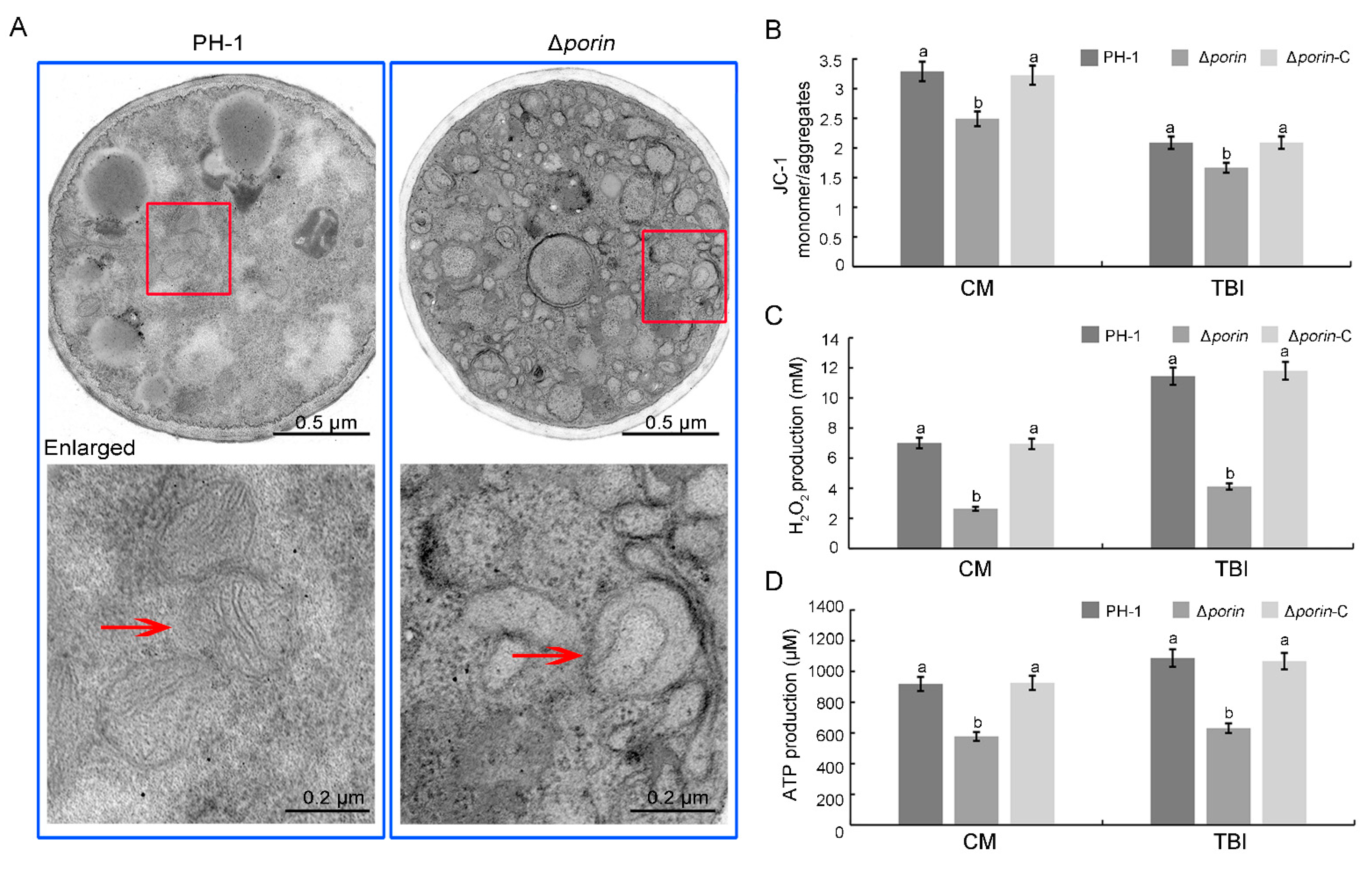
3.6. The Δporin Strain Results in Defects in Mitochondrial Morphology and Function
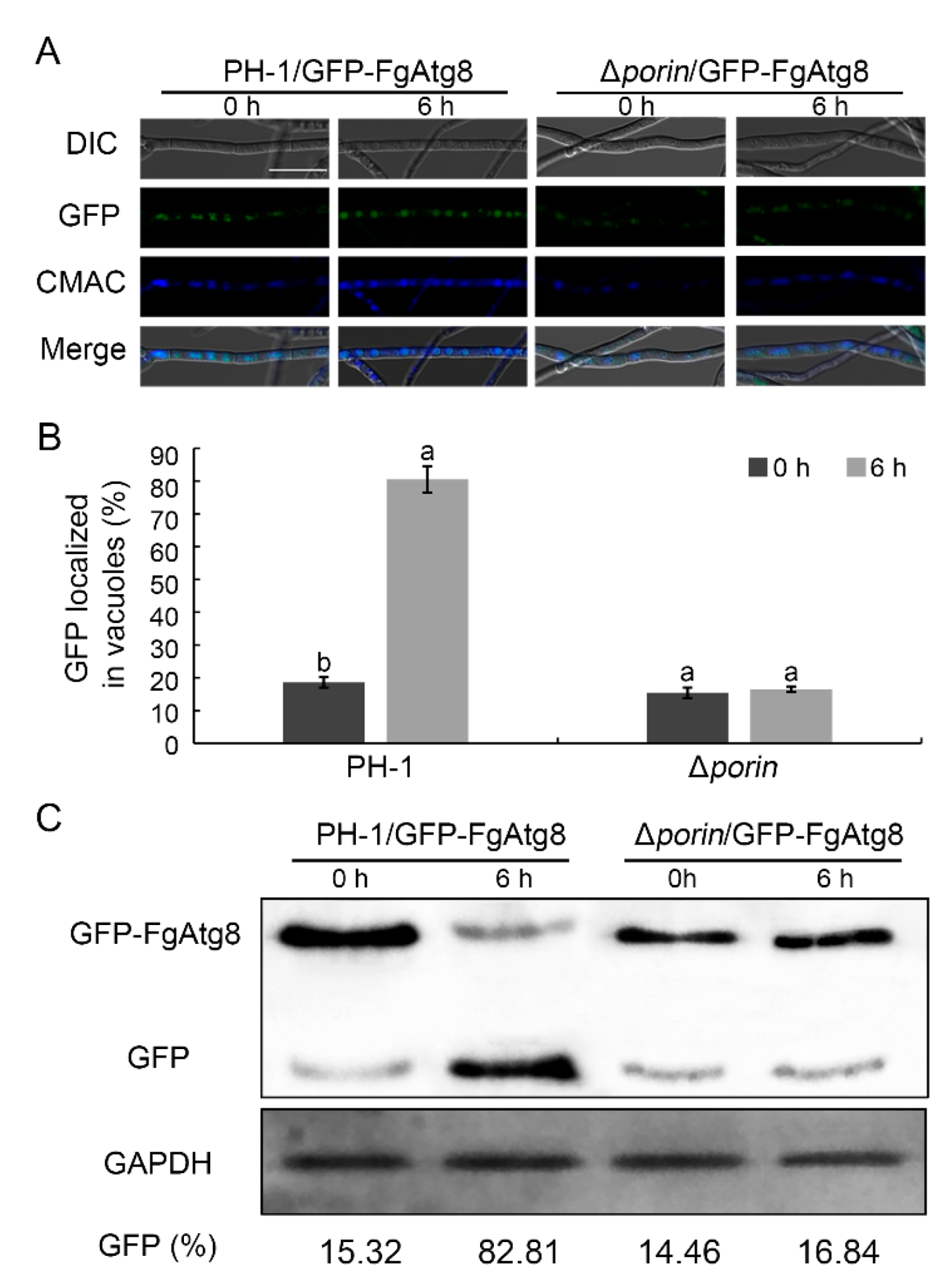
3.7. The Fgporin Deletion Mutant Is Defective in Autophagy
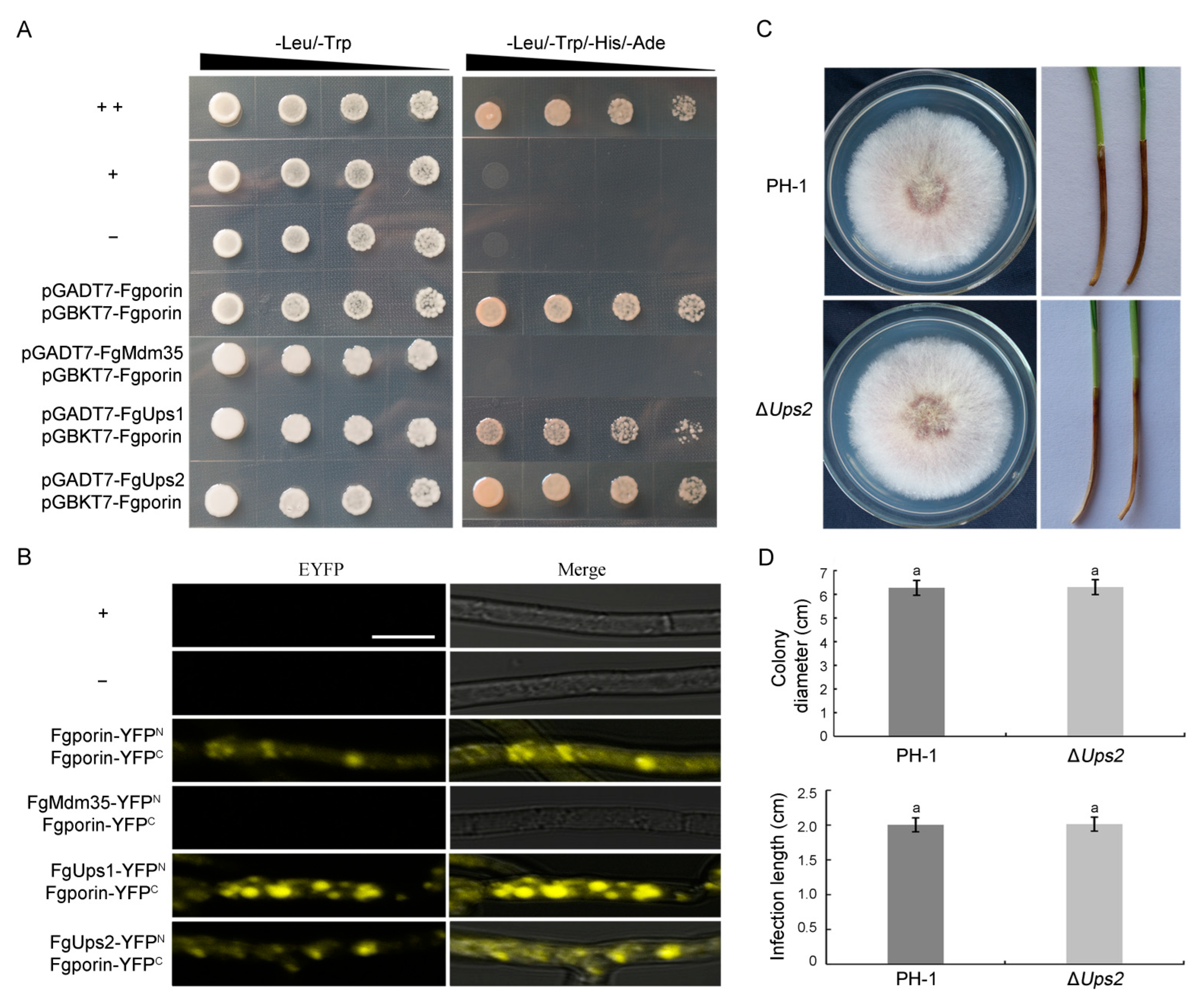
3.8. Fgporin Interacts with Itself and FgUps1/2, but Not with FgMdm35
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devasting impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in fusarium species: Chemistry, genetics, and significance. Microbiol. Rev. 1993, 57, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant-Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum trichothecene mycotoxins: Biosynthesis, regulation, and management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef]
- Schmitt, S.; Prokisch, H.; Schlunck, T.; Camp Ii, D.G.; Ahting, U.; Waizenegger, T.; Scharfe, C.; Meitinger, T.; Imhof, A.; Neupert, W.; et al. Proteome analysis of mitochondrial outer membrane from Neurospora crassa. Proteomics 2006, 6, 72–80. [Google Scholar] [CrossRef]
- Hiller, S.; Abramson, J.; Mannella, C.; Wagner, G.; Zeth, K. The 3D structures of VDAC represent a native conformation. Trends Biochem. Sci. 2010, 35, 514–521. [Google Scholar] [CrossRef]
- Messina, A.; Reina, S.; Guarino, F.; De Pinto, V. VDAC isoforms in mammals. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1466–1476. [Google Scholar] [CrossRef]
- Morgenstern, M.; Stiller, S.B.; Lübbert, P.; Peikert, C.D.; Dannenmaier, S.; Drepper, F.; Weill, U.; Höß, P.; Feuerstein, R.; Gebert, M.; et al. Definition of a high-confidence mitochondrial proteome at quantitative scale. Cell Rep. 2017, 19, 2836–2852. [Google Scholar] [CrossRef]
- Dihanich, M.; Suda, K.; Schatz, G. A yeast mutant lacking mitochondrial porin is respiratory-deficient, but can recover respiration with simultaneous accumulation of an 86-kd extramitochondrial protein. EMBO J. 1987, 6, 723–728. [Google Scholar] [CrossRef]
