
Identification and Characterization of Calonectria Species Associated with Plant Diseases in Southern China


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Fungal Isolation
2.3. DNA Extraction and PCR Amplification
2.4. Phylogenetic Analyses
2.5. Pairwise Homoplasy Index (PHI)
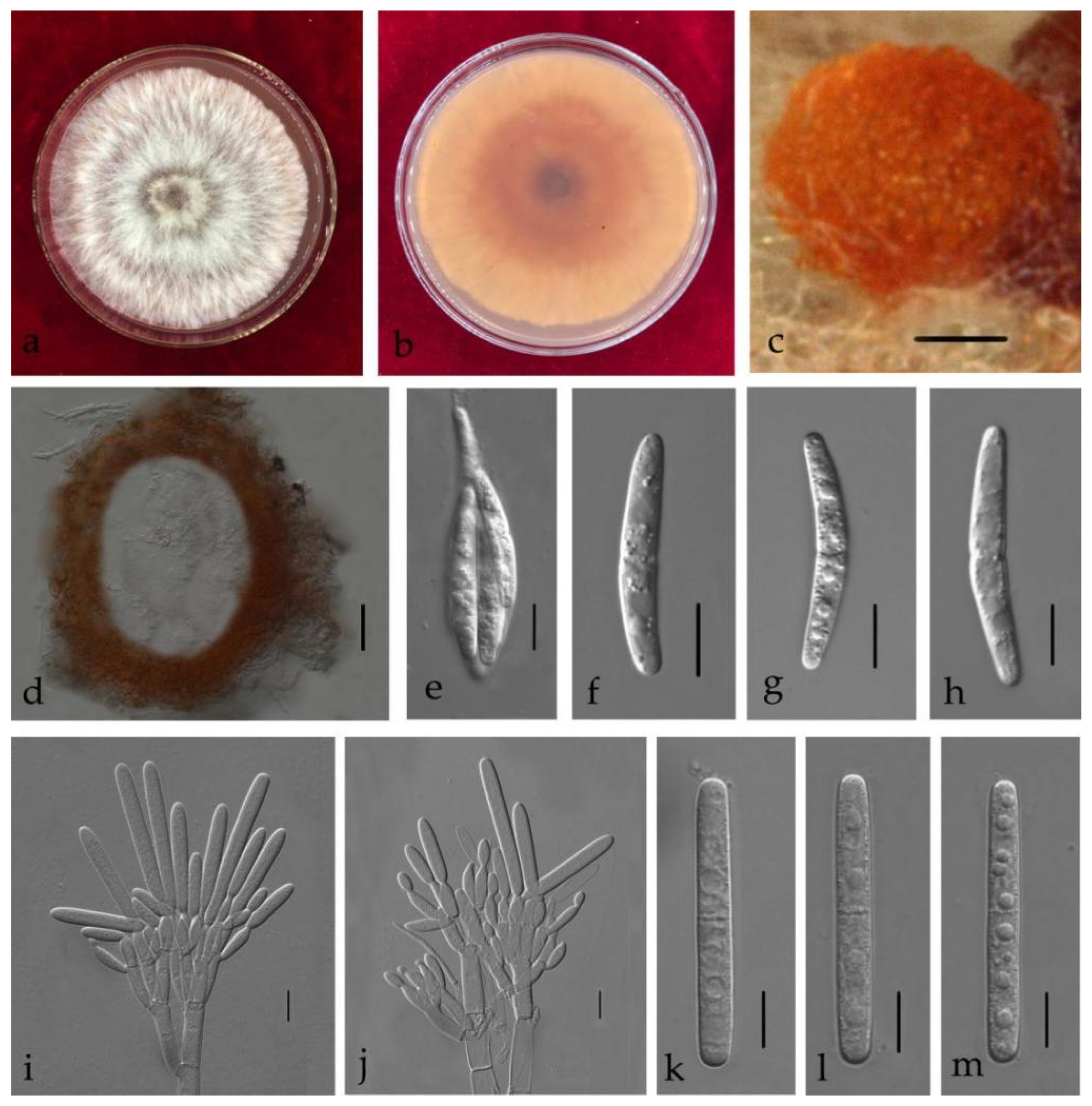
2.6. Morphological Characterisation
3. Results
