
Antifungal Activity of Copper Oxide Nanoparticles against Root Rot Disease in Cucumber
 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of Cu2ONPs
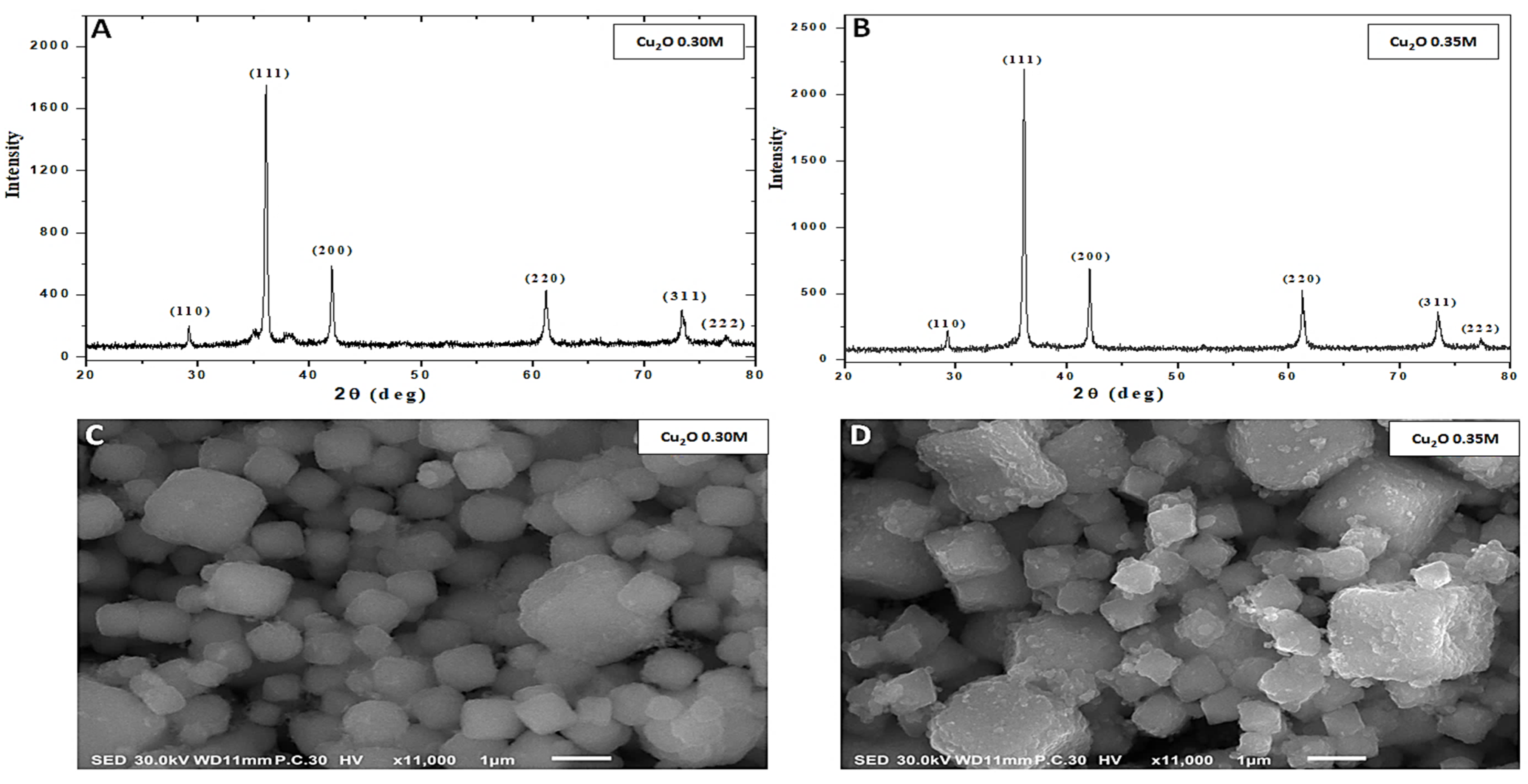
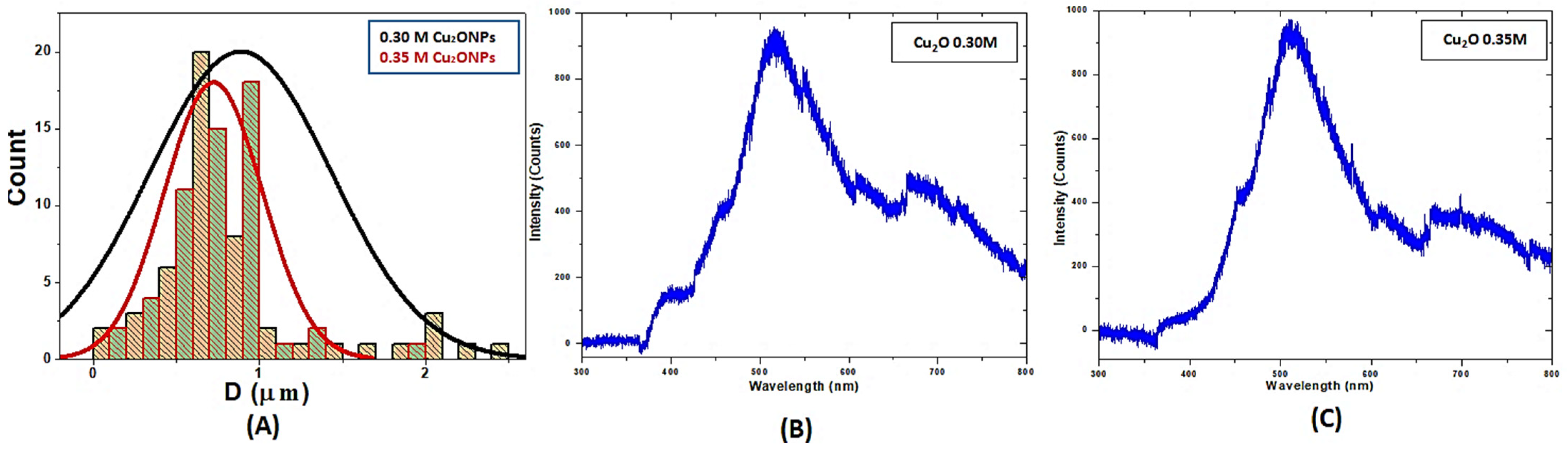
2.2. Characterization of Cu2ONPs
2.3. Plant Materials
2.4. Fungal Pathogen Identification
2.5. Pathogenicity Test
2.6. Effects of Cu2ONP Concentrations on Fungal Growth
2.7. In Vivo (Greenhouse) Experiment
2.8. Laboratory Studies
2.8.1. Enzyme Activity Assay
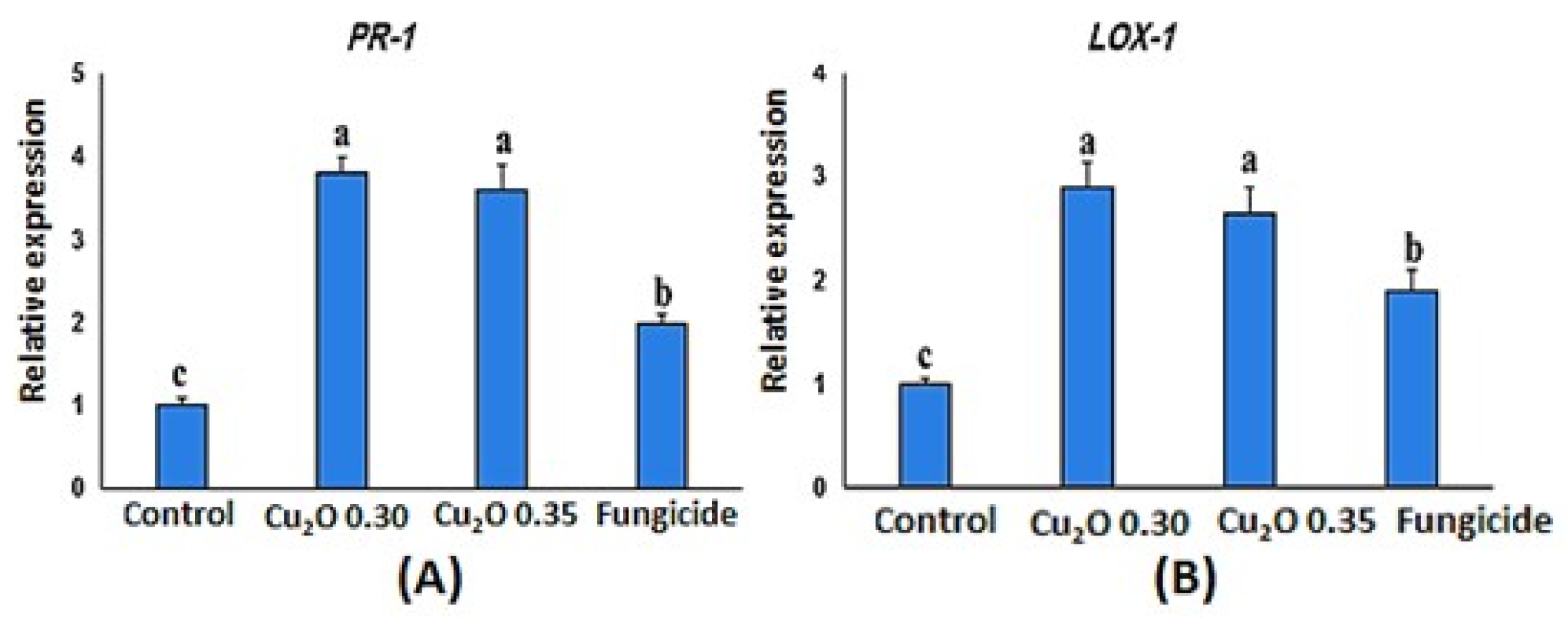
2.8.2. RT-PCR Analysis
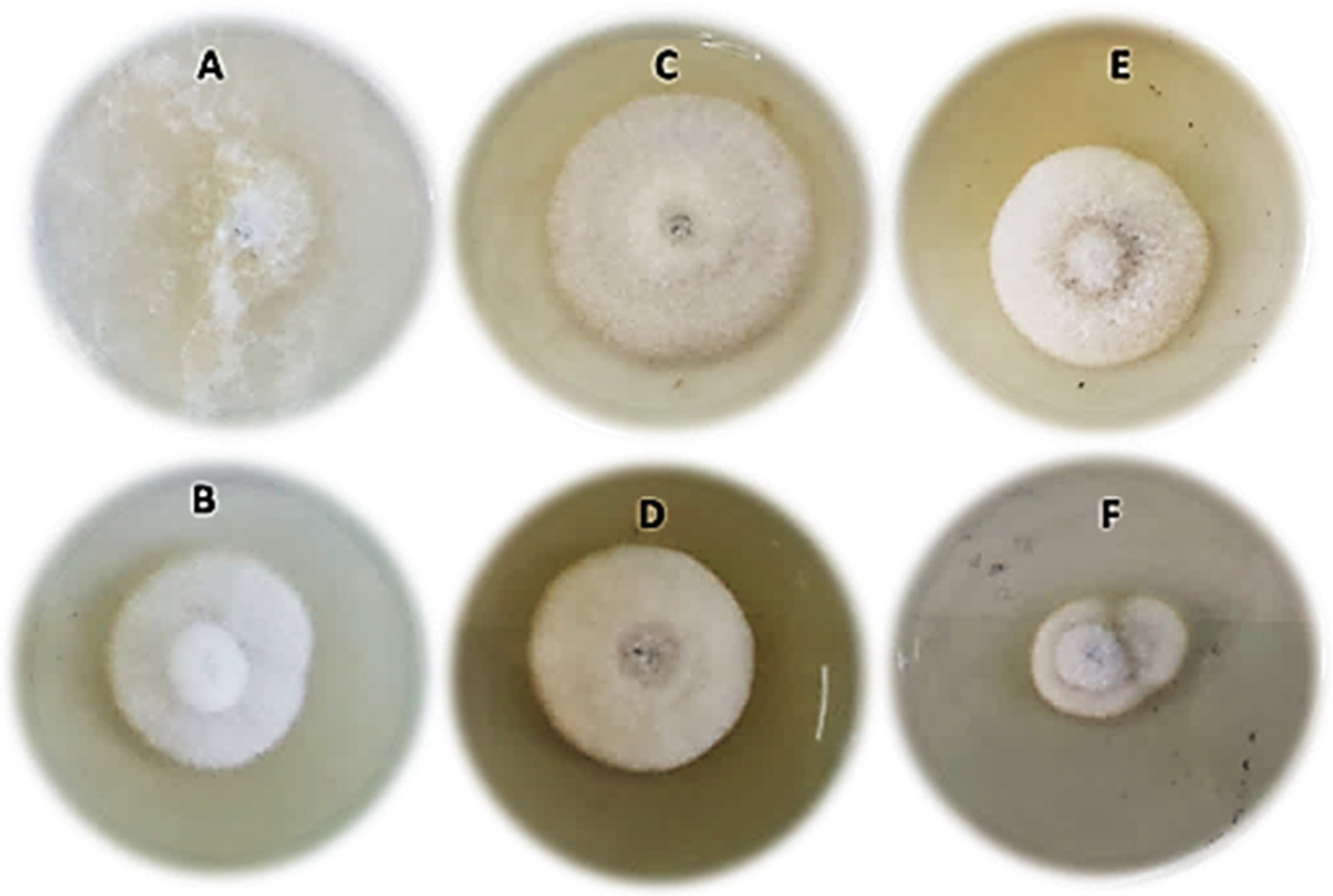
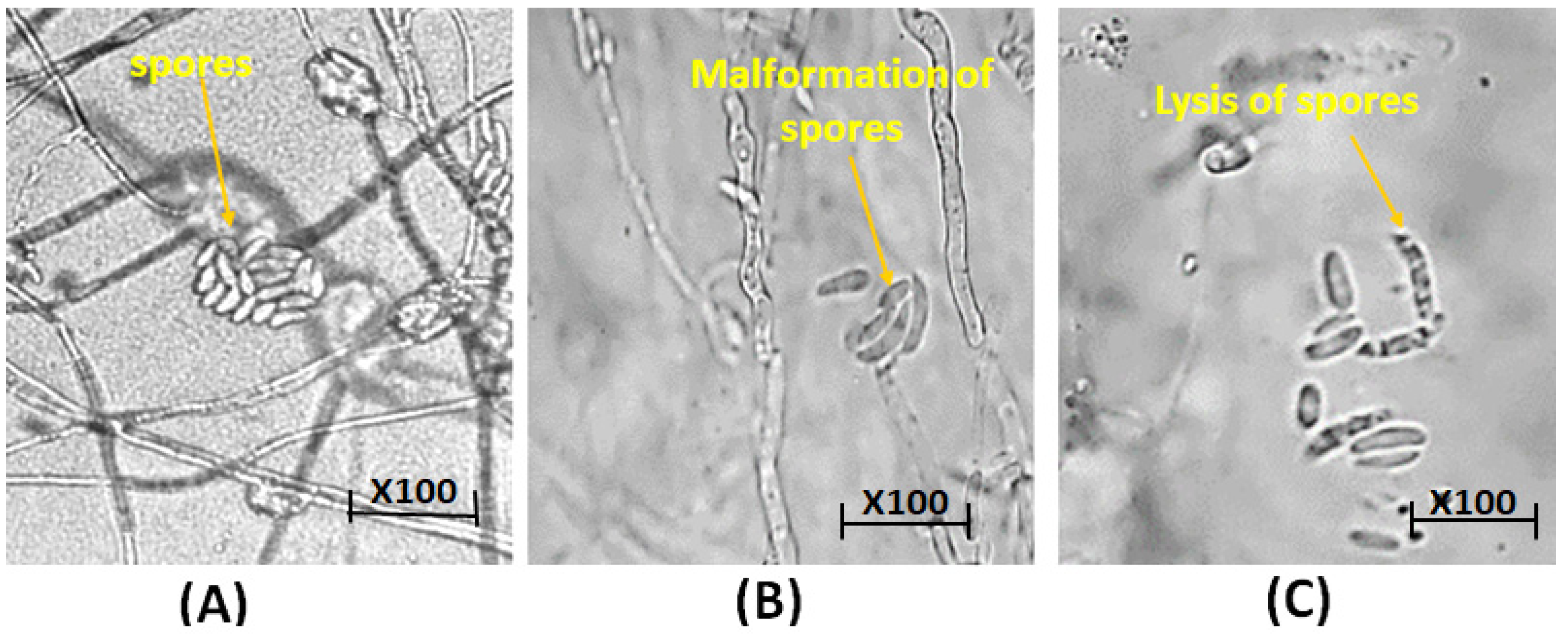
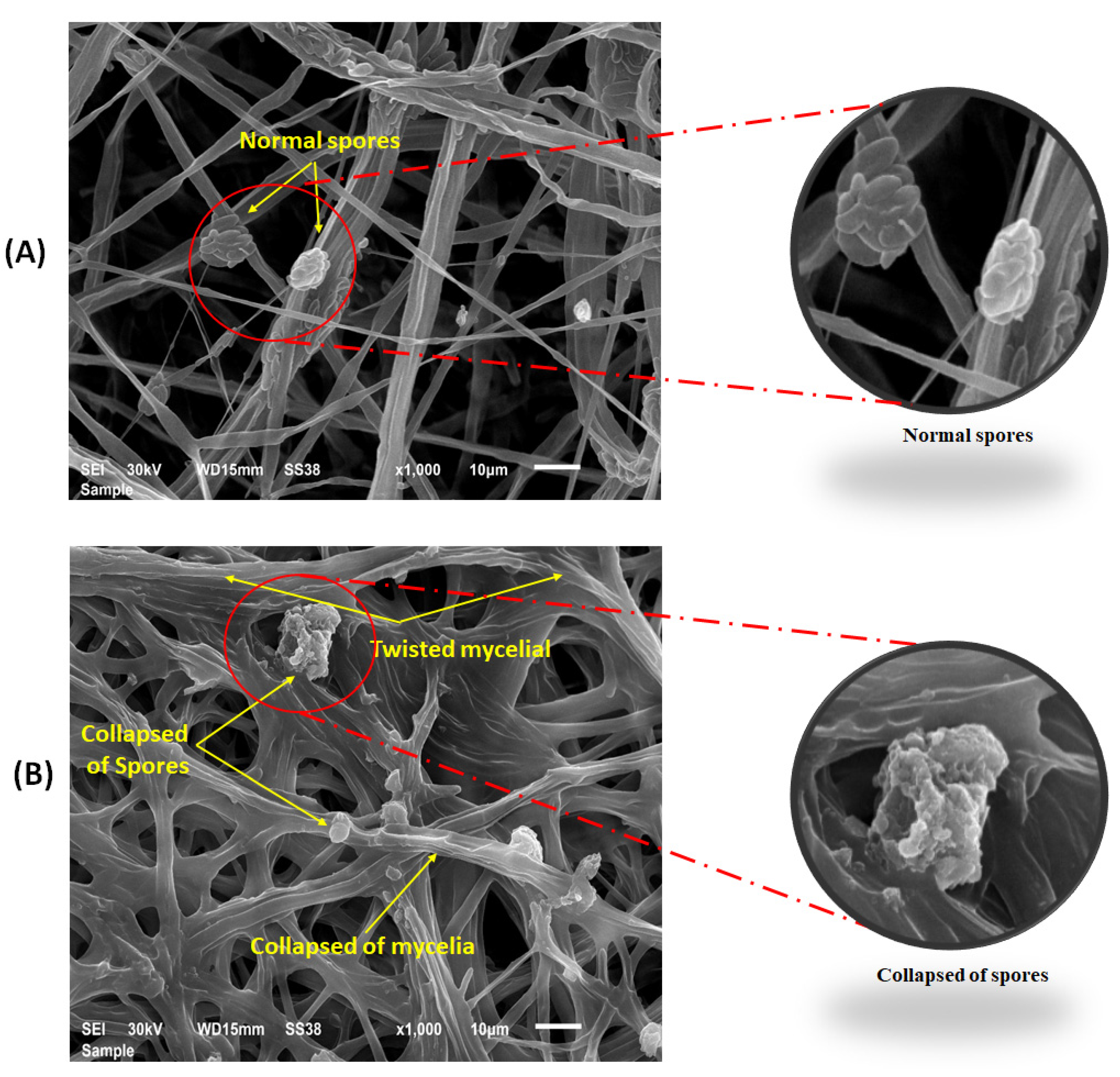
2.8.3. Microscopic Observations of Fungal Morphology
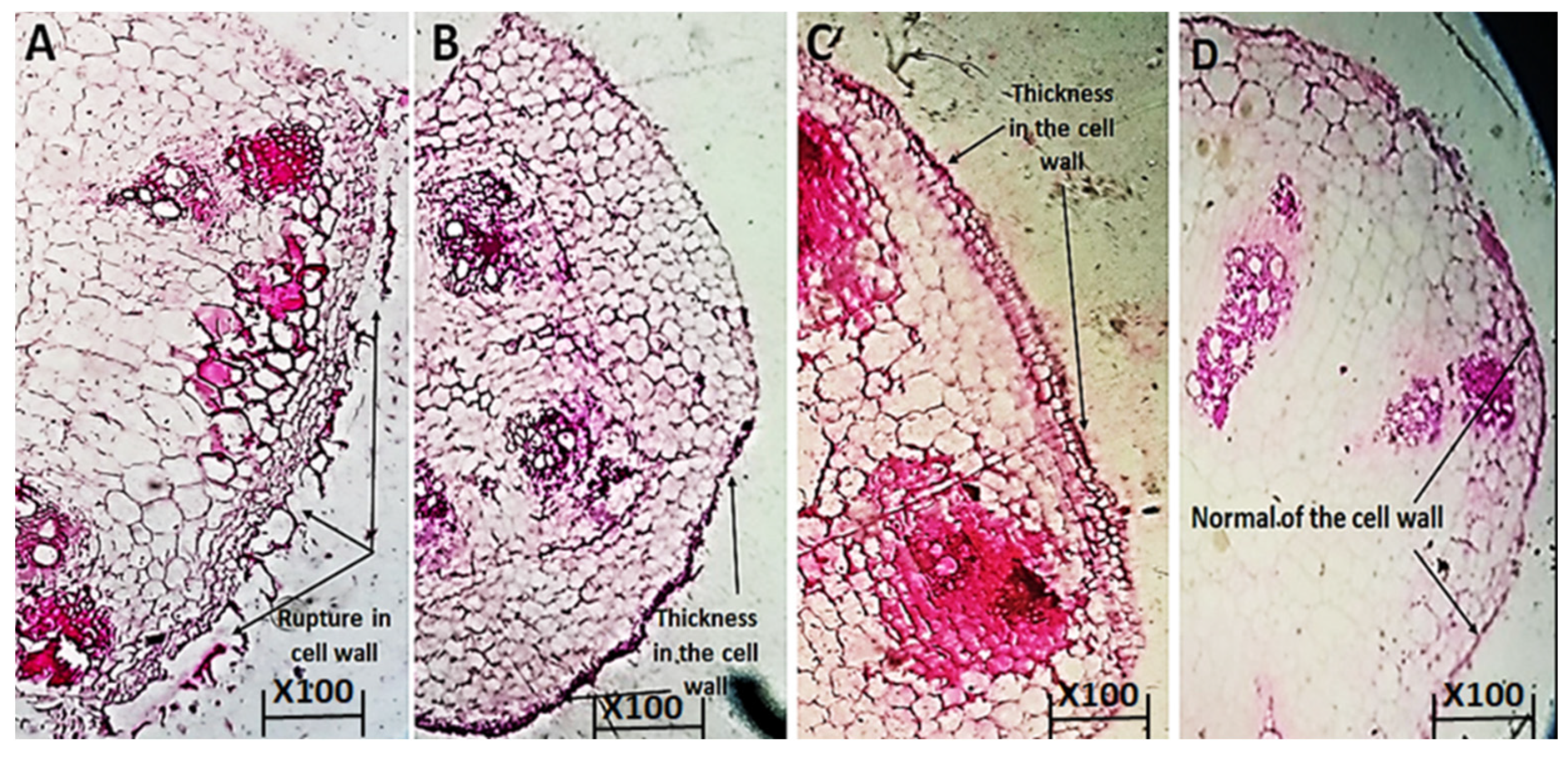
2.8.4. The Anatomical Structure of Cucumber Plants
2.9. Field Experiment
2.10. Growth and Yield Parameters
2.11. Statistical Analysis
3. Results
3.1. Characterization of Cu2ONPs
3.2. Antifungal Activity of Cu2ONPs against F. solani under Laboratory Conditions
3.3. Effect of Cu2ONPs on Disease Incidence under Greenhouse Conditions
3.4. Effect of Cu2ONPs on the Activity of Defense Enzymes in Treated Cucumber Plants
3.5. Relative Expression Assay
3.6. Laboratory Studies
3.6.1. Microscopic Observations of Fungal Morphology
3.6.2. The Anatomical Structure of Cucumber Plants
3.7. Effect of Cu2ONPs on Disease Incidence in Two Locations under Field Conditions
3.8. Effect of Cu2ONPs on Total Chlorophyll and Growth Parameter of Cucumber Plants under Field Conditions
3.9. Effect of Cu2ONPs on Yield Parameter of Cucumber Plants under Field Conditions
3.10. Pearson Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Retrieved 2020-02-15. Countries-Select All; Regions-World + (Total); Elements-Production Quantity; Items-Cucumbers and Gherkins; FAOSTAT: Rome, Italy, 2018. [Google Scholar]
- Szalay, J. Cucumbers: Health Benefits and Nutrition Facts. 2017. Available online: https://www.livescience.com/51000-cucumber-nutrition.html (accessed on 22 January 2022).
- Abd-El-Moneium, M.L. Integrated system to protect cucumber plants in greenhouses against diseases and pests under organic farming conditions. Egypt. J. Agric. Res. 2004, 82, 1–9. [Google Scholar]
- Roberts, D.P.; Lohrke, S.M.; Meyer, S.L.; Chung, S. Biological agents applied individually or in combination for suppression of soil borne diseases of cucumber plants. Crop. Prot. 2005, 24, 135–141. [Google Scholar] [CrossRef]
- Jinghua, Z.; Chang, W.; Xu, W.; Hanlian, W.; Shuge, T. Allelopathy of diseased survival on cucumber Fusarium wilt. Acta Phytophylacica Sin. 2008, 35, 317–321. [Google Scholar]
- Elagamey, E.; Abdellatef, M.A.E.; Kamel, S.M.; Essa, T.A. Fusarium oxysporum isolates collected from the same geographical zone exhibited variations in disease severity and diversity in morphological and molecular characters. Egypt. J. Phytopathol. 2020, 48, 43–57. [Google Scholar] [CrossRef]
- Cuervo-Parra, J.A.; Sánchez-López, V.; Ramírez-Suero, M.; Ramírez-Lepe, M. Morphological and molecular characterization of Moniliophthora roreri causal agent of frosty pod rot of cocoa tree in Tabasco, México. Plant. Pathol. J. 2011, 10, 122–127. [Google Scholar] [CrossRef]
