
Postantifungal Effect of Antifungal Drugs against Candida: What Do We Know and How Can We Apply This Knowledge in the Clinical Setting?

 , , ,
, , ,
Abstract
:1. Introduction
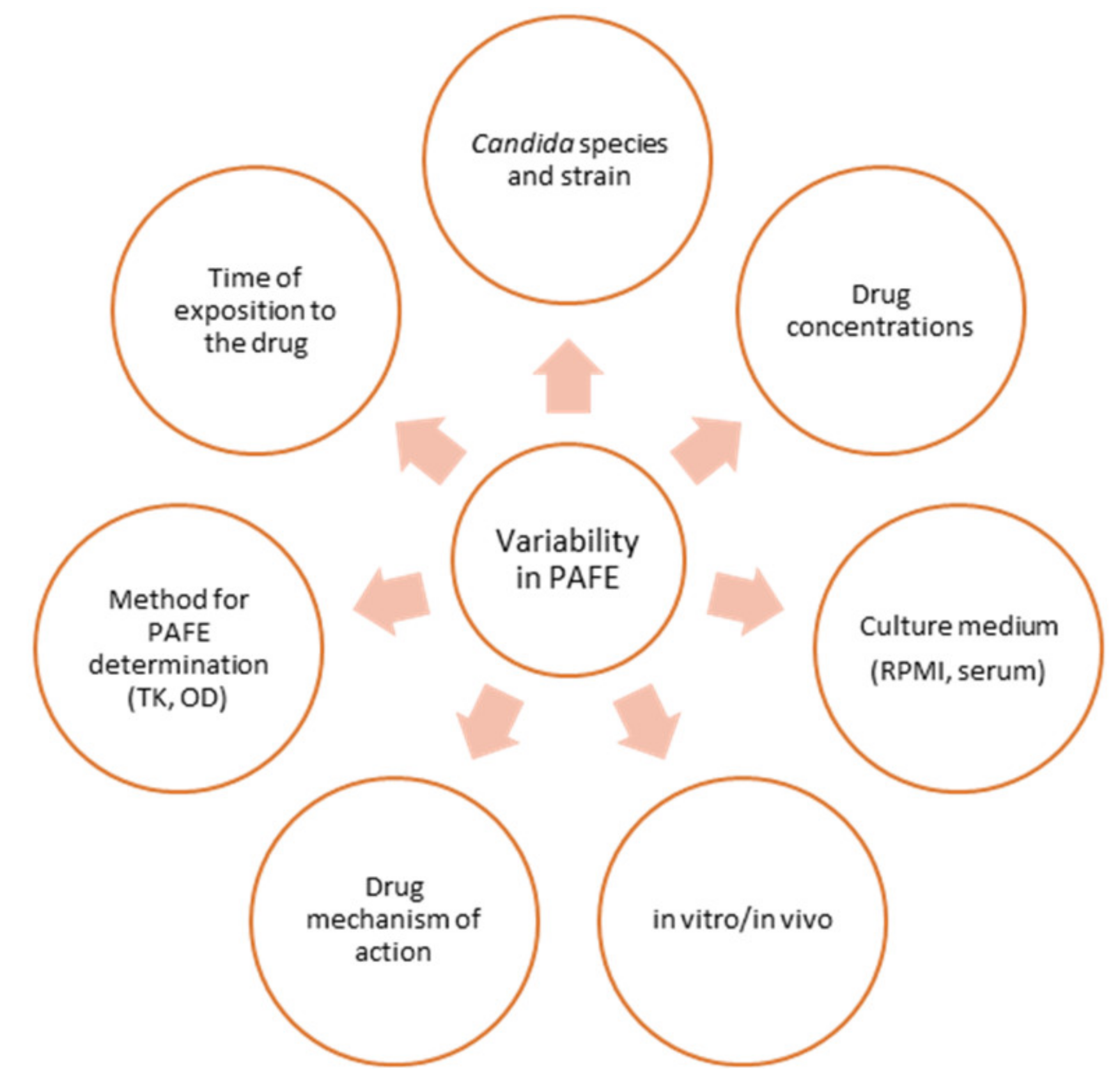
2. In Vitro and In Vivo Methods for PAFE Study
2.1. In Vitro Methods for PAFE Study
2.1.1. PAFE Determination by Optical Density Measurement
Data Analysis
Determination of Carry-Over Effect
2.1.2. PAFE Determination by Colony Counting
Time-Kill Assays
PAFE Assays
Data Analysis
Determination of Carry-Over Effect
In Vitro PAFE of the Combinations between Antifungal Drugs
In Vitro PAFE in Presence of Serum
2.2. In Vivo Methods for PAFE Study
2.2.1. Murine Models of Disseminated Candidiasis
2.2.2. In Vivo Time-Kill and PAFE Assays
Data Analysis
3. PAFE of Polyenes
4. PAFE of Azoles
5. PAFE of Echinocandins
6. PAFE of 5-Fluorocytosine
7. Mechanisms Involved in PAFE
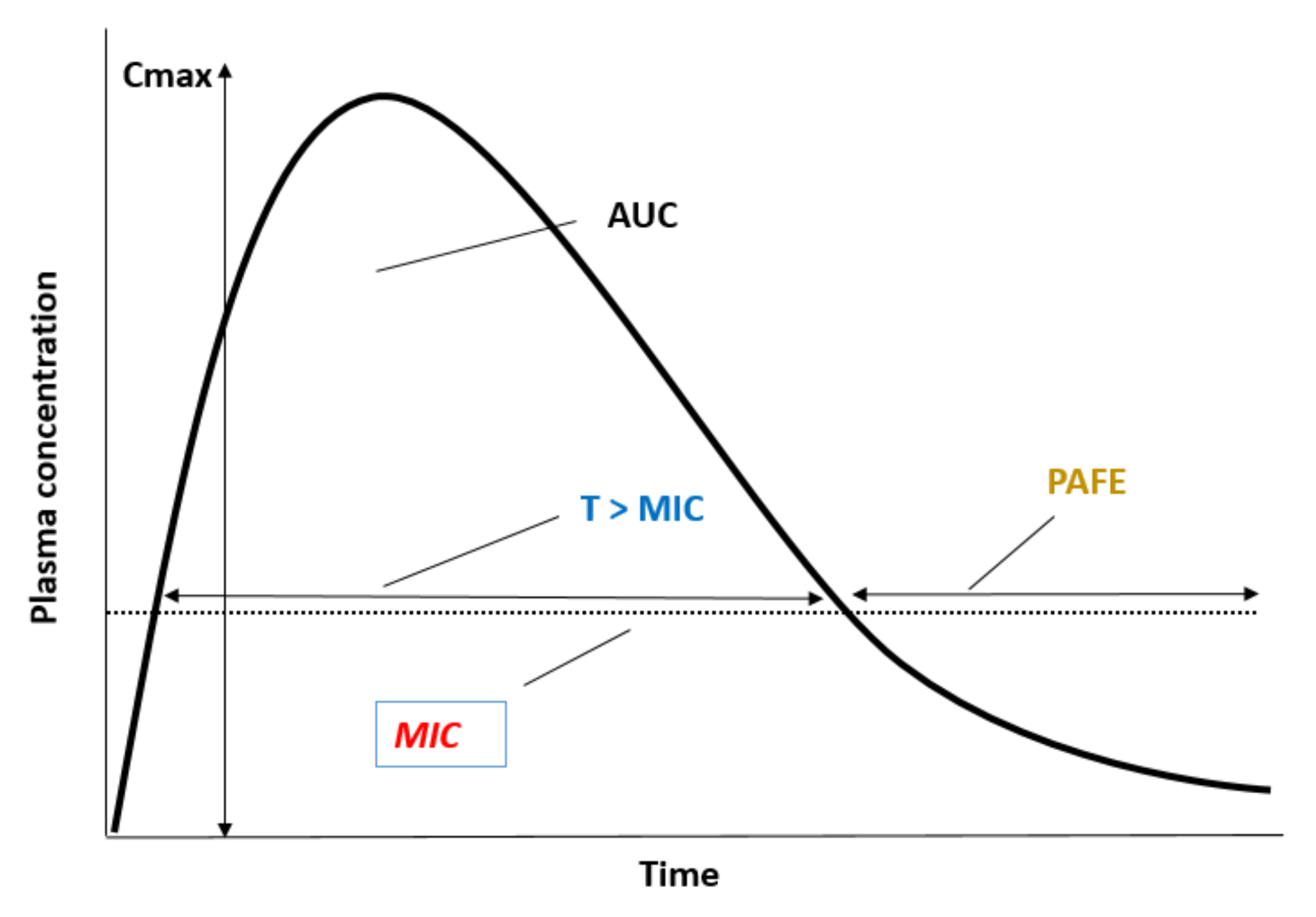
8. Relevance of PAFE in the Design of Dosing Therapeutical Regimens: Clinical Applicability
9. Conclusions and Proposals for Further Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quindós, G. Micología Médica, 1st ed.; Elsevier: Barcelona, Spain, 2015. [Google Scholar]
- Gumbo, T.; Drusano, G.L.; Liu, W.; Kulawy, R.W.; Fregeau, C.; Hsu, V.; Louie, A. Once-weekly micafungin therapy is as effective as daily therapy for disseminated candidiasis in mice with persistent neutropenia. Antimicrob. Agents Chemother. 2007, 51, 968–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oz, Y.; Kiremitci, A.; Dag, I.; Metintas, S.; Kiraz, N. Postantifungal effect of the combination of caspofungin with voriconazole and amphotericin B against clinical Candida krusei isolates. Med. Mycol. 2013, 51, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chryssanthou, E.; Cars, O.; Sjölin, J. New automated method for determining postantifungal effect of amphotericin B against Candida species: Effects of concentration, exposure time, and area under the curve. Antimicrob. Agents Chemother. 2002, 46, 4016–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellepola, A.N.; Samaranayake, L.P. The postantifungal effect (PAFE) of antimycotics on oral C. albicans isolates and its impact on candidal adhesion. Oral Dis. 1998, 4, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Anil, S.; Ellepola, A.N.; Samaranayake, L.P. Post-antifungal effect of polyene, azole and DNA-analogue agents against oral Candida albicans and Candida tropicalis isolates in HIV disease. J. Oral Pathol. Med. 2001, 30, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Ellepola, A.N.; Samaranayake, L.P. Effects of two different growth media on the postantifungal effect induced by polyenes on Candida species. J. Clin. Microbiol. 2001, 39, 2732–2735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkutuk, A.; Dogan, Y.; Kirdar, S.; Ozkutuk, N. Changes in some Candida albicans virulence factors due to amphotericin-B post-antifungal effect. J. Chemother. 2007, 19, 101–103. [Google Scholar] [CrossRef]
- Ellepola, A.N. Amphotericin B-induced in vitro postantifungal effect on Candida species of oral origin. Med. Princ. Pract. 2012, 21, 442–446. [Google Scholar] [CrossRef]
- Ellepola, A.N.; Joseph, B.K.; Chandy, R.; Khan, Z.U. The postantifungal effect of nystatin and its impact on adhesion attributes of oral Candida dubliniensis isolates. Mycoses 2014, 57, 56–63. [Google Scholar] [CrossRef]
- Ellepola, A.N.; Chandy, R.; Khan, Z.U. In vitro postantifungal effect, adhesion traits and haemolysin production of Candida dubliniensis isolates following exposure to 5-fluorocytosine. Mycoses 2015, 58, 536–543. [Google Scholar] [CrossRef]
- Ellepola, A.N.; Chandy, R.; Khan, Z.U. Post-antifungal effect and adhesion to buccal epithelial cells of oral Candida dubliniensis isolates subsequent to limited exposure to amphotericin B, ketoconazole and fluconazole. J. Investig. Clin. Dent. 2015, 6, 186–192. [Google Scholar] [CrossRef]
- Ellepola, A.N.; Chandy, R.; Khan, Z.U.; Samaranayake, L.P. Caspofungin-induced in-vitro post-antifungal effect and its impact on adhesion related traits of oral Candida dubliniensis and Candida albicans isolates. Microbiol. Immunol. 2016, 60, 160–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellepola, A.N.B.; Dassanayake, R.S.; Khan, Z. In vitro Post-Antifungal Effect of Posaconazole and Its Impact on Adhesion-Related Traits and Hemolysin Production of Oral Candida dubliniensis isolates. Med. Princ. Pract. 2019, 28, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E.J.; Klepser, M.E.; Pfaller, M.A. Postantifungal effects of echinocandin, azole, and polyene antifungal agents against Candida albicans and Cryptococcus neoformans. Antimicrob. Agents Chemother. 2000, 44, 1108–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhanel, G.G.; Saunders, D.G.; Hoban, D.J.; Karlowsky, J.A. Influence of human serum on antifungal pharmacodynamics with Candida albicans. Antimicrob. Agents Chemother. 2001, 45, 2018–2022. [Google Scholar] [CrossRef] [Green Version]
- García, M.T.; Llorente, M.T.; Mínguez, F.; Prieto, J. Postantifungal effect and effects of sub-MIC concentrations on previously treated Candida sp. Influence of growth phase. J. Infect. 2002, 45, 263–267. [Google Scholar] [CrossRef]
- García, M.T.; Llorente, M.T.; Mínguez, F.; Prieto, J. Post-antifungal effect and effects of sub-MIC concentrations on previously treated Candida spp.: Influence of exposure time and concentration. Scand. J. Infect Dis. 2002, 34, 197–200. [Google Scholar] [CrossRef]
- Di Bonaventura, G.; Spedicato, I.; Picciani, C.; D’Antonio, D.; Piccolomini, R. In vitro pharmacodynamic characteristics of amphotericin B, caspofungin, fluconazole, and voriconazole against bloodstream isolates of infrequent Candida species from patients with hematologic malignancies. Antimicrob. Agents Chemother. 2004, 48, 4453–4456. [Google Scholar] [CrossRef] [Green Version]