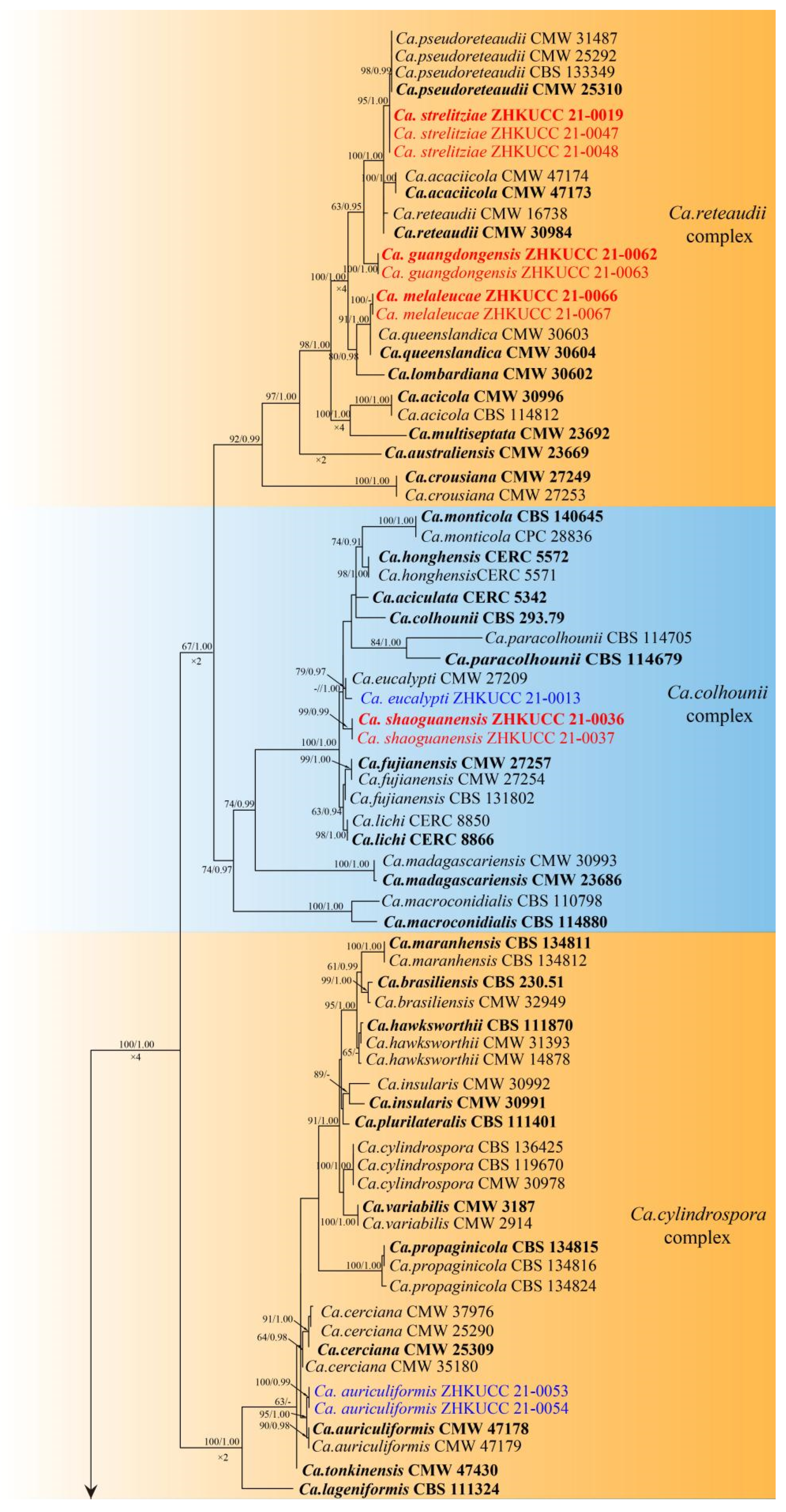
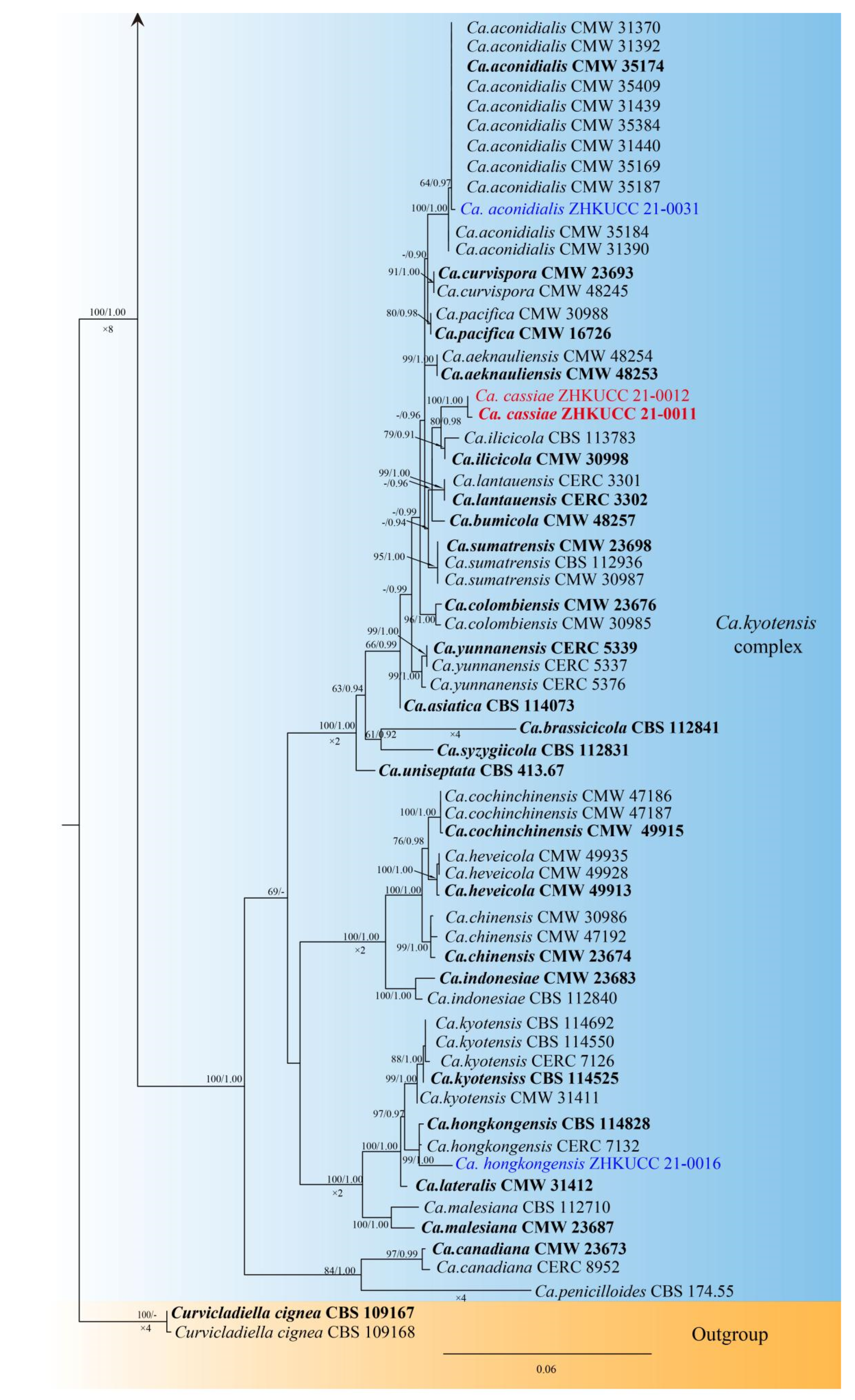
3.1. Phylogenetic Analyses