- Omara, R.I.; Kamel, S.M.; Hafez, Y.M.; Morsy, S.Z. Role of Non-traditional control treatments in inducing resistance against wheat leaf rust caused by Puccinia Triticina. Egypt. J. Biol. Pest. Control. 2015, 25, 335–344. [Google Scholar]
- Omara, R.I.; Essa, T.A.; Khalil, A.A.; Elsharkawy, M.M. A case study of non-traditional treatments for the control of wheat stem rust disease. Egypt. J. Biol. Pest. Control. 2020, 30, 1–12. [Google Scholar] [CrossRef]
- Krauss, U.; Hoopen, M.; Rees, R.; Stirrup, T.; Argyle, T.; George, A.; Arroyo, C.; Corrales, E.; Casanoves, F. Mycoparasitism by Clonostachys byssicola and Clonostachys rosea on Trichoderma spp. from cocoa (Theobroma cacao) and implication for the design of mixed biocontrol agents. Biol. Control. 2013, 67, 317–327. [Google Scholar] [CrossRef]
- Mbarga, J.B.; Begoude, B.A.d.; Ambang, Z.; Meboma, M.; Kuate, J.; Schiffers, B.; Wwbank, W.; Dedieu, I.; Hoopen, G.M. A new oil-based formulation of Trichoderma asperellum for the biological control of cacao black pod disease caused by Phytophthora megakarya. Biol. Control. 2014, 77, 15–22. [Google Scholar] [CrossRef]
- Kazemi, M.; Akbari, A.; Sabouri, Z.; Soleimanpour, S.; Zarrinfar, H.; Khatami, M.; Darroudi, M. Green synthesis of colloidal selenium nanoparticles in starch solutions and investigation of their photocatalytic, antimicrobial, and cytotoxicity effects. Bioprocess Biosyst. Eng. 2021, 44, 1215–1225. [Google Scholar] [CrossRef]
- Kazemi, M.; Akbari, A.; Zarrinfar, H.; Soleimanpour, S.; Sabouri, Z.; Khatami, M.; Darroudi, M. Evaluation of antifungal and photocatalytic activities of gelatin-stabilized selenium oxide nanoparticles. J. Inorg. Organomet. Polym. 2020, 30, 3036–3044. [Google Scholar] [CrossRef]
- Rostami, N.; Alidadi, H.; Zarrinfar, H.; Ketabi, D.; Tabesh, H. Interventional Effect of nanosilver paint on fungal load of indoor air in a hospital ward. Can. J. Infect. Dis. Med. Microbiol. 2021, 20, 8658600. [Google Scholar] [CrossRef] [PubMed]
- Alghuthaymi, M.A.; Kalia, A.; Bhardwaj, K.; Bhardwaj, P.; Abd-Elsalam, K.A.; Valis, M.; Kuca, K. Nanohybrid antifungals for control of plant diseases: Current status and future perspectives. J. Fungi. 2021, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Shenashen, M.; Derbalah, A.; Hamza, A.; Mohamed, A.; El Safty, S. Recent trend in controlling root rot disease of tomato caused by Fusarium solani using aluminasilica nanoparticles. Int. J. Adv. Res. Biol. Sci. 2017, 4, 105–119. [Google Scholar]
- Derbalah, A.; Elsharkawy, M.M.; Hamza, A.; El-Shaer, A. Resistance induction in cucumber and direct antifungal activity of zirconium oxide nanoparticles against Rhizoctonia solani. Pestic. Biochem. Physiol. 2019, 157, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Derbalah, A.S.H.; Elsharkawy, M.M. A new strategy to control Cucumber mosaic virus using fabricated NiO-nanostructures. J. Biotechnol. 2019, 306, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Stoimenov, P.K.; Klinger, R.L.; Marchin, G.L.; Klabunde, K.J. Metal oxide nanoparticles as bactericidal agents. Langmuir 2002, 18, 6679–6686. [Google Scholar] [CrossRef]