- Manavathu, E.K.; Ramesh, M.S.; Baskaran, I.; Ganesan, L.T.; Chandrasekar, P.H. A comparative study of the post-antifungal effect (PAFE) of amphotericin B, triazoles and echinocandins on Aspergillus fumigatus and Candida albicans. J. Antimicrob. Chemother. 2004, 53, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Clancy, C.J.; Huang, H.; Cheng, S.; Derendorf, H.; Nguyen, M.H. Characterizing the effects of caspofungin on Candida albicans, Candida parapsilosis, and Candida glabrata isolates by simultaneous time-kill and postantifungal-effect experiments. Antimicrob. Agents Chemother. 2006, 50, 2569–2572. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Nguyen, M.H.; Derendorf, H.; Cheng, S.; Clancy, C.J. Measurement of voriconazole activity against Candida albicans, C. glabrata, and C. parapsilosis isolates using time-kill methods validated by high-performance liquid chromatography. Antimicrob. Agents Chemother. 2007, 51, 2985–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.T.; Ta, P.; Hoang, B.T.; Cheng, S.; Hao, B.; Nguyen, M.H.; Clancy, C.J. Anidulafungin is fungicidal and exerts a variety of postantifungal effects against Candida albicans, C. glabrata, C. parapsilosis, and C. krusei isolates. Antimicrob. Agents Chemother. 2009, 53, 3347–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.T.; Ta, P.; Hoang, B.T.; Cheng, S.; Hao, B.; Nguyen, M.H.; Clancy, C.J. Characterising the post-antifungal effects of micafungin against Candida albicans, Candida glabrata, Candida parapsilosis and Candida krusei isolates. Int. J. Antimicrob. Agents 2010, 35, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Clancy, C.J. Five-minute exposure to caspofungin results in prolonged postantifungal effects and eliminates the paradoxical growth of Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 3598–3602. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.P.; Baltch, A.; Bopp, L.H.; Ritz, W.J.; Michelsen, P.P. Post-antifungal effects and time-kill studies of anidulafungin, caspofungin, and micafungin against Candida glabrata and Candida parapsilosis. Diagn. Microbiol. Infect Dis. 2011, 71, 131–138. [Google Scholar] [CrossRef]
- Moriyama, B.; Henning, S.A.; Penzak, S.R.; Walsh, T.J. The postantifungal and paradoxical effects of echinocandins against Candida spp. Future Microbiol. 2012, 7, 565–569. [Google Scholar] [CrossRef]
- Kovacs, R.; Gesztelyi, R.; Perlin, D.S.; Kardos, G.; Doman, M.; Berenyi, R.; Majoros, L. Killing rates for caspofungin against Candida albicans after brief and continuous caspofungin exposure in the presence and absence of serum. Mycopathologia 2014, 178, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Gil-Alonso, S.; Jauregizar, N.; Eraso, E.; Quindós, G. Postantifungal effect of micafungin against the species complexes of Candida albicans and Candida parapsilosis. PLoS ONE 2015, 10, e0132730. [Google Scholar] [CrossRef]
- Gil-Alonso, S.; Jauregizar, N.; Eraso, E.; Quindós, G. Postantifungal effect of caspofungin against the Candida albicans and Candida parapsilosis clades. Diagn. Microbiol. Infect. Dis. 2016, 86, 172–177. [Google Scholar] [CrossRef]
- Kardos, T.; Saleh, Q.; Kovacs, R.; Gesztelyi, R.; Kardos, G.; Bozo, A.; Majoros, L. Postantifungal effect of micafungin against Candida albicans, Candida dubliniensis and Candida Africana in the presence and absence of serum. New Microbiol. 2017, 40, 286–288. [Google Scholar]
- Gil-Alonso, S.; Quindós, G.; Eraso, E.; Jauregizar, N. Postantifungal effect of anidulafungin against Candida albicans, Candida dubliniensis, Candida africana, Candida parapsilosis, Candida metapsilosis and Candida orthopsilosis. Rev. Esp. Quimioter. 2019, 32, 183–188. [Google Scholar] [PubMed]
- Gowri, M.; Jayashree, B.; Jeyakanthan, J.; Girija, E.K. Sertraline as a promising antifungal agent: Inhibition of growth and biofilm of Candida auris with special focus on the mechanism of action in vitro. J. Appl. Microbiol. 2020, 128, 426–437. [Google Scholar] [CrossRef] [PubMed]
- García, M.T.; Llorente, M.T.; Mínguez, F.; Prieto, J. Influence of temperature and concentration on the postantifungal effect and the effects of sub-MIC concentrations of four antifungal agents on previously treated Candida species. Chemotherapy 2000, 46, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Scalarone, G.M.; Mikami, Y.; Kurita, N.; Yazawa, K.; Uno, J.; Miyaji, M. In vitro comparative evaluations of the postantifungal effect: Synergistic interaction between flucytosine and fluconazole against Candida albicans. Mycoses 1991, 34, 405–410. [Google Scholar] [CrossRef]
- Mikami, Y.; Scalarone, G.M.; Kurita, N.; Yazawa, K.; Uno, J.; Miyaji, M. Synergistic postantifungal effect of flucytosine and fluconazole on Candida albicans. J. Med. Vet. Mycol. 1992, 30, 197–206. [Google Scholar] [CrossRef]
- Scalarone, G.M.; Mikami, Y.; Kurita, N.; Yazawa, K.; Miyaji, M. Comparative studies on the postantifungal effect produced by the synergistic interaction of flucytosine and amphotericin B on Candida albicans. Mycopathologia 1992, 120, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.; Stepensky, D. Pharmacodynamic aspects of modes of drug administration for optimization of drug therapy. Crit. Rev. Ther. Drug Carrier Syst. 1999, 16, 571–639. [Google Scholar] [CrossRef]
- Venisse, N.; Gregoire, N.; Marliat, M.; Couet, W. Mechanism-based pharmacokinetic-pharmacodynamic models of in vitro fungistatic and fungicidal effects against Candida albicans. Antimicrob. Agents Chemother. 2008, 52, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Foldi, R.; Kovacs, R.; Gesztelyi, R.; Kardos, G.; Berenyi, R.; Juhasz, B.; Szilágyi, J.; Mózes, J.; Majoros, L. Comparison of in vitro and vivo efficacy of caspofungin against Candida parapsilosis, C. orthopsilosis, C. metapsilosis and C. albicans. Mycopathologia 2012, 174, 311–318. [Google Scholar] [CrossRef]