- Lee, A.C.; Xu, X.; Blachly-Dyson, E.; Forte, M.; Colombini, M. The role of yeast VDAC genes on the permeability of the mitochondrial outer membrane. J. Membr. Biol. 1998, 161, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.H.; Craigen, W.J. Genetic approaches to analyzing mitochondrial outer membrane permeability. Curr. Top. Dev. Biol. 2004, 59, 87–118. [Google Scholar] [CrossRef] [PubMed]
- Broeskamp, F.; Edrich, E.S.M.; Knittelfelder, O.; Neuhaus, L.; Meyer, T.; Heyden, J.; Habernig, L.; Kreppel, F.; Gourlay, C.W.; Rockenfeller, P. Porin 1 modulates autophagy in yeast. Cells 2021, 10, 2416. [Google Scholar] [CrossRef]
- Tamura, Y.; Iijima, M.; Sesaki, H. Mdm35p imports Ups proteins into the mitochondrial intermembrane space by functional complex formation. EMBO J. 2010, 29, 2875–2887. [Google Scholar] [CrossRef] [PubMed]
- Miyata, N.; Fujii, S.; Kuge, O. porin proteins have critical functions in mitochondrial phospholipid metabolism in yeast. J. Biol. Chem. 2018, 293, 17593–17605. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Sakaue, H. Multifaceted roles of porin in mitochondrial protein and lipid transport. Biochem. Soc. Trans. 2019, 47, 1269–1277. [Google Scholar] [CrossRef]
- Summers, W.A.T.; Wilkins, J.A.; Dwivedi, R.C.; Ezzati, P.; Court, D.A. Mitochondrial dysfunction resulting from the absence of mitochondrial porin in Neurospora crassa. Mitochondrion 2012, 12, 220–229. [Google Scholar] [CrossRef]
- Craigen, W.J.; Graham, B.H. Genetic strategies for dissecting mammalian and drosophila voltage-dependent anion channel functions. J. Bioenerg. Biomembr. 2008, 40, 207–212. [Google Scholar] [CrossRef]