- Theivasanthi, T.; Alagar, M. Studies of copper nanoparticles effects on micro-Organisms. Ann. Bio. Res. 2011, 2, 368–373. [Google Scholar]
- Chou, W.L.; Yu, D.G.; Yang, M.C. The preparation and characterization of silver-loading cellulose acetate hollow fiber membrane for water treatment. Poly. Adv. Technol. 2005, 16, 600–607. [Google Scholar] [CrossRef]
- Sambhy, V.; MacBride, M.M.; Peterson, B.R.; Sen, A. Silver bromidenanoparticle/polymer composites: Dual action tunable antimicrobial materials. J. Am. Chem. Soc. 2006, 128, 9798–9808. [Google Scholar] [CrossRef]
- Lee, C.; Kim, Y.; Lee, W.I.; Nelson, K.L.; Yoon, J.; Sedlak, D.L. Bactericidal effect of zero-valence iron nanoparticles on Escherichia coli. Environ. Sci. Technol. 2008, 42, 4927–4933. [Google Scholar] [CrossRef] [PubMed]
- Gondal, M.A.; Dastageer, M.A.; Khalil, A.; Hayat, K.; Yamani, Z.H. Nanostructured ZnO synthesis and its application for effective disinfection Escherichia coli microorganism in water. J. Nanopart. Res. 2011, 133, 423–430. [Google Scholar]
- Kavitha, T.; Yuvaraj, H. A facile approach to the synthesis of high-quality NiO nanorods: Electrochemical and antibacterial properties. J. Mater. Chem. 2011, 21, 15686–15691. [Google Scholar] [CrossRef]
- Xiong, L.; Tong, Z.H.; Chen, J.J.; Li, L.L.; Yu, H.Q. Morphology-dependent antimicrobial activity of Cu/Cu𝑥O nanoparticles. Ecotoxicology 2015, 24, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Giannousi, K.; Sarafidis, S.; Mourdikoudis, G.; Pantazaki, A.; Dendrinou-Samara, C. Selective synthesis of Cu2O and Cu/Cu2O NPs: Antifungal activity to yeast saccharomyces cerevisiae and DNA interaction. Inorg. Chem. 2014, 53, 9657–9666. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wang, L.; Liu, L.; Hou, Y. Nanotechnology in agriculture, livestock, and aquaculture in China. “A review”. Agron. Sustain. Dev. 2015, 35, 369–400. [Google Scholar] [CrossRef] [Green Version]
- Khatami, M.; Varma, R.; Heydari, M.; Peydayesh, M.; Sedighi, A.; Askari, H.A.; Rohani, M.; Baniasadi, M.; Arkia, S.; Seyedi, F.; et al. Copper oxide nanoparticles greener aynthesis using tea and its antifungal efficiency on Fusarium solani. Geomicrobiol. J. 2019, 36, 777–781. [Google Scholar] [CrossRef]
- Evans, I.; Solberg, E.; Huber, D.M. Mineral Nutrition and Plant; Datnoff, L.E., Elmer, W.H., Huber, D.N., Eds.; APS Press: Paul, MN, USA, 2007. [Google Scholar]
- Elmer, W.H.; White, J. Nanoparticles of CuO improves growth of eggplant and tomato in disease infested soils. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Borgatta, J.; Ma, C.; Hudson-Smith, N.; Elmer, W.; Pérez, C.D.P.; De La Torre-Roche, R.; Zuverza, M.N.; Haynes, C.L.; White, J.C.; Hamers, R.L. Copper nanomaterials suppress root fungal disease in watermelon (Citrullus lanatus): Role of particle morphology, composition, and dissolution behavior. CS Sustain. Chem. Eng. 2018, 6, 14847–14856. [Google Scholar] [CrossRef]