- Spreghini, E.; Orlando, F.; Tavanti, A.; Senesi, S.; Giannini, D.; Manso, E.; Barchiesi, F. In vitro and in vivo effects of echinocandins against Candida parapsilosis sensu stricto, Candida orthopsilosis and Candida metapsilosis. J. Antimicrob. Chemother. 2012, 67, 2195–2202. [Google Scholar] [CrossRef] [Green Version]
- Tóth, Z.; Kardos, T.; Kovács, R.; Kardos, G.; Nagy, F.; Prépost, E.; Barta, Z.; Takacs, I.; Majoros, L. comparison of killing activity of micafungin against six candida species isolated from peritoneal and pleural cavities in RPMI-1640, 10 and 30% serum. Mycopathologia 2018, 183, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Tóth, Z.; Locke, J.B.; Forgács, L.; Kardos, G.; Nagy, F.; Borman, A.; Majoros, L. comparison of in vitro killing activity of rezafungin, anidulafungin, caspofungin, and micafungin against four Candida auris clades in RPMI-1640 in the absence and presence of human serum. Microorganisms 2021, 9, 863. [Google Scholar] [CrossRef] [PubMed]
- Mínguez, F.; García, M.T.; Lima, J.E.; Llorente, M.T.; Prieto, J. Postantifungal effect and effects of low concentrations of amphotericin B and fluconazole on previously treated Candida albicans. Scand. J. Infect. Dis. 1996, 28, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Mínguez, F.; Lima, J.E.; García, M.T.; Prieto, J. Influence of human serum on the postantifungal effect of four antifungal agents on Candida albicans. Chemotherapy 1996, 42, 273–279. [Google Scholar] [CrossRef]
- Jota Baptista, C.V.; Faustino-Rocha, A.I.; Oliveira, P.A. Animal models in pharmacology: A brief history awarding the nobel prizes for physiology or medicine. Pharmacology 2021, 106, 356–368. [Google Scholar] [CrossRef]
- Andes, D.; Marchillo, K.; Stamstad, T.; Conklin, R. In vivo pharmacodynamics of a new triazole, ravuconazole, in a murine candidiasis model. Antimicrob. Agents Chemother. 2003, 47, 1193–1199. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.; Marchillo, K.; Lowther, J.; Bryskier, A.; Stamstad, T.; Conklin, R. In vivo pharmacodynamics of HMR 3270, a glucan synthase inhibitor, in a murine candidiasis model. Antimicrob. Agents Chemother. 2003, 47, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.; Marchillo, K.; Conklin, R.; Krishna, G.; Ezzet, F.; Cacciapuoti, A.; Loebenberg, D. Pharmacodynamics of a new triazole, posaconazole, in a murine model of disseminated candidiasis. Antimicrob. Agents Chemother. 2004, 48, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.; Diekema, D.J.; Pfaller, M.A.; Prince, R.A.; Marchillo, K.; Ashbeck, J.; Hou, J. In vivo pharmacodynamic characterization of anidulafungin in a neutropenic murine candidiasis model. Antimicrob. Agents Chemother. 2008, 52, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Warn, P.A.; Sharp, A.; Parmar, A.; Majithiya, J.; Denning, D.W.; Hope, W.W. Pharmacokinetics and pharmacodynamics of a novel triazole, isavuconazole: Mathematical modeling, importance of tissue concentrations, and impact of immune status on antifungal effect. Antimicrob. Agents Chemother. 2009, 53, 3453–3461. [Google Scholar] [CrossRef] [Green Version]
- Maki, K.; Kaneko, S. Effective concentration-based serum pharmacodynamics for antifungal azoles in a murine model of disseminated Candida albicans infection. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavassin, F.B.; Baú-Carneiro, J.L.; Vilas-Boas, R.R.; Queiroz-Telles, F. Sixty years of amphotericin B: An overview of the main antifungal agent used to treat invasive fungal infections. Infect. Dis. Ther. 2021, 10, 115–147. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D. Antifungals in clinical use and the pipeline. Infect. Dis. Clin. North Am. 2021, 35, 341–371. [Google Scholar] [CrossRef] [PubMed]
- Turnidge, J.D.; Gudmundsson, S.; Vogelman, B.; Craig, W.A. The postantibiotic effect of antifungal agents against common pathogenic yeasts. J. Antimicrob. Chemother. 1994, 34, 83–92. [Google Scholar] [CrossRef]
- Egusa, H.; Ellepola, A.N.; Nikawa, H.; Hamada, T.; Samaranayake, L.P. Sub-therapeutic exposure to polyene antimycotics elicits a post-antifungal effect (PAFE) and depresses the cell surface hydrophobicity of oral Candida albicans isolates. J. Oral Pathol. Med. 2000, 29, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Ellepola, A.N.; Khajah, R.; Jayatilake, S.; Samaranayake, L.; Sharma, P.; Khan, Z. Impact of brief exposure to antifungal agents on the post-antifungal effect and hemolysin activity of oral Candida albicans. J. Appl. Oral Sci. 2015, 23, 412–418. [Google Scholar] [CrossRef]
- Ellepola, A.N.; Samaranayake, L.P. The in vitro post-antifungal effect of nystatin on Candida species of oral origin. J. Oral Pathol. Med. 1999, 28, 112–116. [Google Scholar] [CrossRef]
- Samaranayake, Y.H.; Yau, J.Y.; Thein, Z.M.; Jayatilake, J.A.; Yeung, K.W.; Samaranayake, L.P. The post-antifungal effect (PAFE) of amphotericin B, nystatin, ketoconazole and 5-fluorocytosine and its impact on the colonization traits of Candida glabrata. Med. Mycol. 2010, 48, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.; van Ogtrop, M. Characterization and quantitation of the pharmacodynamics of fluconazole in a neutropenic murine disseminated candidiasis infection model. Antimicrob. Agents Chemother. 1999, 43, 2116–2120. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrial.gov Rezafungin. Available online: https://clinicaltrials.gov/ct2/results?cond=rezafungin&term=&cntry=&state=&city=&dist= (accessed on 16 May 2022).
