
Evaluation of the Potential Entomopathogenic Fungi Purpureocillium lilacinum and Fusarium verticillioides for Biological Control of Forcipomyia taiwana (Shiraki)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Forcipomyia Taiwana Collection and Rearing
2.2. Fungal Isolation and Selection
2.3. Molecular Identification
2.4. Phylogenetic Analysis
2.5. Thermotolerance Assay
2.6. Conidia Productivity
2.7. Ranking of EPF Isolates by Effective Conidia Number (ECN)
2.8. Virulence Screening against F. taiwana Last Instar Larvae
2.9. Bioassay of EPF against Different Life Stages of F. taiwana
2.10. Statistical Analysis
3. Results
3.1. Isolation and Selection of Entomopathogenic Fungi
3.2. Molecular Identification
3.3. ECN Ranking of the Entomopathogenic Fungi
3.4. Virulence Screening against F. taiwana Larvae
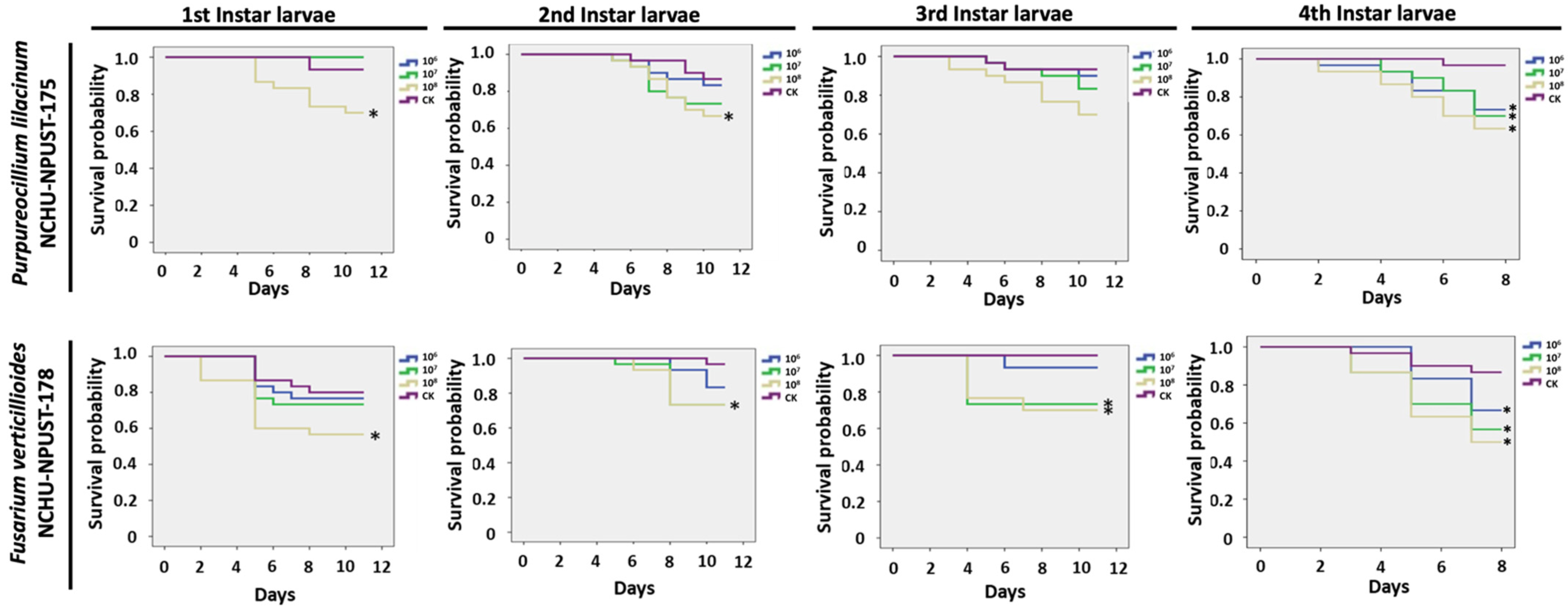
3.5. Bioassay of EPF against Different Life Stages of F. taiwana
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chuang, Y.-Y.; Lin, C.-S.; Wang, C.-H.; Yeh, C.-C. Distribution and seasonal occurrence of Forcipomyia taiwana (Diptera: Ceratopogonidae) in the Nantou area in Taiwan. J. Med. Entomol. 2000, 37, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Chou, J.-Y.; Lin, C.-C.; Wen, Y.-D.; Wang, W.-L. Seasonal yeast compositions in Forcipomyia taiwana (Diptera: Ceratopogonidae). J. Asia-Pac. Entomol. 2016, 19, 509–514. [Google Scholar] [CrossRef]
- Luo, Y.P. Establishing and maintaining colonies of Forcipomyia taiwana in the laboratory. J. Vector Ecol. 2018, 43, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-E.; Tsai, M.-H.; Huang, H.-T.; Tsai, C.-C.; Chen, M.-J.; Yang, D.-S.; Yang, T.-Z.; Wang, J.; Huang, R.-N. Transcriptome profiling reveals the developmental regulation of NaCl-treated Forcipomyia taiwana eggs. BMC Genom. 2021, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Pope, E.C.; Carpenter, S.; Scholte, E.-J.; Butt, T.M. Entomopathogenic fungus as a biological control for an important vector of livestock disease: The Culicoides biting midge. PLoS ONE 2011, 6, e16108. [Google Scholar] [CrossRef]
- Scholte, E.-J.; Knols, B.G.; Samson, R.A.; Takken, W. Entomopathogenic fungi for mosquito control: A review. J. Insect Sci. 2004, 4, 19. [Google Scholar] [CrossRef]
- Glare, T.R.; O’Callaghan, M. Environmental and health impacts of Bacillus thuringiensis israelensis. Rep. Minist. Health 1998, 57, 3–58. [Google Scholar]
- Helyer, N.; Gill, G.; Bywater, A.; Chambers, R. Elevated humidities for control of chrysanthemum pests with Verticillium lecanii. Pestic. Sci. 1992, 36, 373–378. [Google Scholar] [CrossRef]
- Inglis, G.D.; Johnson, D.L.; Goettel, M.S. Effects of temperature and sunlight on mycosis (Beauveria bassiana) (Hyphomycetes: Sympodulosporae) of grasshoppers under field conditions. Environ. Entomol. 1997, 26, 400–409. [Google Scholar] [CrossRef]
- Castillo Lopez, D.; Zhu-Salzman, K.; Ek-Ramos, M.J.; Sword, G.A. The entomopathogenic fungal endophytes Purpureocillium lilacinum (formerly Paecilomyces lilacinus) and Beauveria bassiana negatively affect cotton aphid reproduction under both greenhouse and field conditions. PLoS ONE 2014, 9, e103891. [Google Scholar] [CrossRef]