- Elmer, W.H.; De La Torre-Roche, R.; Pagano, L.; Majumdar, S.; Zuverza-Mena, N.; Dimpka, C.; Gardea-Torresdey, J.; White, W. Effect of metalloid and metallic oxide nanoparticles on Fusarium wilt of watermelon. Plant. Dis. 2018, 102, 1394–1401. [Google Scholar] [CrossRef]
- Nelson, E.B.; Boehm, M.J. Compost-induced suppression of turf grass diseases. Bio. Cycle 2002, 43, 51. [Google Scholar]
- Ismail, W.; El-Shafai, N.M.; El-Shaer, A.; Abdelfatah, M. Impact of substrate type on the surface and properties of electrodeposited Cu2O nanostructure films as an absorber layer for solar cell applications. Mater. Sci Semicond. Process. 2020, 120, 105335. [Google Scholar] [CrossRef]
- Booth, C. The Genus Fusarium, 2nd ed.; Commonwealth Mycological Institute: Surrey, UK, 1971; p. 237. [Google Scholar]
- Allen, O.N. Experiments in Soil Bacteriology; Burgess Publishing Co.: Minneapolis, MN, USA, 1950. [Google Scholar]
- Shaban, W.I.; El-Bramawy, M.A. Impact of dual inoculation with Rhizobium and Trichoderma on damping-off, root rot diseases and plant growth parameters of some legumes field crop under greenhouse conditions. Int. Res. J. Agric. Sci. Soil. Sci. 2011, 1, 98–108. [Google Scholar]
- Hwang, S.F.; Chang, K.F. Incidence and severity of root rot disease complex of field pea in northern Alberta. Can. Plant Dis Surv 1989, 69, 139–141. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum Lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Emynology and Histoenzymology, Kalyani Publishers. Indian Print. Navin. Shanndara Delhi 1980, 54–56. [Google Scholar]
- Tek, M.I.; Calis, O. Mechanisms of resistance to powdery mildew in cucumber. Phytopathol. Mediterr. 2022, 61, 119–127. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Harley, M.M.; Ferguson, I.K. The role of the SEM in pollen morphology and plant systematics. In Scanning Electron Microscopy in Taxonomy and Functional Morphology; Oxford University Press: Oxford, England, 1990; pp. 45–68. [Google Scholar]
- Nassar, M.A.; El-Sahar, K.F. Botanical Preparations and Microscopy (Micro Technique); Academic Bookshop: Giza Governorate, Egypt, 1998; p. 219. [Google Scholar]
- Torres-Netto, A.; Campostrini, E.; Oliveira, J.G.; Smith, R.E.B. Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Sci. Hortic. 2005, 104, 199–209. [Google Scholar] [CrossRef]
- El-Shaer, A.; Ismail, W.; Abdelfatah, M. Towards low cost fabrication of inorganic white light emitting diode based on electrodeposited Cu2O thin film/TiO2 nanorods heterojunction. Mater. Res. Bull. 2019, 116, 111–116. [Google Scholar] [CrossRef]
- Abdelfatah, M.; Ismail, W.; El-Shaer, A. Low cost inorganic white light emitting diode based on submicron ZnO rod arrays and electrodeposited Cu2O thin film. Mater. Sci. Semicond. Process. 2018, 81, 44–47. [Google Scholar] [CrossRef]
- El-Shafai, N.M.; Abdelfatah, M.; El-Mehasseb, I.M.; Ramadan, M.S.; Ibrahim, M.M.; El-Shaer, A.; El-Kemary, M.A.; Masoud, M.S. Enhancement of electrochemical properties and photocurrent of copper oxide by heterojunction process as a novel hybrid nanocomposite for photocatalytic anti-fouling and solar cell applications. Sep. Purif. Technol. 2021, 267, 118631. [Google Scholar] [CrossRef]