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice guideline for the management of candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, 1. [Google Scholar] [CrossRef]
- Ernst, E.J.; Roling, E.E.; Petzold, C.R.; Keele, D.J.; Klepser, M.E. In vitro activity of micafungin (FK-463) against Candida spp.: Microdilution, time-kill, and postantifungal-effect studies. Antimicrob. Agents Chemother. 2002, 46, 3846–3853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepak, A.J.; Zhao, M.; Andes, D.R. Pharmacodynamic Evaluation of rezafungin (CD101) against Candida auris in the neutropenic mouse invasive candidiasis model. Antimicrob Agents Chemother. 2018, 62, e01572-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miesel, L.; Lin, K.Y.; Ong, V. Rezafungin treatment in mouse models of invasive candidiasis and aspergillosis: Insights on the PK/PD pharmacometrics of rezafungin efficacy. Pharmacol. Res. Perspect. 2019, 7, e00546. [Google Scholar] [CrossRef] [PubMed]
- Andes, D.; van Ogtrop, M. In vivo characterization of the pharmacodynamics of flucytosine in a neutropenic murine disseminated candidiasis model. Antimicrob. Agents Chemother. 2000, 44, 938–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louie, A.; Deziel, M.; Liu, W.; Drusano, M.F.; Gumbo, T.; Drusano, G.L. Pharmacodynamics of caspofungin in a murine model of systemic candidiasis: Importance of persistence of caspofungin in tissues to understanding drug activity. Antimicrob. Agents Chemother. 2005, 12, 5058–5068. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Khan, A.; Khan, S.H.; Azam, M.; Khan, M.M.U.; Khalilullah, H.; Younus, H. Coadministration of liposomal methylglyoxal increases the activity of amphotericin B against Candida albicans in leukopoenic mice. J. Drug Target. 2021, 29, 78–87. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Rueda, C.; Román, E.; Quintin, J.; Terrón, M.C.; Luque, D.; Netea, M.G.; Pla, J.; Zaragoza, O. Cell wall changes in amphotericin B-resistant strains from Candida tropicalis and relationship with the immune responses elicited by the host. Antimicrob. Agents Chemother. 2016, 60, 2326–2335. [Google Scholar] [CrossRef] [Green Version]
- Asín-Prieto, E.; Rodríguez-Gascón, A.; Isla, A. Applications of the pharmacokinetic/pharmacodynamic (PK/PD) analysis of antimicrobial agents. J. Infect. Chemother. 2015, 21, 319–329. [Google Scholar] [CrossRef]
- Sy, S.K.; Zhuang, L.; Derendorf, H. Pharmacokinetics and pharmacodynamics in antibiotic dose optimization. Expert Opin. Drug Metab. Toxicol. 2016, 12, 93–114. [Google Scholar] [CrossRef]
- Lewis, R.E.; Andes, D.R. Managing uncertainty in antifungal dosing: Antibiograms, therapeutic drug monitoring and drug-drug interactions. Curr. Opin. Infect. Dis. 2021, 34, 288–296. [Google Scholar] [CrossRef]
- McAleenan, A.; Ambrose, P.G.; Bhavnani, S.M.; Drusano, G.L.; Hope, W.W.; Mouton, J.W.; Higgins, J.P.T.; MacGowan, A.P. Methodological features of clinical pharmacokinetic-pharmacodynamic studies of antibacterials and antifungals: A systematic review. J. Antimicrob. Chemother. 2020, 75, 1374–1389. [Google Scholar] [CrossRef]
- Arredondo, G.; Suárez, E.; Calvo, R.; Vazquez, J.A.; García-Sanchez, J.; Martinez-Jordá, R. Serum protein binding of itraconazole and fluconazole in patients with diabetes mellitus. J. Antimicrob. Chemother. 1999, 43, 305–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florent, A.; Gandia, P.; Seraissol, P.; Chatelut, E.; Houin, G. Determination of plasma unbound fraction of voriconazole in patients treated with a prophylactic or a curative treatment. Ther. Drug Monit. 2014, 36, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Sakurama, K.; Nishi, K.; Tsukigawa, K.; Seo, H.; Otagiri, M.; Taguchi, K. An in-vitro comparative study of the binding of caspofungin and micafungin to plasma proteins. J. Pharm. Pharmacol. 2022, 74, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Andes, D. Optimizing antifungal choice and administration. Curr. Med. Res. Opin. 2013, 29 (Suppl. 4), 13–18. [Google Scholar] [CrossRef] [Green Version]
- Prépost, E.; Tóth, Z.; Perlin, D.S.; Gesztelyi, R.; Kardos, G.; Kovács, R.; Nagy, F.; Forgács, L.; Majoros, L. Efficacy of humanized single large doses of caspofungin on the lethality and fungal tissue burden in a deeply neutropenic murine model against Candida albicans and Candida dubliniensis. Infect. Drug Resist. 2019, 12, 1805–1814. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Prideaux, B.; Nagasaki, Y.; Lee, M.H.; Chen, P.Y.; Blanc, L.; Ho, H.; Clancy, C.J.; Nguyen, M.H.; Dartois, V.; et al. Unraveling drug penetration of echinocandin antifungals at the site of infection in an intra-abdominal abscess model. Antimicrob. Agents Chemother. 2017, 61, e01009-17. [Google Scholar] [CrossRef] [Green Version]
- CANCIDAS, INN-Caspofungin (as Acetate)–Europa EU. Available online: https://www.ema.europa.eu/en/documents/product-information/cancidas-epar-product-information_en.pdf (accessed on 18 May 2022).