- Kinnally, K.W.; Peixoto, P.M.; Ryu, S.-Y.; Dejean, L.M. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochim. Biophys. Acta Molecul. 2011, 1813, 616–622. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Sheldon, K.L.; DeHart, D.N.; Patnaik, J.; Manevich, Y.; Townsend, D.M.; Bezrukov, S.M.; Rostovtseva, T.K.; Lemasters, J.J. Voltage-dependent anion channels modulate mitochondrial metabolism in cancer cells: Regulation by free tubulin and erastin. J. Biol. Chem. 2013, 288, 11920–11929. [Google Scholar] [CrossRef] [Green Version]
- Bosc, C.; Broin, N.; Fanjul, M.; Saland, E.; Farge, T.; Courdy, C.; Batut, A.; Masoud, R.; Larrue, C.; Skuli, S.; et al. Autophagy regulates fatty acid availability for oxidative phosphorylation through mitochondria-endoplasmic reticulum contact sites. Nat. Commun. 2020, 11, 4056. [Google Scholar] [CrossRef]
- Hou, Z.; Xue, C.; Peng, Y.; Katan, T.; Kistler, H.C.; Xu, J.-R. A mitogen-activated protein kinase gene (Mgv1) in Fusarium graminearum is required for female fertility, heterokaryon formation, and plant infection. Mol. Plant-Microbe Interact. 2002, 15, 1119–1127. [Google Scholar] [CrossRef]
- Cavinder, B.; Sikhakolli, U.; Fellows, K.M.; Trail, F. Sexual development and ascospore discharge in Fusarium graminearum. J. Vis. Exp. 2012, 61, 3895. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Kazan, K.; Manners, J.M. Nutrient profilin reveals potent inducers of trichothecene biosynthesis in Fusarium graminearum. Fungal Genet. Biol. 2009, 46, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Catlett, N.L.; Lee, B.; Yoder, O.; Turgeon, B. Split-marker recombination for efficient targeted deletion of fungal genes. Fung. Genet. Rep. 2003, 50, 9–11. [Google Scholar] [CrossRef]