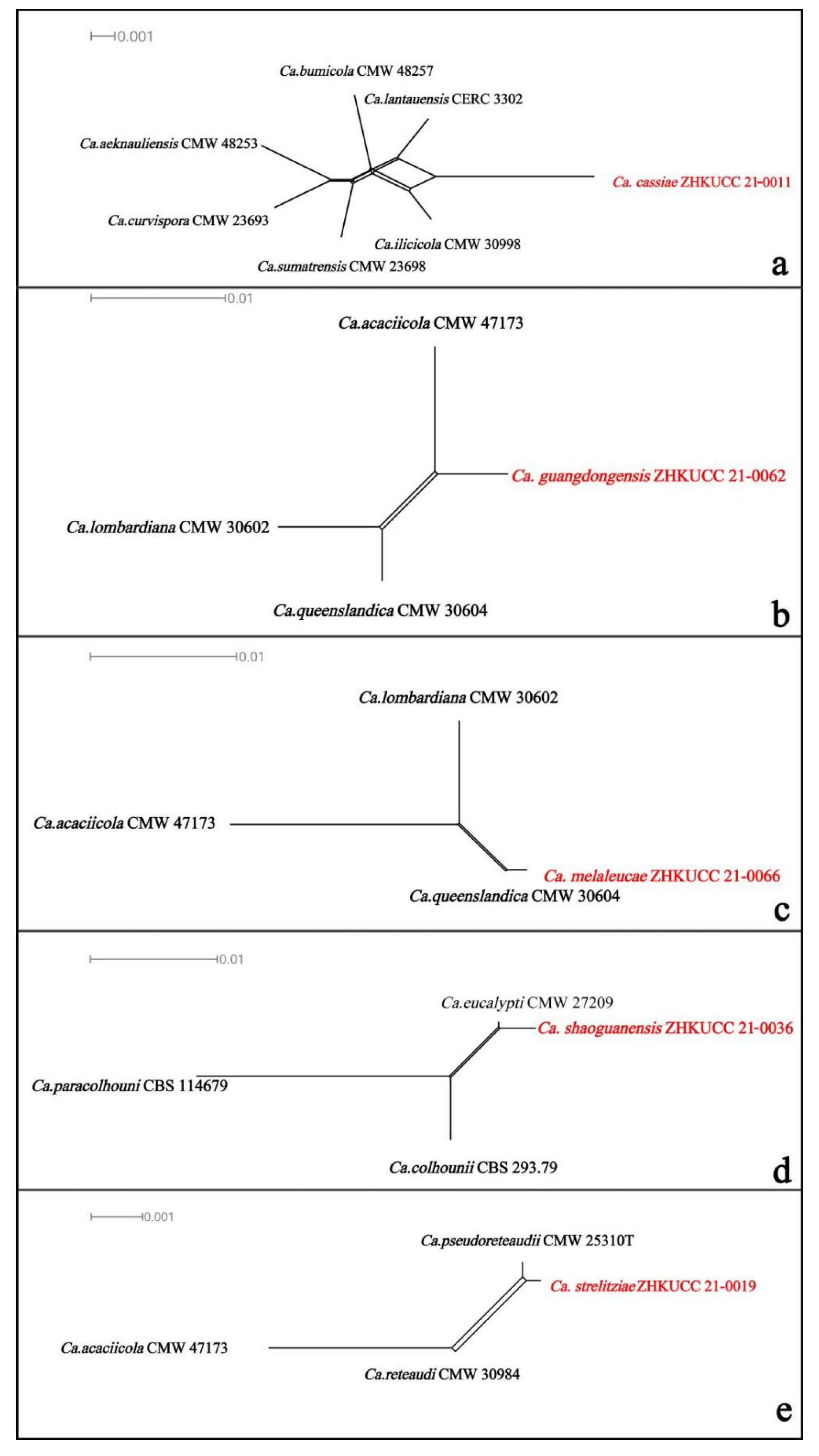
3.2. PHI Analyses
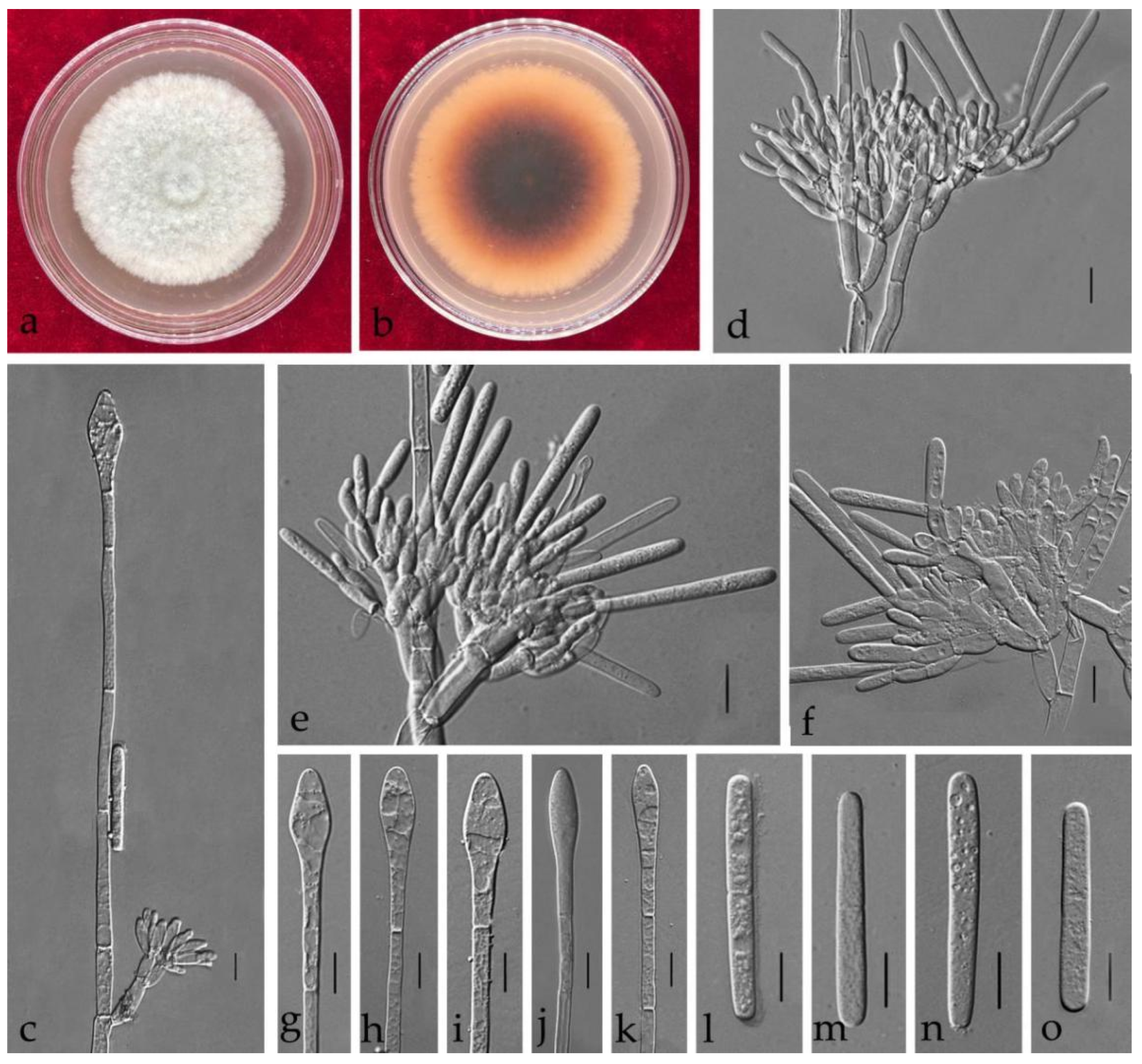