- Cornely, O.A.; Vehreschild, J.J.; Vehreschild, M.J.; Würthwein, G.; Arenz, D.; Schwartz, S.; Heussel, C.P.; Silling, G.; Mahne, M.; Franklin, J.; et al. Phase II dose escalation study of caspofungin for invasive aspergillosis. Antimicrob. Agents Chemother. 2011, 55, 5798–5803. [Google Scholar] [CrossRef] [Green Version]
- Chatelon, J.; Cortegiani, A.; Hammad, E.; Cassir, N.; Leone, M. Choosing the right antifungal agent in ICU patients. Adv. Ther. 2019, 36, 3308–3320. [Google Scholar] [CrossRef] [Green Version]
- Hope, W.W.; Kaibara, A.; Roy, M.; Arrieta, A.; Azie, N.; Kovanda, L.L.; Benjamin, D.K., Jr. Population pharmacokinetics of micafungin and its metabolites M1 and M5 in children and adolescents. Antimicrob. Agents Chemother. 2015, 59, 905–913. [Google Scholar] [CrossRef] [Green Version]
- Yanni, S.B.; Smith, P.B.; Benjamin, D.K., Jr.; Augustijns, P.F.; Thakker, D.R.; Annaert, P.P. Higher clearance of micafungin in neonates compared with adults: Role of age-dependent micafungin serum binding. Biopharm. Drug Dispos. 2011, 32, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ittel, T.H.; Legler, U.F.; Polak, A.; Glöckner, W.M.; Sieberth, H.G. 5-Fluorocytosine kinetics in patients with acute renal failure undergoing continuous hemofiltration. Chemotherapy 1987, 33, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Weiler, S.; Seger, C.; Pfisterer, H.; Stienecke, E.; Stippler, F.; Welte, R.; Joannidis, M.; Griesmacher, A.; Bellmann, R. Pharmacokinetics of caspofungin in critically ill patients on continuous renal replacement therapy. Antimicrob. Agents Chemother. 2013, 57, 4053–4057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Undre, N.; Pretorius, B.; Stevenson, P. Pharmacokinetics of micafungin in subjects with severe hepatic dysfunction. Eur. J. Drug Metab. Pharmacokinet. 2015, 40, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.; Schmitt-Hoffmann, A.H.; Mujais, S.; Townsend, R. Population pharmacokinetics of isavuconazole in subjects with mild or moderate hepatic impairment. Antimicrob. Agents Chemother. 2016, 60, 3025–3031. [Google Scholar] [CrossRef] [Green Version]
- Oshima, K.; Kanda, Y.; Kako, S.; Ohno, K.; Kishino, S.; Kurokawa, M. Pharmacokinetics of micafungin in patients undergoing allogeneic hematopoietic stem cell transplantation. Transpl. Infect. Dis. 2013, 15, 323–327. [Google Scholar] [CrossRef]
- Würthwein, G.; Young, C.; Lanvers-Kaminsky, C.; Hempel, G.; Trame, M.N.; Schwerdtfeger, R.; Ostermann, H.; Heinz, W.J.; Cornely, O.A.; Kolve, H.; et al. Population pharmacokinetics of liposomal amphotericin B and caspofungin in allogeneic hematopoietic stem cell recipients. Antimicrob. Agents Chemother. 2012, 56, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.; Campbell, L.; Rudham, S.; Nguyen, Q.; Marriott, D. Posaconazole plasma concentrations in critically ill patients. Ther. Drug Monit. 2011, 33, 387–392. [Google Scholar] [CrossRef]
- Maseda, E.; Grau, S.; Hernandez-Gancedo, C.; Suarez-de-la-Rica, A.; Aguilar, L.; Gilsanz, F. Pharmacokinetics/pharmacodynamics of micafungin in a surgical critically ill patient during extracorporeal carbon dioxide removal and continuous renal replacement therapy. J. Crit. Care 2015, 30, 1129–1130. [Google Scholar] [CrossRef]
- Sinnollareddy, M.G.; Roberts, J.A.; Lipman, J.; Akova, M.; Bassetti, M.; De Waele, J.J.; Kaukonen, K.M.; Koulenti, D.; Martin, C.; Montravers, P.; et al. Pharmacokinetic variability and exposures of fluconazole, anidulafungin, and caspofungin in intensive care unit patients: Data from multinational Defining Antibiotic Levels in Intensive care unit (DALI) patients Study. Crit. Care 2015, 19, 33. [Google Scholar] [CrossRef] [Green Version]
- Roger, C.; Wallis, S.C.; Muller, L.; Saissi, G.; Lipman, J.; Brüggemann, R.J.; Lefrant, J.Y.; Roberts, J.A. Caspofungin population pharmacokinetics in critically ill patients undergoing continuous veno-venous haemofiltration or haemodiafiltration. Clin. Pharmacokinet. 2017, 56, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Bellmann, R.; Smuszkiewicz, P. Pharmacokinetics of antifungal drugs: Practical implications for optimized treatment of patients. Infection 2017, 45, 737–779. [Google Scholar] [CrossRef] [PubMed]
- Andes, D.; Azie, N.; Yang, H.; Harrington, R.; Kelley, C.; Tan, R.D.; Wu, E.Q.; Franks, B.; Kristy, R.; Lee, E.; et al. Drug-drug interaction associated with mold-active triazoles among hospitalized patients. Antimicrob. Agents Chemother. 2016, 60, 3398–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing. Antimicrobial Wild Type Distribution of Microorganism. Available online: https://mic.eucast.org/Eucast2/ (accessed on 15 May 2022).
- Heffernan, A.J.; Sime, F.B.; Lipman, J.; Roberts, J.A. Individualising therapy to minimize bacterial multidrug resistance. Drugs 2018, 78, 621–641. [Google Scholar] [CrossRef] [PubMed]
- Srimani, J.K.; Huang, S.; LopaCCin, A.J.; You, L. Drug detoxification dynamics explain the postantibiotic effect. Mol. Syst. Biol. 2017, 13, 948. [Google Scholar] [CrossRef] [PubMed]
- Andes, D.; Ambrose, P.G.; Hammel, J.P.; Van Wart, S.A.; Iyer, V.; Reynolds, D.K.; Buell, D.N.; Kovanda, L.L.; Bhavnani, S.M. Use of pharmacokinetic-pharmacodynamic analyses to optimize therapy with the systemic antifungal micafungin for invasive candidiasis or candidemia. Antimicrob. Agents Chemother. 2011, 55, 2113–2121. [Google Scholar] [CrossRef] [Green Version]
- Tascini, C.; Sozio, E.; Di Paolo, A.; Tintori, G.; Leonildi, A.; Bertolino, G.; Carmassi, F.; Tagliaferri, E.; Menichetti, F.; Barchiesi, F. Fungicidal activity and PK/PD of caspofungin as tools to guide antifungal therapy in a fluconazole-resistant C. parapsilosis candidemia. J. Chemother. 2017, 29, 376–379. [Google Scholar] [CrossRef] [Green Version]