- Abdelfatah, M.; Salah, H.; El-Henawey, M.; Oraby, A.; El-Shaer, A.; Ismail, W. Insight into co concentrations effect on the structural, optical, and photoelectrochemical properties of ZnO rod arrays for optoelectronic applications. J. Alloy Compd. 2021, 873, 159875. [Google Scholar] [CrossRef]
- El-Shaer, A.; Abdelfatah, K.R.; Mahmoud, S.; Eraky, M. Correlation between photoluminescence and positron annihilation lifetime spectroscopy to characterize defects in calcined MgO nanoparticles as a first step to explain antibacterial activity. J. Alloy Compd. 2020, 817, 152799. [Google Scholar] [CrossRef]
- El-Shafai, N.; Shukry, M.M.; El-Mehasseb, I.M.; Abdelfatah, M.; Ramadan, M.S.; El-Shaer, A.; El-Kemary, M. Electrochemical property, antioxidant activities, water treatment and solar cell applications of titanium dioxide-zinc oxide hybrid nanocomposite based on graphene oxide nanosheet. Mater. Sci. Eng. B-Adv. 2020, 259, 114596. [Google Scholar] [CrossRef]
- Ziedan, E.H.E.; Moataza, S. Efficacy of nanoparticles on seed borne fungi and their pathological potential of cucumber. Inter. J. PharmTech. Res. 2016, 9, 16–24. [Google Scholar]
- Consolo, V.F.; AndrésTorres-Nicolini, A.T.; Alvarez, V.A. Mycosinthetized Ag, CuO and ZnO nanoparticles from a promising Trichoderma harzianum strain and their antifungal potential against important phytopathogens. Nat. Res. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Franich, R.A. Chemistry of weathering and solubilisation of copper fungicide and the effect of copper on germination, growth, metabolism, and reproduction of Dothistroma pini. New Zealand J. For. Sci. 1988, 18, 318–328. [Google Scholar]
- Freeman, M.H.; McIntyre, C.R. A comprehensive review of cooper-based wood preservatives with a focus on new micronized or dispersed copper systems. For. Prod. J. 2008, 58, 6–27. [Google Scholar]
- Jamdagni, P.; Khatri, P.; Rana, J.S. Green synthesis of zinc oxide nanoparticles using flower extract of Nyctanthes arbor-tristis and their antifungal activity. J. King Saud. Univ. Sci. 2018, 30, 168–175. [Google Scholar] [CrossRef]
- Babaei, S.; Bajelani, F.; Mansourizaveleh, O.; Abbasi, A.; Oubari, F. A study of the bactericidal effect of copper oxide nanoparticles on Shigella sonnei and Salmonella typhimurium. J. Babol. Univ. Med. Sci. 2017, 19, 76–81. [Google Scholar]
- Hamedi, S.; Shojaosadati, S.A.; Mohammadi, A. Evaluation of the catalytic, antibacterial and anti-biofilm activities of the Convolvulus arvensis extract functionalized silver nanoparticles. J. Photochem. Photobiol. B 2017, 167, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Karthik, K.; Dhanuskodi, S.; Gobinath, C.; Prabukumar, S.; Sivaramakrishnan, S. Photocatalytic and antibacterial activities of hydrothermally prepared cdo nanoparticles. J. Mater. Sci. 2017, 28, 11420–11429. [Google Scholar] [CrossRef]