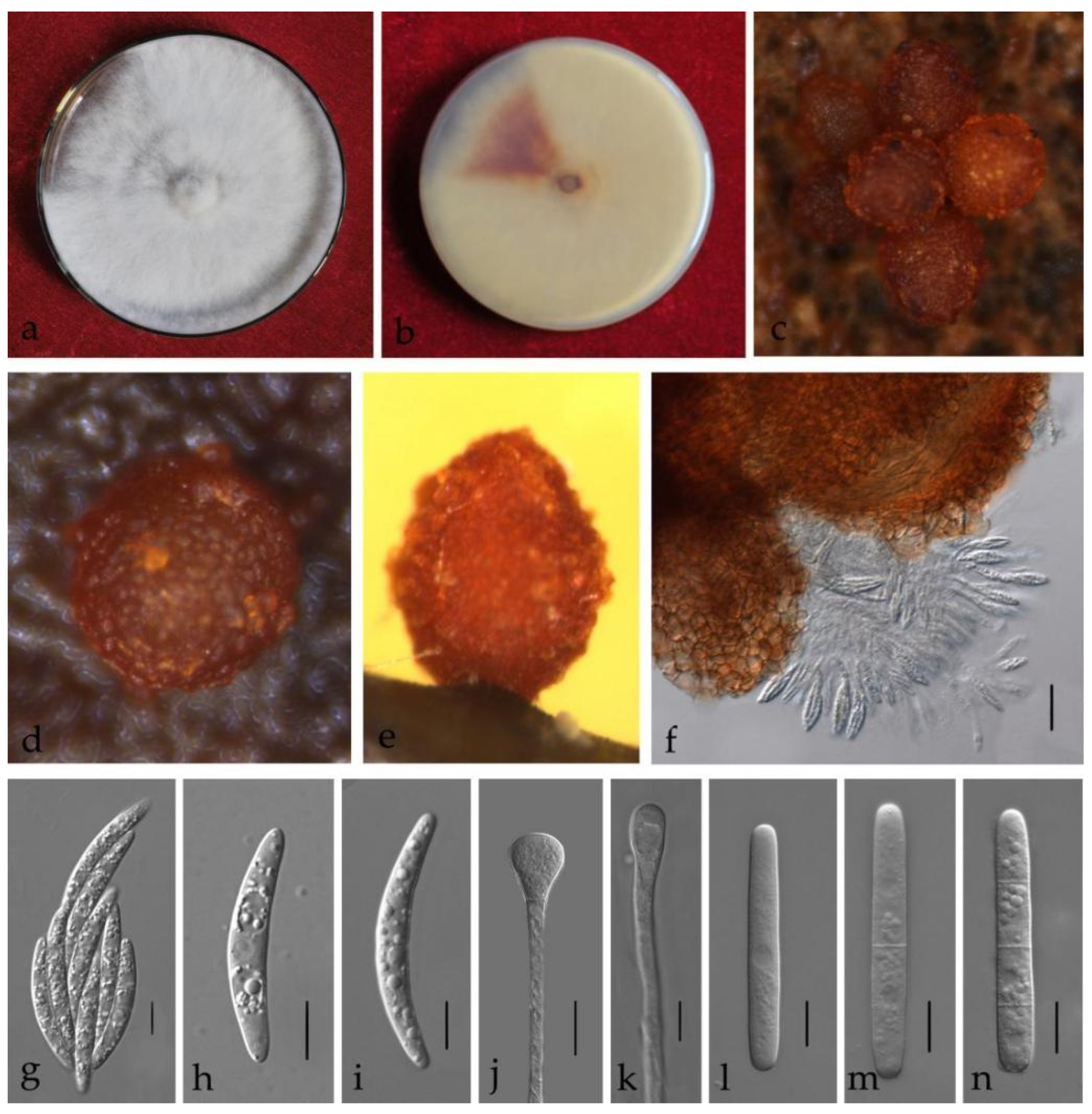
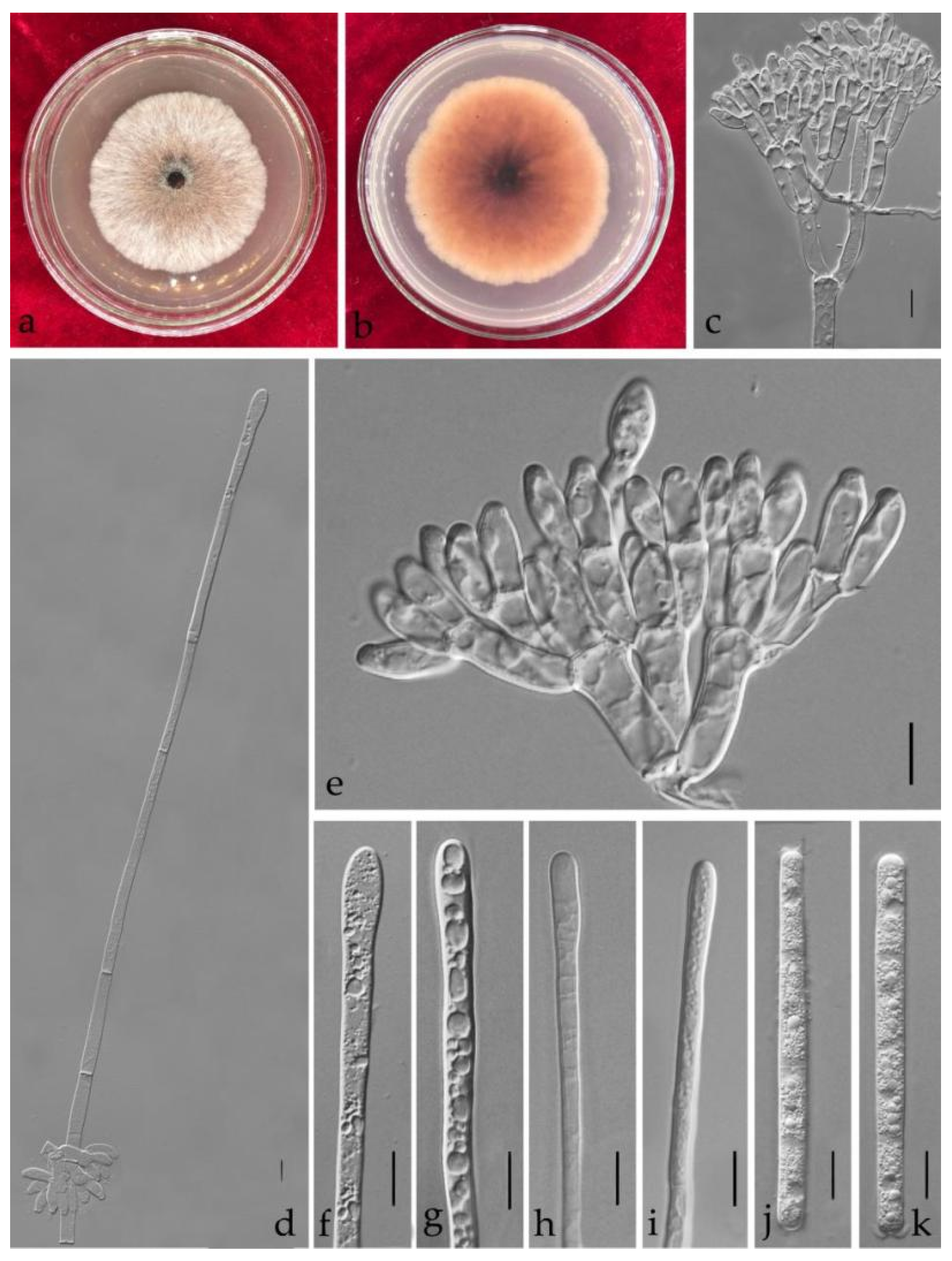
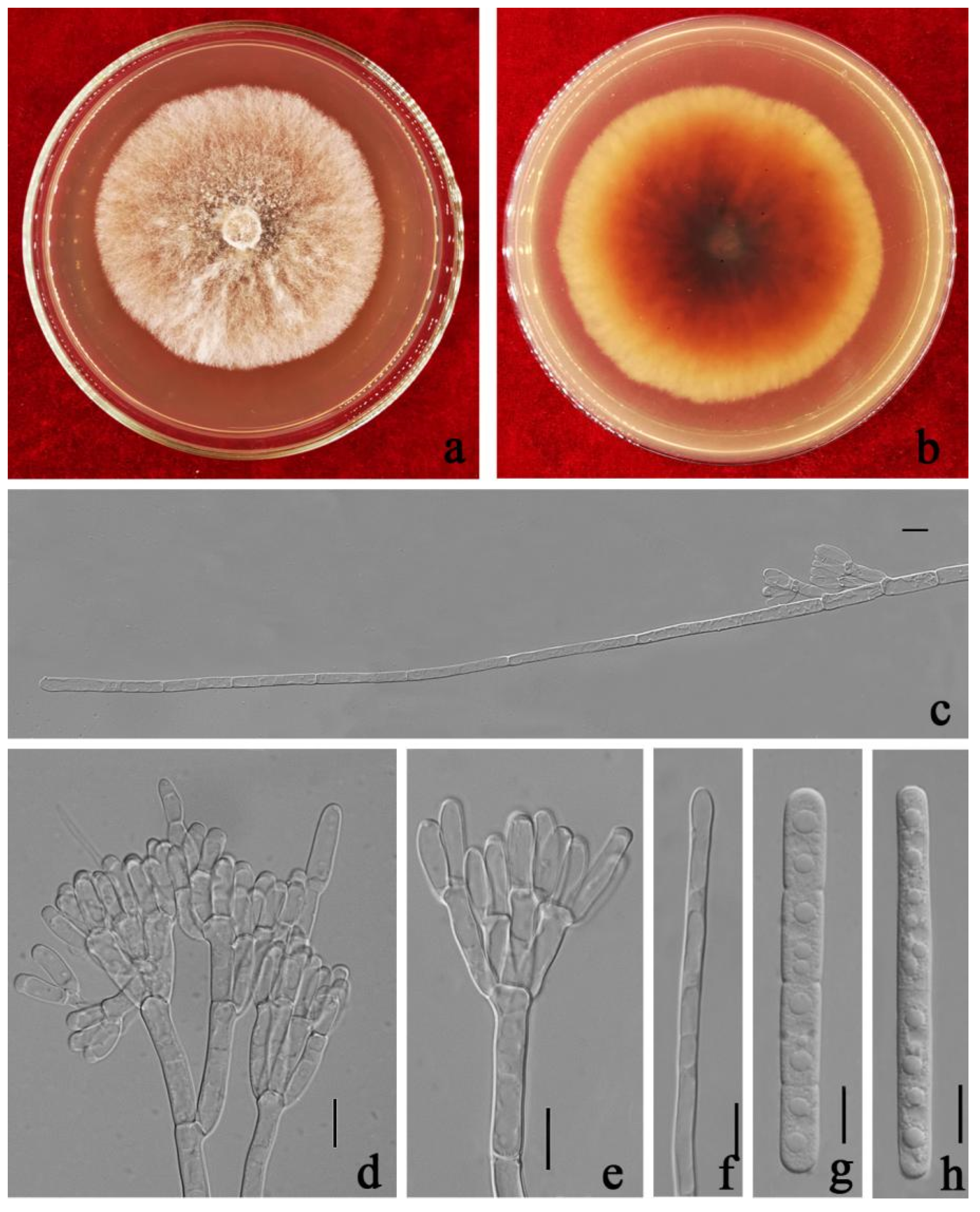