- Pea, F. From bench to bedside: Perspectives on the utility of pharmacokinetics/pharmacodynamics in predicting the efficacy of antifungals in invasive candidiasis. Mycoses 2020, 63, 854–858. [Google Scholar] [CrossRef]
- Lepak, A.J.; Andes, D.R. Antifungal pharmacokinetics and pharmacodynamics. Cold Spring Harb Perspect. Med. 2014, 5, a019653. [Google Scholar] [CrossRef]
- Perez-Pitarch, A.; Guglieri-Lopez, B.; Ferriols-Lisart, R.; Pérez, A.; Ezquer-Garín, C.; Hernández-Boluda, J.C.; Piñana, J.L.; Navarro, D.; Solano, C.; Alós-Almiñana, M. Pharmacokinetic/Pharmacodynamic analysis of voriconazole against Candida spp. and Aspergillus spp. in allogeneic stem cell transplant recipients. Ther. Drug Monit. 2019, 41, 740–747. [Google Scholar] [CrossRef]
- Andes, D.; Diekema, D.J.; Pfaller, M.A.; Bohrmuller, J.; Marchillo, K.; Lepak, A. In vivo comparison of the pharmacodynamic targets for echinocandin drugs against Candida species. Antimicrob. Agents Chemother. 2010, 54, 2497–2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, J.C.; Bhavnani, S.M.; Andes, D.R.; Ambrose, P.G. We can do better: A fresh look at echinocandin dosing. J. Antimicrob. Chemother. 2018, 73, 831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Velde, F.; Mouton, J.W.; de Winter, B.C.M.; van Gelder, T.; Koch, B.C.P. Clinical applications of population pharmacokinetic models of antibiotics: Challenges and perspectives. Pharmacol. Res. 2018, 134, 280–288. [Google Scholar] [CrossRef]
- Trang, M.; Dudley, M.N.; Bhavnani, S.M. Use of Monte Carlo simulation and considerations for PK-PD targets to support antibacterial dose selection. Curr. Opin. Pharmacol. 2017, 36, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Rayner, C.R.; Smith, P.F.; Andes, D.; Andrews, K.; Derendorf, H.; Friberg, L.E.; Hanna, D.; Lepak, A.; Mills, E.; Polasek, T.M.; et al. Model-informed drug development for anti-infectives: State of the art and future. Clin. Pharmacol. Ther. 2021, 109, 867–891. [Google Scholar] [CrossRef]
- Gómez-López, A. Antifungal therapeutic drug monitoring: Focus on drugs without a clear recommendation. Clin. Microbiol. Infect. 2020, 26, 1481–1487. [Google Scholar] [CrossRef]
- McCarthy, M.W. Pharmacokinetics and Pharmacodynamics of Ibrexafungerp. Drugs R D 2022, 22, 9–13. [Google Scholar] [CrossRef]
- Quindós, G.; Miranda-Cadena, K.; San-Millán, R.; Borroto-Esoda, K.; Cantón, E.; Linares-Sicilia, M.J.; Hamprecht, A.; Montesinos, I.; Tortorano, A.M.; Prigitano, A.; et al. In Vitro Antifungal Activity of Ibrexafungerp (SCY-078) Against Contemporary Blood Isolates From Medically Relevant Species of Candida: A European Study. Front Cell Infect. Microbiol. 2022, 16, 906563. [Google Scholar] [CrossRef]
- Davis, M.R.; Donnelley, M.A.; Thompson, G.R. Ibrexafungerp: A novel oral glucan synthase inhibitor. Med. Mycol. 2020, 1, 579–592. [Google Scholar] [CrossRef]
- Chryssanthou, E.; Loebig, A.; Sjölin, J. Post-antifungal effect of amphotericin B and voriconazole against germinated Aspergillus fumigatus conidia. J. Antimicrob. Chemother. 2008, 61, 1309–1311. [Google Scholar] [CrossRef]
- D’Agostino, M.; Tesse, N.; Lavergne, R.; Pape, P.L.; Bouchara, J.P.; Frippiat, J.P.; Machouart, M.; Debourgogne, A. Characterisation of the antifungal effects of a plant-based compound, CIN-102, on the main septal filamentous fungi involved in human pathology. J. Glob. Antimicrob. Resist. 2021, 25, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Vitale, R.; Meis, J.F.G.M.; Mouton, J.W.; Verweij, P.E. Evaluation of the post-antifungal effect (PAFE) of amphotericin B and nystatin against 30 zygomycetes using two different media. J. Antimicrob. Chemother. 2003, 52, 65–70. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antifungal | Species (Strains) | MIC (mg/L) | Exposure | Concentration | PAFE (h) | Methodology * | Reference |
|---|---|---|---|---|---|---|---|
| Amphotericin B | C. albicans (2) | 0.25–1 | 12 h | 0.5, 1 mg/L | 0.9–2.5, 2.4–4.1 | CC, bYNBg | [44] |
| Amphotericin B | C. albicans (12) | 0.19 | 1 h | 2 × MIC | 4.69–13.44 | OD, RPMI | [56] |
| Amphotericin B | C. albicans (2) | 0.5–1 | 1 h | 0.125–4 × MIC | 2–>12 | CC, RPMI | [15] |
| Amphotericin B | C. albicans (10) | 0.19–0.38 | 1 h | 2 × MIC | 9.93 | OD, RPMI | [6] |
| Amphotericin B | C. albicans (1) | 0.5 | 0.25–0.5 h | 1, 2.5, 5, 10, 20 × MIC | 0.96–4.04, 2.54–7.68, 3.92–9.13, 7.67–11.54, 10.67 | CO2, RPMI | [4] |
| Amphotericin B | C. albicans (1) | 1 | 1.5–12 h | 1, 4, 8 × MIC | 0.8–4.9, 3.5–8.0, 4.6–12 | CC, bYNBg | [18] |
| Amphotericin B | C. albicans (1) | 0.125 | 1 h | 1–16 mg/L | 5.3 | CC, RPMI | [20] |
| Amphotericin B | C. albicans (1) | 0.125–1 | 1.5 h | 1, 2, 4, 8 × MIC | 0.54–16.35, 3.48–19.31, 9.11–19.86, 16.25–21.54 | OD, RPMI | [8] |
| Amphotericin B | C. albicans (50) | 0.004–0.19 | 1 h | 2 × MIC | 2.18 | OD, RPMI | [57] |
| Amphotericin B | C. dubliniensis (20) | 0.002–0.125 | 1 h | 3 × MIC | 1.92–2.41 | OD, RPMI | [12] |
| Amphotericin B | C. glabrata (1) | 0.5 | 0.25 h | 5, 20 × MIC | 3.19, 5.02 | CO2, RPMI | [4] |
| Amphotericin B | C. glabrata (1) | 0.5 | 0.5 h | 2.5, 10 × MIC | 4.18, 6.65 | CO2, RPMI | [4] |
| Amphotericin B | C. glabrata | 0.5 | 1.5–12 h | 1, 4, 8 × MIC | 1.3–5.3, 3.5–8.6, 4.8–13 | CC, bYNBg | [18] |
| Amphotericin B | C. glabrata (14) | 0.256–0.512 | 1 h | 1, 2, 4 xMIC | 5.92–10.50, 6.42–18.50, 8.67–22.00 | OD, RPMI | [59] |
| Amphotericin B | C. guilliermondii (2), C. kefyr (2), C. lusitaniae (2) | 0.5–4 | 1 h | 0.125, 0.25, 1 × MIC | 1.3–9.4, 3.6–10, 9.2–14.9 | CC, RPMI | [19] |