- Liu, Z.; Friesen, T.L. Polyethylene glycol (PEG)-mediated transformation in filamentous fungal pathogens. Plant Fung. Pathog. 2011, 835, 365–375. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ohneda, M.; Arioka, M.; Nakajima, H.; Kitamoto, K. Visualization of vacuoles in aspergillus oryzae by expression of CPY–EGFP. Fungal Genet. Biol. 2002, 37, 29–38. [Google Scholar] [CrossRef]
- Shi, H.-B.; Chen, N.; Zhu, X.-M.; Su, Z.-Z.; Wang, J.-Y.; Lu, J.-P.; Liu, X.-H.; Lin, F.-C. The casein kinase moyck1 regulates development, autophagy, and virulence in the rice blast fungus. Virulence 2019, 10, 719–733. [Google Scholar] [CrossRef] [Green Version]
- Schiestl, R.H.; Gietz, R.D. High efficiency transformation of intact yeast cells using single stranded nucleic acids as a carrier. Curr. Genet. 1989, 16, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, W.; Wang, C.; Xu, Q.; Wang, Y.; Ding, S.; Xu, J.-R. A MADS-box transcription factor MoMcm1 is required for male fertility, microconidium production and virulence in Magnaporthe oryzae. Mol. Microbiol. 2011, 80, 33–53. [Google Scholar] [CrossRef]
- Sampson, M.J.; Lovell, R.S.; Craigen, W.J. The murine voltage-dependent anion channel gene family: Conserved structure and function. J. Biol. Chem. 1997, 272, 18966–18973. [Google Scholar] [CrossRef] [PubMed]
- Shuvo, S.R.; Kovaltchouk, U.; Zubaer, A.; Kumar, A.; Summers, W.A.; Donald, L.J.; Hausner, G.; Court, D.A. Functional characterization of an N-terminally truncated mitochondrial porin expressed in Neurospora crassa. Can. J. Microbiol. 2017, 63, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Menke, J.; Weber, J.; Broz, K.; Kistler, H.C. Cellular development associated with induced mycotoxin synthesis in the filamentous fungus Fusarium graminearum. PLoS ONE 2013, 8, e63077. [Google Scholar] [CrossRef]
- Chong, X.; Wang, C.; Wang, Y.; Wang, Y.; Zhang, L.; Liang, Y.; Chen, L.; Zou, S.; Dong, H. The dynamin-like GTPase FgSey1 plays a critical role in fungal development and virulence in Fusarium graminearum. Appl. Environ. Microbiol. 2020, 86, e02720-19. [Google Scholar] [CrossRef]