- Khorrami, M.B.; Sadeghnia, H.R.; Pasdar, A.; Ghayour-Mobarhan, M.; RiahiZanjani, B.; Darroudi, M. Role of pullulan in preparation of ceria nanoparticles and investigation of their biological activities. J. Mol. Struct. 2018, 1157, 127–131. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.; Cui, J. Graphene oxide loaded with copper oxide nanoparticles as an antibacterial agent against Pseudomonas syringae pv. tomato. RSC Adv. 2017, 7, 38853–38860. [Google Scholar] [CrossRef]
- Ashraf, H.; Anjum, T.; Riaz, S.; Ahmad, I.S.; Irudayaraj, J.; Javed, S.; Qaiserf, U.; Shahzad Naseem, S. Inhibition mechanism of green-synthesized copper oxide nanoparticles from Cassia fistula towards Fusarium oxysporum by boosting growth and defense response in tomatoes. Env. Sci. Nano 2021, 8, 1729–1748. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon nanotubes induce growth enhancement of tobacco cells. Am. Chem. Soc. Nano 2012, 3, 2128–2135. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Omara, R.I.; Mostafa, Y.S.; Alamri, S.A.; Hashem, A.; Alrumman, S.; Ahmed, A.A. Mechanism of wheat leaf rust control using chitosan nanoparticles and salicylic acid. J. Fungi 2022, 8, 304. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kim, K.S.; Lamsal, K.; Kim, Y.J.; Kim, S.B.; Jung, M.; Sim, S.J.; Kim, H.S.; Chang, S.J.; Kim, H.K.; et al. An in vitro study of the antifungal effect of silver nanoparticles on oak wilt pathogen Raffaelea sp. J. Microbio.l Biotechnol. 2009, 19, 760–764. [Google Scholar]
- Yasmeen, F.; Raja, N.I.; Razzaq, A.; Komatsu, S. Proteomic and physiological analyses of wheat seeds exposed to copper and iron nanoparticles. Biochim. Biophys. Acta 2017, 1865, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.A.; Huang, Y.; Nelson, J. Detection of nanoparticles in edible plant tissues exposed to nano-copper using single-particle ICP-MS. J. Nanopart. Res. 2018, 20, 1–13. [Google Scholar] [CrossRef]
- Su, Y.; Zheng, X.; Chen, Y.; Li, M.; Liu, K. Alteration of intracellular protein expressions as a key mechanism of the deterioration of bacterial denitrification caused by copper oxide nanoparticles. Sci. Rep. 2015, 5, 15824. [Google Scholar] [CrossRef]
- Goyal, P.; Chakraborty, S.; Misra, S.K. Multifunctional Fe3O4-ZnO nanocomposites for environmental remediation applications. Environ. Nanotechnol. Monit. Manag. 2018, 10, 28–35. [Google Scholar] [CrossRef]
- Dreyer, D.R.; Park, S.; Bielawski, C.W.; Ruoff, R.S. The chemistry of graphene oxide. Chem. Soc. Rev. 2010, 39, 228–240. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (′5–′3) | Reverse Primer (′5–′3) |
|---|---|---|
| PR-1 | TGCTCAACAATATGCGAACC | TCATCCACCCACAACTGAAC |
| LOX-1 | CTCTTGGGTGGTGGTGTTTC | TGGTGGGATTGAAGTTAGCC |
| Actin | TGCTGGTCGTGACCTTACTG | GAATCTCTCAGCTCCGATGG |
| Treatment | Conc. (µg/L) | Radial Growth (mm) | Inhibition % | Regression Equation | R² |
|---|---|---|---|---|---|
| Cu2ONPs (0.35) | 10 | 7.0 b ±0.54 * | 22.22 f ± 0.56 | Y = 1.7611x − 33.49 | 0.99 |
| 25 | 5.8 c ± 0.66 | 35.56 e ± 0.76 | |||
| 50 | 4.6 d ±0.48 | 48.89 d ±0.73 | |||
| 100 | 2.3 d ± 0.52 | 74.44 b± 0.86 | |||
| Cu2ONPs (0.30) | 10 | 6.8 c ± 0.64 | 24.44 d± 0.78 | Y = 1.6779x − 36.244 | 0.981 |