| Amphotericin B | C. krusei (1) | 0.5 | 0.25 h | 5, 20 × MIC | 3.33, 9.65 | CO2, RPMI | [4] |
| Amphotericin B | C. krusei (1) | 0.5 | 0.5 h | 2.5, 10 × MIC | 5.27, 14.24 | CO2, RPMI | [4] |
| Amphotericin B | C. tropicalis (10) | 0.25–0.38 | 1 h | 2 × MIC | 12.42 | OD, RPMI | [6] |
| Nystatin | C. albicans (5) | 0.78–1.56 | 1 h | 1 × MIC | 6.85 | OD, RPMI | [58] |
| Nystatin | C. albicans (12) | 0.78–1.56 | 1 h | 2 × MIC | 1.91–7.99 | OD, RPMI | [56] |
| Nystatin | C. albicans (10) | 0.78–1.56 | 1 h | 2 × MIC | 12.31 | OD, RPMI | [6] |
| Nystatin | C. albicans (50) | 0.78–1.56 | 1 h | 2 × MIC | 2.20 | OD, RPMI | [57] |
| Nystatin | C. dubliniensis (20) | 0.09–0.78 | 1 h | 3 × MIC | 1.92–2.41 | OD, RPMI | [10] |
| Nystatin | C. glabrata (5) | 0.78–1.56 | 1 h | 1 × MIC | 8.51 | OD, RPMI | [58] |
| Nystatin | C. glabrata (14) | 0.39–1.56 | 1 h | 1, 2, 4 × MIC | 0–9.36, 3.60–17.83, 0–13.57 | OD, RPMI | [59] |
| Nystatin | C. guilliermondii (5) | 0.39–0.78 | 1 h | 1 × MIC | 8.68 | OD, RPMI | [58] |
| Nystatin | C. krusei (5) | 3.12 | 1 h | 1 × MIC | 11.58 | OD, RPMI | [58] |
| Nystatin | C. parapsilosis (5) | 1.56–3.12 | 1 h | 1 × MIC | 15.17 | OD, RPMI | [58] |
| Nystatin | C. tropicalis (5) | 1.56–3.12 | 1 h | 1 × MIC | 12.73 | OD, RPMI | [58] |
| Nystatin | C. tropicalis (10) | 0.78 | 1 h | 2 × MIC | 14.83 | OD, RPMI | [6] |
| Antifungal | Species (Strains) | MIC (mg/L) | Exposure | Concentration | PAFE (h) | Methodology * | Reference |
|---|---|---|---|---|---|---|---|
| Fluconazole | C. albicans (2) | 2–4 | 12 h | 4, 8 mg/L | No measurable | CC, bYNBg | [44] |
| Fluconazole | C. albicans (2) | 0.25 | 1 h | 1, 2, 4 × MIC | No measurable | CC, RPMI | [15] |
| Fluconazole | C. albicans (10) | 0.125–0.38 | 1 h | 2 × MIC | No measurable | OD, RPMI | [6] |
| Fluconazole | C. albicans (1) | 4 | 1.5–12 h | 1, 4, 8 × MIC | No measurable | CC, bYNBg | [18] |
| Fluconazole | C. albicans (50) | 0.047–0.125 | 1 h | 2 × MIC | No measurable | OD, RPMI | [57] |
| Fluconazole | C. dubliniensis (20) | 0.016–0.38 | 1 h | 3 × MIC | No measurable | OD, RPMI | [12] |
| Fluconazole | C. guilliermondii (2), C. kefyr (2), C. lusitaniae (2) | 0.12–2 | 1 h | 0.125–8 × MIC | No measurable | CC, RPMI | [19] |
| Fluconazole | C. tropicalis (10) | 0.25–0.50 | 1 h | 2 × MIC | No measurable | OD, RPMI | [6] |
| Itraconazole, Voriconazole, Posaconazole, Ravuconazole | C. albicans (1) | 0.06–0.25 | 1 h | 1–16 mg/L | ≤0.5 | CC, RPMI | [20] |
| Ketoconazole | C. albicans (10) | 0.012–0.016 | 1 h | 2 × MIC | 1.14 | OD, RPMI | [6] |
| Ketoconazole | C. albicans (1) | 1 | 1.5–12 h | 1, 4, 8 × MIC | No measurable | CC, bYNBg | [18] |
| Ketoconazole | C. albicans (50) | 0.004–0.032 | 1 h | 2 × MIC | 0.62 | OD, RPMI | [57] |
| Ketoconazole | C. dubliniensis (20) | 0.002–0.012 | 1 h | 3 × MIC | 0.50–0.75 | OD, RPMI | [12] |
| Ketoconazole | C. glabrata (1) | 1 | 1.5–12 h | 1, 4, 8 × MIC | No measurable, 0–0.3 | CC, bYNBg | [18] |
| Ketoconazole | C. glabrata (14) | 100–520 | 1 h | 1, 2, 4 xMIC | 1.8–5.72, 1.04–5.54, 0.69–5.7 | OD, RPMI | [59] |
| Ketoconazole | C. tropicalis (10) | 0.064–0.125 | 1 h | 2 × MIC | 2.03 | OD, RPMI | [6] |
| Voriconazole | C. guilliermondii (2), C. kefyr (2), C. lusitaniae (2) | 0.12–1 | 1 h | 0.125–8 × MIC | No measurable | CC, RPMI | [19] |
| Antifungal | Species (Strains) | MIC (mg/L) | Exposure | Concentration | PAFE (h) | Methodology * | Reference |
|---|---|---|---|---|---|---|---|
| Anidulafungin | C. africana (2) | 0.003–0.006 | 1 h | 0.12, 0.5, 2 mg/L | 0.7–2.8, 2–>37.7, 36.6–>37.7 | CC, RPMI | [32] |
| Anidulafungin | C. albicans (2) | 0.015 | 1 h | 0.125–4 × MIC | >12 | CC, RPMI | [15] |
| Anidulafungin | C. albicans (4) | 0.008–0.03 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [23] |
| Anidulafungin | C. albicans (7) | 0.003–0.006 | 1 h | 0.12, 0.5, 2 mg/L | 0–>43, 0–>42, 39, 1–>44 | CC, RPMI | [32] |
| Anidulafungin | C. dubliniensis (5) | 0.003–0.006 | 1 h | 0.12, 0.5, 2 mg/L | 0. 0–>42, 18–>44 | CC, RPMI | [32] |
| Anidulafungin | C. glabrata (3) | 0.03–0.06 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [23] |
| Anidulafungin | C. glabrata (2) | 0.06 | 1 h | 0.5, 2, 16 × MIC | >24 | CC, RPMI | [26] |
| Anidulafungin | C. krusei (2) | 0.03–0.06 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [23] |
| Anidulafungin | C. metapsilosis (2) | 1 | 1 h | 0.25, 2, 8 mg/L | 0, 0–2, >24 | CC, RPMI | [32] |
| Anidulafungin | C. orthopsilosis (2) | 1 | 1 h | 0.25, 2, 8 mg/L | 0, 0–2, 42–>44 | CC, RPMI | [32] |
| Anidulafungin | C. parapsilosis (2) | 0.5, 1 | 1 h | 0.5, 2, 16 × MIC | 9, 17, >24, >24 | CC, RPMI | [26] |
| Anidulafungin | C. parapsilosis (3) | 1.0–2.0 | 1 h | 0.25, 2, 8 mg/L | 0, 0–5.7, 5.2–42 | CC, RPMI | [32] |
| Anidulafungin | C. parapsilosis (3) | 1.0–2.0 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [23] |
| Caspofungin | C. africana (2) | 0.5 | 1 h | 0.12, 0.5, 2 mg/L | 0–0.7, 0–0.8, 13.5–37.7 | CC, RPMI | [30] |
| Caspofungin | C. albicans (2) | 0.03 | 1 h | 0.125–4 × MIC | 0–>12 | CC, RPMI | [15] |
| Caspofungin | C. albicans (1) | 0.03 | 1 h | 0.25 mg/L | 5.6 | CC, RPMI | [20] |
| Caspofungin | C. albicans (4) | 0.03–0.25 | 1 h | 1, 4, 16 × MIC | >24 | CC, RPMI | [21] |
| Caspofungin | C. albicans (5) | 0.03–0.125 | 5 min | 0.25, 1, 8 mg/L | 1.4–>24, 1.4–>24, 1.7–3.6 | CC, RPMI | [25] |
| Caspofungin | C. albicans (5) | 0.03–0.125 | 15 min | 0.25, 1, 8 mg/L | 0.09–1.8, 1.2–2.9, 0.8–2.7 | CC, RPMI | [25] |
| Caspofungin | C. albicans (5) | 0.03–0.125 | 30 min | 0.25, 1, 8 mg/L | 0.07–1.9, 0.5–2.1, 0.8–2.5 | CC, RPMI | [25] |
| Caspofungin | C. albicans (5) | 0.03–0.125 | 60 min | 0.25, 1, 8 mg/L | 1.2–>24, 0.02–>24, 1.2–3.0 | CC, RPMI | [25] |