- Teetor-Barsch, G.H.; Roberts, D.W. Entomogenous Fusarium species. Mycopathologia 1983, 84, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Boetel, M.A.; Jaronski, S.T. Discovery of Fusarium solani as a naturally occurring pathogen of sugarbeet root maggot (Diptera: Ulidiidae) pupae: Prevalence and baseline susceptibility. J. Invertebr. Pathol. 2008, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Marques, G. Fusarium, an entomopathogen—A myth or reality? Pathogens 2018, 7, 93. [Google Scholar] [CrossRef]
- Chang, J.-C.; Wu, S.-S.; Liu, Y.-C.; Yang, Y.-H.; Tsai, Y.-F.; Li, Y.-H.; Tseng, C.-T.; Tang, L.-C.; Nai, Y.-S. Construction and selection of an entomopathogenic fungal library from soil samples for controlling Spodoptera litura. Front. Sustain. Food Syst. 2021, 5, 596316. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Ni, N.-T.; Chang, J.-C.; Li, Y.-H.; Lee, M.R.; Kim, J.S.; Nai, Y.-S. Isolation and Selection of Entomopathogenic Fungi from Soil Samples and Evaluation of Fungal Virulence against Insect Pests. J. Vis. Exp. JOVE 2021, 175, 62882. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Nosratabadi, M.; Kachuei, R.; Rezaie, S.; Harchegani, A.B. Beta-tubulin gene in the differentiation of Fusarium species by PCR-RFLP analysis. Infez Med. 2018, 26, 52–60. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, M.R.; Kim, S.; Lee, S.J.; Park, S.E.; Nai, Y.-S.; Lee, G.S.; Shin, T.Y.; Kim, J.S. Tenebrio molitor-mediated entomopathogenic fungal library construction for pest management. J. Asia-Pac. Entomol. 2018, 21, 196–204. [Google Scholar] [CrossRef]
- Pelizza, S.A.; Stenglein, S.A.; Cabello, M.N.; Dinolfo, M.I.; Lange, C.E. First record of Fusarium verticillioides as an entomopathogenic fungus of grasshoppers. J. Insect Sci. 2011, 11, 1–8. [Google Scholar] [CrossRef]
- Sun, T.; Wu, J.; Ali, S. Morphological and molecular identification of four Purpureocillium isolates and evaluating their efficacy against the sweet potato whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control. 2021, 31, 1–9. [Google Scholar] [CrossRef]
- Bush, B.; Carson, M.; Cubeta, M.; Hagler, W.; Payne, G. Infection and fumonisin production by Fusarium verticillioides in developing maize kernels. Phytopathology 2004, 94, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Howard, R.J. Biology of maize kernel infection by Fusarium verticillioides. Mol. Plant-Microbe Interact. 2010, 23, 6–16. [Google Scholar] [CrossRef]
- da Silva Santos, A.C.; Diniz, A.G.; Tiago, P.V.; de Oliveira, N.T. Entomopathogenic Fusarium species: A review of their potential for the biological control of insects, implications and prospects. Fungal Biol. Rev. 2020, 34, 41–57. [Google Scholar] [CrossRef]
- Sain, S.K.; Monga, D.; Hiremani, N.S.; Nagrale, D.T.; Kranthi, S.; Kumar, R.; Kranthi, K.R.; Tuteja, O.; Waghmare, V.N. Evaluation of bioefficacy potential of entomopathogenic fungi against the whitefly (Bemisia tabaci Genn.) on cotton under polyhouse and field conditions. J. Invertebr. Pathol. 2021, 183, 107618. [Google Scholar] [CrossRef]