| 25 | 5.4 e± 0.57 | 38.89 c ± 0.91 | |||
| 50 | 4.1 f ± 0.43 | 54.44 b ±0.93 | |||
| 100 | 1.9 f ± 0.39 | 78.89 a± 1.12 | |||
| Fungicide (Uniform 390 SE) | 10 | 6.3 d ± 0.67 | 30.0 d ± 0.71 | Y = 1.627x − 43.269 | 0.983 |
| 25 | 5.1 e ± 0.55 | 43.33 c ± 0.89 | |||
| 50 | 3.5 f ± 0.45 | 61.11 b± 1.07 | |||
| 100 | 1.3 f ± 0.41 | 85.56 a± 1.14 | |||
| Control | 0 | 9.0 a ± 0.69 | 0.0 g ± 0.19 |
| Treatment | Damping-Off % | Disease Incidence% | % Efficacy | ||
|---|---|---|---|---|---|
| Pre-Emergence | Post-Emergence | Survival | |||
| Cu2ONPs (0. 30) | 13.3 c ± 0.68 * | 6.0 c ± 0.26 | 80.7 a ±1.35 | 26.9 c ± 0.57 | 50.0 |
| Cu2ONPs (0.35) | 30.0 a ± 0.89 | 0.0 d ± 0.02 | 70.0 c ± 1.23 | 28.6 b ± 0.63 | 46.8 |
| Fungicide (Uniform 390 SE) | 13.3 c ± 0.63 | 13.3 b ± 0.46 | 73.4 b ± 1.41 | 20.0 d ± 0.48 | 62.8 |
| Control | 17.5 b ± 0.47 | 17.5 a ± 0.53 | 65.0 d ± 1.12 | 53.8 a ± 0.63 | 0.0 |
| Treatments | Enzyme Activity | ||
|---|---|---|---|
| CAT (Catalase) mM H2O2 g−1 FW Min−1 | POX (Peroxidase) mM H2O2 g−1 FW Min−1 | PPO (Polyphenol Oxidase) µ mol/min−1 g−1 (FW) | |
| Cu2ONPs (0.30) | 23.3 a ± 0.63 * | 1.397 a ± 0.23 | 0.127 a ± 0.52 |
| Cu2ONPs (0.35) | 22.1 b ± 0.61 | 1.239 a ± 0.28 | 0.098 b ± 0.43 |
| Fungicide (Uniform 390 SE) | 19.7 c ± 0.46 | 0.741 b ± 0.25 | 0.054 c ± 0.35 |
| Control | 12.8 d ± 0.42 | 0.329 c ± 0.21 | 0.034 d ± 0.22 |
| Treatments | Menoufia Governorate | ||||
|---|---|---|---|---|---|
| Damping-Off % | Disease Incidence % | % Efficacy | |||
| Pre-Emergence | Post-Emergence | Survival | |||
| Cu2ONPs (0.35) | 12.0 b ± 0.65 | 18.7 c ± 0.68 | 69.3 b ± 1.23 | 24.7 b ± 0.54 | 63.8 |
| Cu2ONPs (0.30) | 9.3 c ± 0.56 | 10.3 b ± 0.54 | 80.4 a ± 1.41 | 21.9 c ± 0.46 | 67.9 |
| Fungicide (Uniform 390 SE) | 7.3 c ± 0.51 | 11.3 b ± 0.57 | 81.4 a ± 1.46 | 18.0 d ± 0.44 | 73.6 |
| Control | 14.5a ± 0.72 | 29.2 a ± 0.81 | 56.3 c ± 1.27 | 68.3 a ± 1.75 | 0.0 |
| Giza Governorate | |||||
| Cu2ONPs (0.35) | 9.7 b ± 0.58 | 12.7 c ± 0.59 | 77.6 c ± 1.34 | 21.3 b ± 0.58 | 70.17 |
| Cu2ONPs (0.30) | 7.4 c ± 0.51 | 11.3 b ± 0.56 | 81.6 b ± 1.42 | 20.4 b ± 0.66 | 71.43 |
| Fungicide (Uniform 390 SE) | 5.7 c ± 0.47 | 10.3 b ± 0.51 | 84.0 a ± 1.44 | 17.3 c ± 0.51 | 75.77 |
| Control | 13.5 a ± 0.62 | 31.7 a ± 0.74 | 54.8 d ± 1.04 | 71.4 a ± 1.42 | 0.0 |
| Treatment | Total Chlorophyll (SPAD) | Shoot Length (cm) | Root Length (cm) | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|---|
| Menoufia Governorate | |||||
| Cu2ONPs (0.35) | 32.1 b ± 0.74 | 176.4 a ± 1.21 | 30.8 b ± 0.85 | 45.2 a ± 0.67 | 4.8 b ± 0.23 |
| Cu2ONPs (0.30) | 33.7 b ± 0.77 | 177.2 a ± 1.24 | 31.1 b ± 0.82 | 47.7 b ± 0.86 | 5.2 b ± 0.23 |
| Fungicide (Uniform 390 SE) | 35.5 a ± 0.71 | 178.8 a ± 1.32 | 32.9 a ± 0.79 | 47.3 a ± 0.90 | 6.7 a ± 0.31 |
| Control | 23.4 c ± 0.59 | 130.3 b ± 1.21 | 23.2 c ± 0.98 | 29.3 c ± 0.47 | 3.3 c ± 0.22 |
| Giza Governorate | |||||
| Cu2ONPs (0.35) | 35.7 a ± 0.73 | 181.3 b ± 1.34 | 31.2 b ± 0.79 | 48.9 a ± 0.76 | 6.1 a ± 0.43 |
| Cu2ONPs (0.30) | 36.1 a ± 0.74 | 182.4 a ± 1.43 | 33.2 a ± 0.88 | 49.3 a ± 0.56 | 6.4 a ± 0.46 |