| Caspofungin | C. albicans (5) | 0.015–3.0 | 1 h | 4, 16, 32 mg/L | 4.9–>19.3, 13.8–>19.9, 7.7–>19.9 | CC, RPMI | [28] |
| Caspofungin | C. albicans (5) | 0.125–0.5 | 1 h | 4, 16, 32 mg/L | 0–2.3, 0.3–10.1, 0.2–>18.8 | CC, RPMI + 50% serum | [28] |
| Caspofungin | C. albicans (1) | 4 | 1 h | 4, 16, 32 mg/L | 0 | CC, RPMI | [28] |
| Caspofungin | C. albicans (1) | >32 | 1 h | 4, 16, 32 mg/L | 0 | CC, RPMI + 50% serum | [28] |
| Caspofungin | C. albicans (7) | 0.25–0.5 | 1 h | 0.12, 0.5, 2 mg/L | 0–2.9, 0–2.3, >39.5–>44 | CC, RPMI | [30] |
| Caspofungin | C. albicans (20) | 0.004–0.125 | 1 h | 3 × MIC | 2.14 | OD, RPMI | [13] |
| Caspofungin | C. dubliniensis (20) | 0.003–0.19 | 1 h | 3 × MIC | 2.17 | OD, RPMI | [13] |
| Caspofungin | C. dubliniensis (5) | 0.25–0.5 | 1 h | 0.12, 0.5, 2 mg/L | 0, 0, 20–>42 | CC, RPMI | [30] |
| Caspofungin | C. glabrata (2) | 0.25 | 1 h | 1, 4, 16 × MIC | >24 | CC, RPMI | [21] |
| Caspofungin | C. glabrata (2) | 0.5 | 1 h | 0.5, 2, 16 × MIC | >24, >24, >24 | CC, RPMI | [25] |
| Caspofungin | C. guillermondii (2) | 128 | 1 h | 4, 8 × MIC | No measurable | CC, RPMI | [19] |
| Caspofungin | C. kefyr (2) | 0.25 | 1 h | 4, 8 × MIC | No measurable | CC, RPMI | [19] |
| Caspofungin | C. krusei (30) | 0.25–0.125 | 1 h | 0.25, 1.0, 4.0 × MIC | 12->45, 10–>45, 20->45 | CC, RPMI | [3] |
| Caspofungin | C. lusitaniae (2) | 0.5 | 1 h | 4, 8 × MIC | ** | CC, RPMI | [19] |
| Caspofungin | C. metapsilosis (2) | 1–2 | 1 h | 0.25, 2, 8 mg/L | 0, 0, 6.6–>42 | CC, RPMI | [30] |
| Caspofungin | C. orthopsilosis (2) | 1–2 | 1 h | 0.25, 2, 8 mg/L | 0, 0–2, 9–20 | CC, RPMI | [30] |
| Caspofungin | C. parapsilosis (2) | 0.06–0.5 | 1 h | 1, 4, 16 × MIC | >24 | CC, RPMI | [21] |
| Caspofungin | C. parapsilosis (2) | 1–0.5 | 1 h | 0.5, 2, 16 × MIC | 12, 17, >24, >24 | CC, RPMI | [25] |
| Caspofungin | C parapsilosis (3) | 1–2 | 1 h | 0.25, 2, 8 mg/L | 0, 0, 3.7–11.6 | CC, RPMI | [30] |
| Micafungin | C. africana (2) | 0.06–0.12 | 1 h | 0.12, 0.5, 2 mg/L | 0 h, 0–3 h, >37.5->37.7 | CC, RPMI | [29] |
| Micafungin | C. africana (3) | 0.015 | 1 h | 4, 16, 32 mg/L | 9.7->19.2, >19.2–19.9, >19.20–>20.1 | CC, RPMI | [31] |
| Micafungin | C. albicans (4) | 0.0312–0.125 | 1 h | 0.25, 1, 4 × MIC | 0–0.5,−0.30–4.7, 0.90–>16.6 | CC, RPMI | [63] |
| Micafungin | C. albicans (1) | 0.125 | 1 h | 0.25 mg/L | 5.0 | CC, RPMI | [20] |
| Micafungin | C. albicans (4) | 0.008–0.125 | 1 h | 1, 4, 16 × MIC | ≥ 12 | CC, RPMI | [24] |
| Micafungin | C. albicans (7) | 0.12–0.25 | 1 h | 0.12, 0.5, 2 mg/L | 0–2.4, 00–>43, >39.50–>44 | CC, RPMI | [29] |
| Micafungin | C. albicans (3) | 0.03–1 | 1 h | 4, 16, 32 mg/L | 1.50–>19.3, >18.20–>19.4, >18.2–>19.4 | CC, RPMI | [31] |
| Micafungin | C. dubliniensis (5) | 0.06–0.25 | 1 h | 0.12, 0.5, 2 mg/L | 0 h, 0–20, 42–>44 | CC, RPMI | [29] |
| Micafungin | C. dubliniensis (4) | 0.015–0.63 | 1 h | 4, 16, 32 mg/L | >15.9–> 18.5, >15.9–19.9, >15.9–18.5 | CC, RPMI | [31] |
| Micafungin | C. glabrata (2) | 0.0156–0.0625 | 1 h | 0.25, 1, 4 × MIC | 019, 0.09, 0.12, 0.45, 3.4, 2.6 | CC, RPMI | [63] |
| Micafungin | C. glabrata (2) | 0.015–0.125 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [24] |
| Micafungin | C. glabrata (2) | 0.03 | 1 h | 0.5, 2, 16 × MIC | 0.3–0.8, 7, > 24, >24 | CC, RPMI | [26] |
| Micafungin | C. krusei (2) | 0.5 | 1 h | 0.25, 1, 4 × MIC | 2.4–4.1, ≥4.5, ≥19.5 | CC, RPMI | [63] |
| Micafungin | C. krusei (2) | 0.03–0.06 | 1 h | 1, 4, 16 × MIC | ≥ 12 | CC, RPMI | [24] |
| Micafungin | C. metapsilosis (2) | 2 | 1 h | 0.25, 2, 8 mg/L | 0, 0, 5.4–9.3 | CC, RPMI | [29] |
| Micafungin | C. orthopsilosis (2) | 1 | 1 h | 0.25, 2, 8 mg/L | 0–2, 0–2, 3.8–11 | CC, RPMI | [29] |
| Micafungin | C. parapsilosis (2) | 0.5–1 | 1 h | 1, 4, 16 × MIC | ≥12 | CC, RPMI | [24] |
| Micafungin | C. parapsilosis (2) | 1 | 1 h | 0.5, 2, 16 × MIC | 0.2, 3, 11, 10, >24 | CC, RPMI | [26] |
| Micafungin | C parapsilosis (3) | 1–2 | 1 h | 0.25, 2, 8 mg/L | 0, 0, 5.3–15.7 | CC, RPMI | [29] |
| Micafungin | C. tropicalis (2) | 0.5 | 1 h | 0.25, 1, 4 × MIC | <0.2, 5, 0.4, ≥11.6, 2.6 | CC, RPMI | [63] |
| Antifungal | Species (Strains) | MIC (mg/L) | Exposure | Concentration | PAFE (h) | Methodology * | Reference |
|---|---|---|---|---|---|---|---|
| 5-Fluorocytosine | C. albicans (10) | 0.25–0.50 | 1 h | 2 × MIC | 2.37 | OD, RPMI | [6] |
| 5-Fluorocytosine | C. albicans (1) | 0.5 | 1.5–12 h | 1, 4, 8 × MIC | 0.6–2.6, 2.2–5.2, 4.1–6.8 | CC, bYNBg | [18] |
| 5-Fluorocytosine | C. glabrata (1) | 0.0625 | 1.5–12 h | 1, 4, 8 × MIC | 1–3, 3–6.2, 4.8–10.8 | CC, bYNBg | [18] |
| 5-Fluorocytosine | C. glabrata (14) | 0.0008–0.025 | 1 h | 1, 2, 4 × MIC | 2.2–9.5, 2.5–16.6, 2.8–17.5 | OD, RPMI | [59] |
| 5-Fluorocytosine | C. tropicalis (10) | 0.094–0.125 | 1 h | 2 × MIC | 4.41 | OD, RPMI | [6] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jauregizar, N.; Quindós, G.; Gil-Alonso, S.; Suárez, E.; Sevillano, E.; Eraso, E. Postantifungal Effect of Antifungal Drugs against Candida: What Do We Know and How Can We Apply This Knowledge in the Clinical Setting? J. Fungi 2022, 8, 727. https://doi.org/10.3390/jof8070727
Jauregizar N, Quindós G, Gil-Alonso S, Suárez E, Sevillano E, Eraso E. Postantifungal Effect of Antifungal Drugs against Candida: What Do We Know and How Can We Apply This Knowledge in the Clinical Setting? Journal of Fungi. 2022; 8(7):727. https://doi.org/10.3390/jof8070727
Chicago/Turabian StyleJauregizar, Nerea, Guillermo Quindós, Sandra Gil-Alonso, Elena Suárez, Elena Sevillano, and Elena Eraso. 2022. "Postantifungal Effect of Antifungal Drugs against Candida: What Do We Know and How Can We Apply This Knowledge in the Clinical Setting?" Journal of Fungi 8, no. 7: 727. https://doi.org/10.3390/jof8070727