- Liu, N.; Yun, Y.; Yin, Y.; Hahn, M.; Ma, Z.; Chen, Y. Lipid droplet biogenesis regulated by the FgNem1/Spo7-FgPah1 phosphatase cascade plays critical roles in fungal development and virulence in Fusarium graminearum. New Phytol. 2019, 223, 412–429. [Google Scholar] [CrossRef]
- Magrì, A.; Di Rosa, M.C.; Orlandi, I.; Guarino, F.; Reina, S.; Guarnaccia, M.; Morello, G.; Spampinato, A.; Cavallaro, S.; Messina, A.; et al. Deletion of voltage-dependent anion channel 1 knocks mitochondria down triggering metabolic rewiring in yeast. Cell. Mol. Life Sci. 2020, 77, 3195–3213. [Google Scholar] [CrossRef]
- Mallo, N.; Ovciarikova, J.; Martins Duarte, E.; Baehr, S.; Biddau, M.; Wilde, M.-L.; Uboldi, A.D.; Lemgruber, L.; Tonkin, C.J.; Wideman, J.G.; et al. Depletion of a Toxoplasma porin leads to defects in mitochondrial morphology and contacts with the ER. J. Cell Sci. 2021, 134. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.; Choi, S.; Koh, H.; Lee, S.-H.; Kim, J.-M.; Chung, J. Drosophila porin/VDAC affects mitochondrial morphology. PLoS ONE 2010, 5, e13151. [Google Scholar] [CrossRef] [Green Version]
- Handy, D.E.; Loscalzo, J. Redox regulation of mitochondrial function. Antioxid. Redox Signal. 2012, 16, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- Danley, D.L.; Hilger, A.E.; Winkel, C.A. Generation of hydrogen peroxide by candida albicans and influence on murine polymorphonuclear leukocyte activity. Infect Immun. 1983, 40, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Staniek, K.; Gille, L.; Kozlov, A.V.; Nohl, H. Mitochondrial superoxide radical formation is controlled by electron bifurcation to the high and low potential pathways. Free Radic. Res. 2002, 36, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Magrì, A.; Di