- Lee, W.W.; Shin, T.Y.; Bae, S.M.; Woo, S.D. Screening and evaluation of entomopathogenic fungi against the green peach aphid, Myzus persicae, using multiple tools. J. Asia-Pac. Entomol. 2015, 18, 607–615. [Google Scholar] [CrossRef]
- Fang, W.; Bidochka, M.J. Expression of genes involved in germination, conidiogenesis and pathogenesis in Metarhizium anisopliae using quantitative real-time RT-PCR. Mycol. Res. 2006, 110, 1165–1171. [Google Scholar] [CrossRef]
- Shin, T.Y.; Lee, M.R.; Park, S.E.; Lee, S.J.; Kim, W.J.; Kim, J.S. Pathogenesis-related genes of entomopathogenic fungi. Arch. Insect Biochem. Physiol. 2020, 105, e21747. [Google Scholar] [CrossRef]
- Akutse, K.S.; Kimemia, J.W.; Ekesi, S.; Khamis, F.M.; Ombura, O.L.; Subramanian, S. Ovicidal effects of entomopathogenic fungal isolates on the invasive fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2019, 143, 626–634. [Google Scholar] [CrossRef]
- Usman, M.; Wakil, W.; Piñero, J.C.; Wu, S.; Toews, M.D.; Shapiro-Ilan, D.I. Evaluation of Locally Isolated Entomopathogenic Fungi against Multiple Life Stages of Bactrocera zonata and Bactrocera dorsalis (Diptera: Tephritidae): Laboratory and Field Study. Microorganisms 2021, 9, 1791. [Google Scholar] [CrossRef]
- Idrees, A.; Qadir, Z.A.; Akutse, K.S.; Afzal, A.; Hussain, M.; Islam, W.; Waqas, M.S.; Bamisile, B.S.; Li, J. Effectiveness of entomopathogenic fungi on immature stages and feeding performance of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae. Insects 2021, 12, 1044. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Singh, G.; Soni, N.; Sharma, S. Pathogenicity of Fusarium oxysporum against the larvae of Culex quinquefasciatus (Say) and Anopheles stephensi (Liston) in laboratory. Parasitol. Res. 2010, 107, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Mohamed, H.; El-Naggar, S.; Swelim, M.; Elkhawaga, O. Isolation and selection of entomopathogenic fungi as biocontrol agent against the greater wax moth, Galleria mellonella L.(Lepidoptera: Pyralidae). Egypt. J. Biol. Pest Control 2016, 26, 249. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, N.-T.; Wu, S.-S.; Liao, K.-M.; Tu, W.-C.; Lin, C.-F.; Nai, Y.-S. Evaluation of the Potential Entomopathogenic Fungi Purpureocillium lilacinum and Fusarium verticillioides for Biological Control of Forcipomyia taiwana (Shiraki). J. Fungi 2022, 8, 861. https://doi.org/10.3390/jof8080861
Ni N-T, Wu S-S, Liao K-M, Tu W-C, Lin C-F, Nai Y-S. Evaluation of the Potential Entomopathogenic Fungi Purpureocillium lilacinum and Fusarium verticillioides for Biological Control of Forcipomyia taiwana (Shiraki). Journal of Fungi. 2022; 8(8):861. https://doi.org/10.3390/jof8080861
Chicago/Turabian StyleNi, Nian-Tong, Sing-Shan Wu, Kuei-Min Liao, Wu-Chun Tu, Chuen-Fu Lin, and Yu-Shin Nai. 2022. "Evaluation of the Potential Entomopathogenic Fungi Purpureocillium lilacinum and Fusarium verticillioides for Biological Control of Forcipomyia taiwana (Shiraki)" Journal of Fungi 8, no. 8: 861. https://doi.org/10.3390/jof8080861