| Fungicide (Uniform 390 SE) | 36.5 a ± 0.68 | 179.3 b ± 1.44 | 33.9 a ± 0.81 | 50.7 a ± 0.67 | 6.3 a ± 0.42 |
| Control | 24.4 b ± 0.42 | 131.8 c ± 1.07 | 25.1 c ± 0.57 | 28.1 b ± 0.37 | 3.5 c ± 0.32 |
| Treatments | No. of Fruits | Mean Weight of Fruits (g) | Fruits Weight/Plant (kg) | % Rate of Yield Increase ** |
|---|---|---|---|---|
| Menoufia Governorate | ||||
| Cu2ONPs (0.35) | 27.7 b ± 0.78 * | 73.4 b ± 0.99 | 2.033 b ± 0.34 | 96.4 c ± 1.03 |
| Cu2ONPs (0.30) | 28.3 a ± 0.76 | 74.2 a ± 0.95 | 2.099 b ± 0.37 | 102.8 b ± 1.11 |
| Fungicide (Uniform 390 SE) | 29.4 a ± 0.79 | 74.8 a ± 0.97 | 2.199 a ± 0.29 | 112.5 a ±1.14 |
| Control | 14.3 c ± 0.49 | 72.4 c ± 0.93 | 1.035 c ± 0.19 | 0.00 d ± 0.78 |
| Giza Governorate | ||||
| Cu2ONPs (0.35) | 28.3 b ± 0.89 | 74.1 a ± 0.94 | 2.097 b ± 0.37 | 89.7 c ± 0.99 |
| Cu2ONPs (0.30) | 29.4 a ± 0.87 | 74.9 a ± 0.98 | 2.202 a ± 0.43 | 99.3 b ± 0.94 |
| Fungicide | 29.7 a ± 0.84 | 75.2 a ± 1.04 | 2.233 a ± 0.34 | 102.1a ± 1.05 |
| Control (Uniform 390 SE) | 15.4 c ± 0.53 | 71.8 c ± 0.97 | 1.105 c ± 0.29 | 0.0 d ± 1.01 |
| Efficacy | Total Chlorophyll | Shoot Length | Root Length | Fresh Weight | Dry Weight | No. of Fruits | Mean Weight of Fruits | Fruits Weight/Plant | Rate of Yield Increase | |
|---|---|---|---|---|---|---|---|---|---|---|
| Efficacy | 1 | 0.978 ** | 0.995 ** | 0.972 ** | 0.989 ** | 0.914 ** | 0.998 ** | 0.923 ** | 0.998 ** | 0.991 ** |
| Total chlorophyll | 1 | 0.976 ** | 0.978 ** | 0.987 ** | 0.965 ** | 0.978 ** | 0.921 ** | 0.980 ** | 0.952 ** | |
| Shoot length | 1 | 0.961 ** | 0.988 ** | 0.904 ** | 0.995 ** | 0.896 ** | 0.993 ** | 0.983 ** | ||
| Root length | 1 | 0.963 ** | 0.943 ** | 0.979 ** | 0.941 ** | 0.983 ** | 0.960 ** | |||
| Fresh weight | 1 | 0.917 ** | 0.986 ** | 0.911 ** | 0.986 ** | 0.968 ** | ||||
| Dry weight | 1 | 0.914 ** | 0.922 ** | 0.920 ** | 0.888 ** | |||||
| No. of fruits | 1 | 0.916 ** | 1.000 ** | 0.992 ** | ||||||
| Mean weight of fruits | 1 | 0.927 ** | 0.904 ** | |||||||
| Fruits weight/plant | 1 | 0.991 ** | ||||||||
| Rate of yield increase | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamel, S.M.; Elgobashy, S.F.; Omara, R.I.; Derbalah, A.S.; Abdelfatah, M.; El-Shaer, A.; Al-Askar, A.A.; Abdelkhalek, A.; Abd-Elsalam, K.A.; Essa, T.; et al. Antifungal Activity of Copper Oxide Nanoparticles against Root Rot Disease in Cucumber. J. Fungi 2022, 8, 911. https://doi.org/10.3390/jof8090911
Kamel SM, Elgobashy SF, Omara RI, Derbalah AS, Abdelfatah M, El-Shaer A, Al-Askar AA, Abdelkhalek A, Abd-Elsalam KA, Essa T, et al. Antifungal Activity of Copper Oxide Nanoparticles against Root Rot Disease in Cucumber. Journal of Fungi. 2022; 8(9):911. https://doi.org/10.3390/jof8090911
Chicago/Turabian StyleKamel, Said M., Samah F. Elgobashy, Reda I. Omara, Aly S. Derbalah, Mahmoud Abdelfatah, Abdelhamed El-Shaer, Abdulaziz A. Al-Askar, Ahmed Abdelkhalek, Kamel A. Abd-Elsalam, Tarek Essa, and et al. 2022. "Antifungal Activity of Copper Oxide Nanoparticles against Root Rot Disease in Cucumber" Journal of Fungi 8, no. 9: 911. https://doi.org/10.3390/jof8090911