Rosa, M.C.; Tomasello, M.F.; Guarino, F.; Reina, S.; Messina, A.; De Pinto, V. Overexpression of human SOD1 in VDAC1-less yeast restores mitochondrial functionality modulating beta-barrel outer membrane protein genes. Biochim. Biophys. Acta 2016, 1857, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Josefsen, L.; Droce, A.; Sondergaard, T.E.; Sørensen, J.L.; Bormann, J.; Schäfer, W.; Giese, H.; Olsson, S. Autophagy provides nutrients for nonassimilating fungal structures and is necessary for plant colonization but not for infection in the necrotrophic plant pathogen Fusarium graminearum. Autophagy 2012, 8, 326–337. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Conidial Length (μm) a | Disease Index on Wheat Heads b | Lesion Length on Coleoptiles (cm) c |
|---|---|---|---|
| PH-1 | 45.41 ± 6.52 a | 12.4 ± 1.5 a | 1.82 ± 0.18 a |
| ∆porin | 35.73 ± 5.12 b | 0.7 ± 0.3 b | 0.12 ± 0.04 b |
| ∆porin-C | 45.32 ± 4.32 a | 12.6 ± 1.8 a | 1.88 ± 0.13 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Li, Q.; Li, X.; Lv, X.; Zhang, L.; Zou, S.; Yu, J.; Dong, H.; Chen, L.; Liang, Y. Mitochondrial Porin Is Involved in Development, Virulence, and Autophagy in Fusarium graminearum. J. Fungi 2022, 8, 936. https://doi.org/10.3390/jof8090936
Han X, Li Q, Li X, Lv X, Zhang L, Zou S, Yu J, Dong H, Chen L, Liang Y. Mitochondrial Porin Is Involved in Development, Virulence, and Autophagy in Fusarium graminearum. Journal of Fungi. 2022; 8(9):936. https://doi.org/10.3390/jof8090936
Chicago/Turabian StyleHan, Xueqin, Qingyi Li, Xuenan Li, Xiang Lv, Li Zhang, Shenshen Zou, Jinfeng Yu, Hansong Dong, Lei Chen, and Yuancun Liang. 2022. "Mitochondrial Porin Is Involved in Development, Virulence, and Autophagy in Fusarium graminearum" Journal of Fungi 8, no. 9: 936. https://doi.org/10.3390/jof8090936