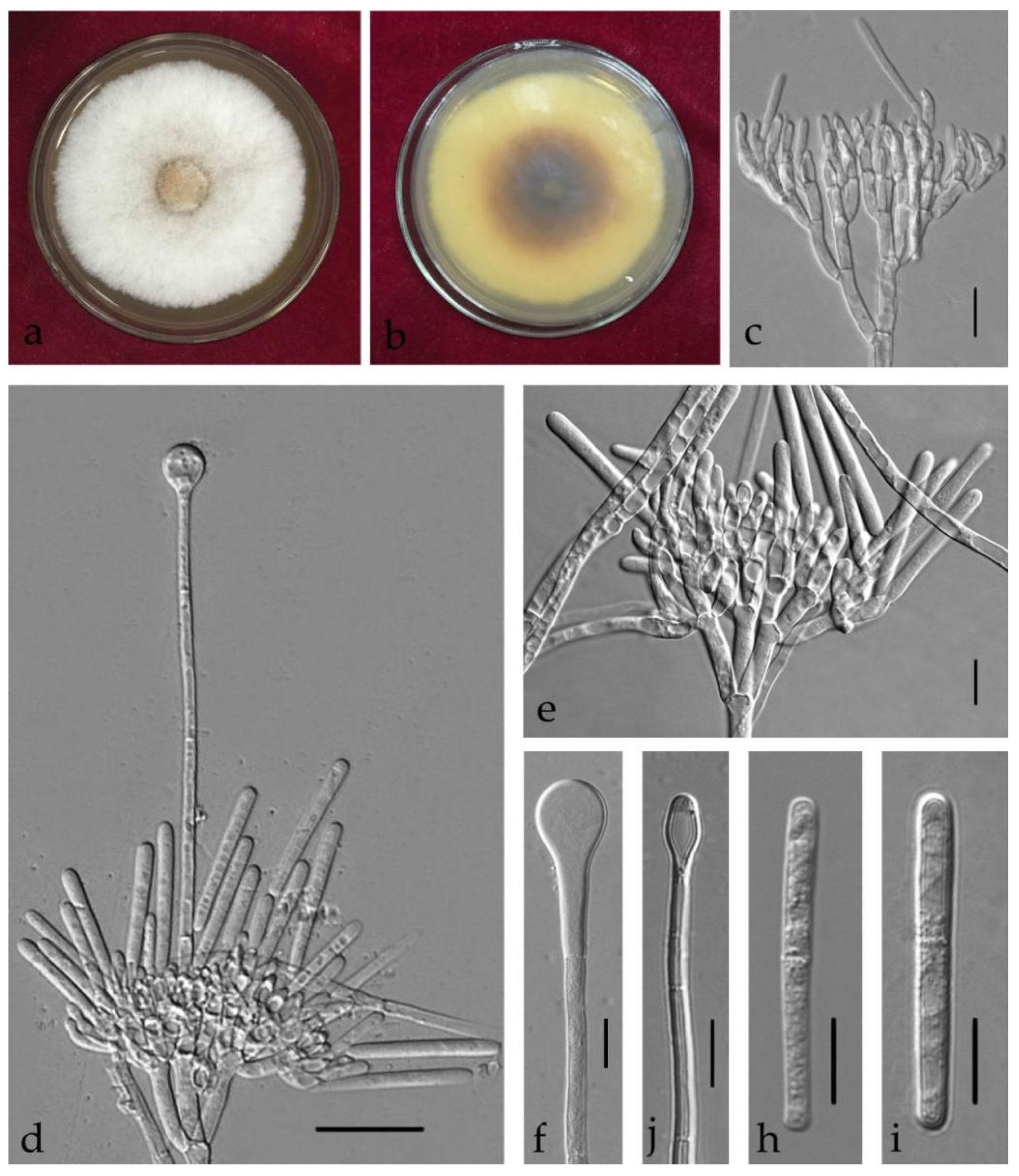
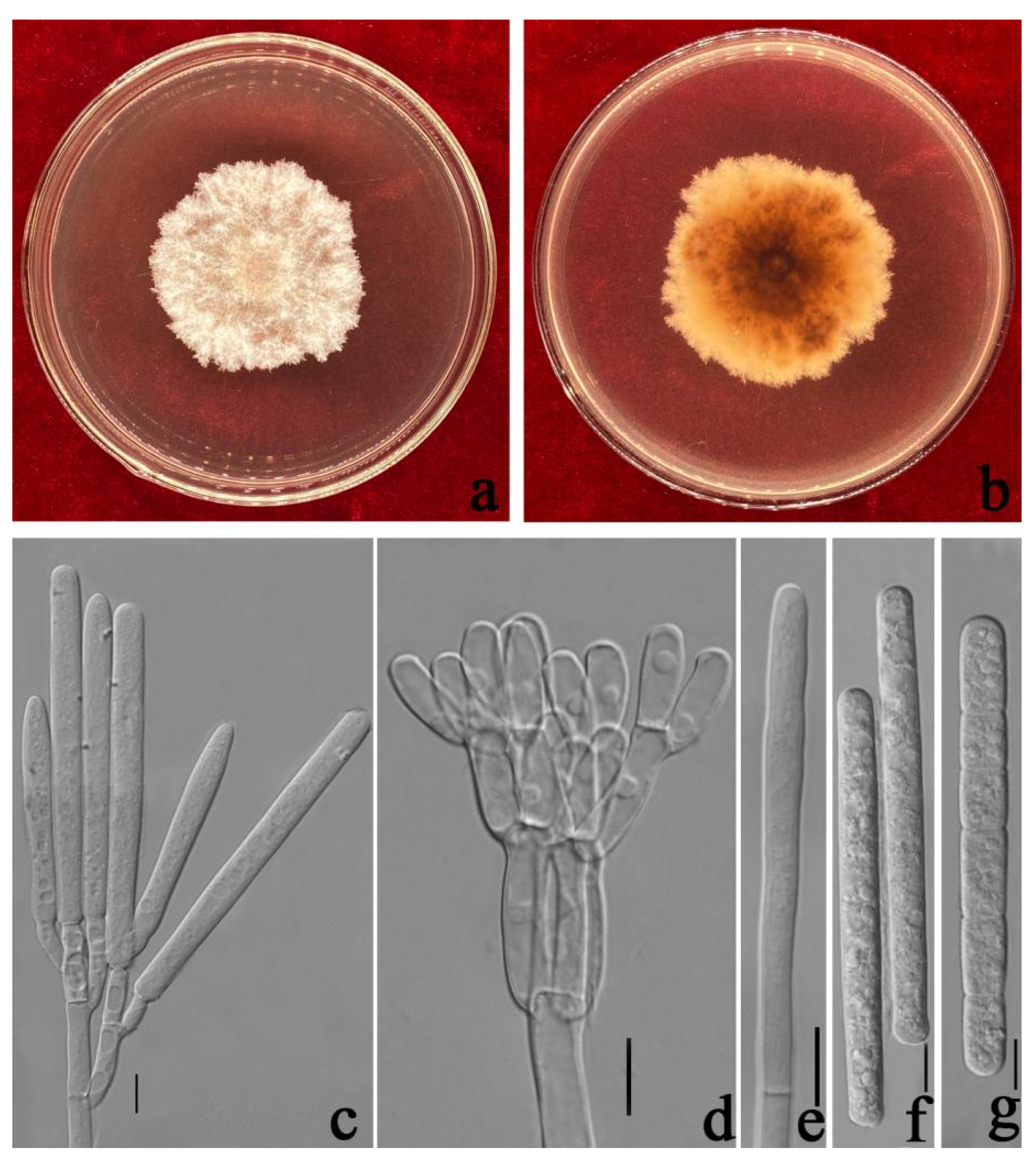
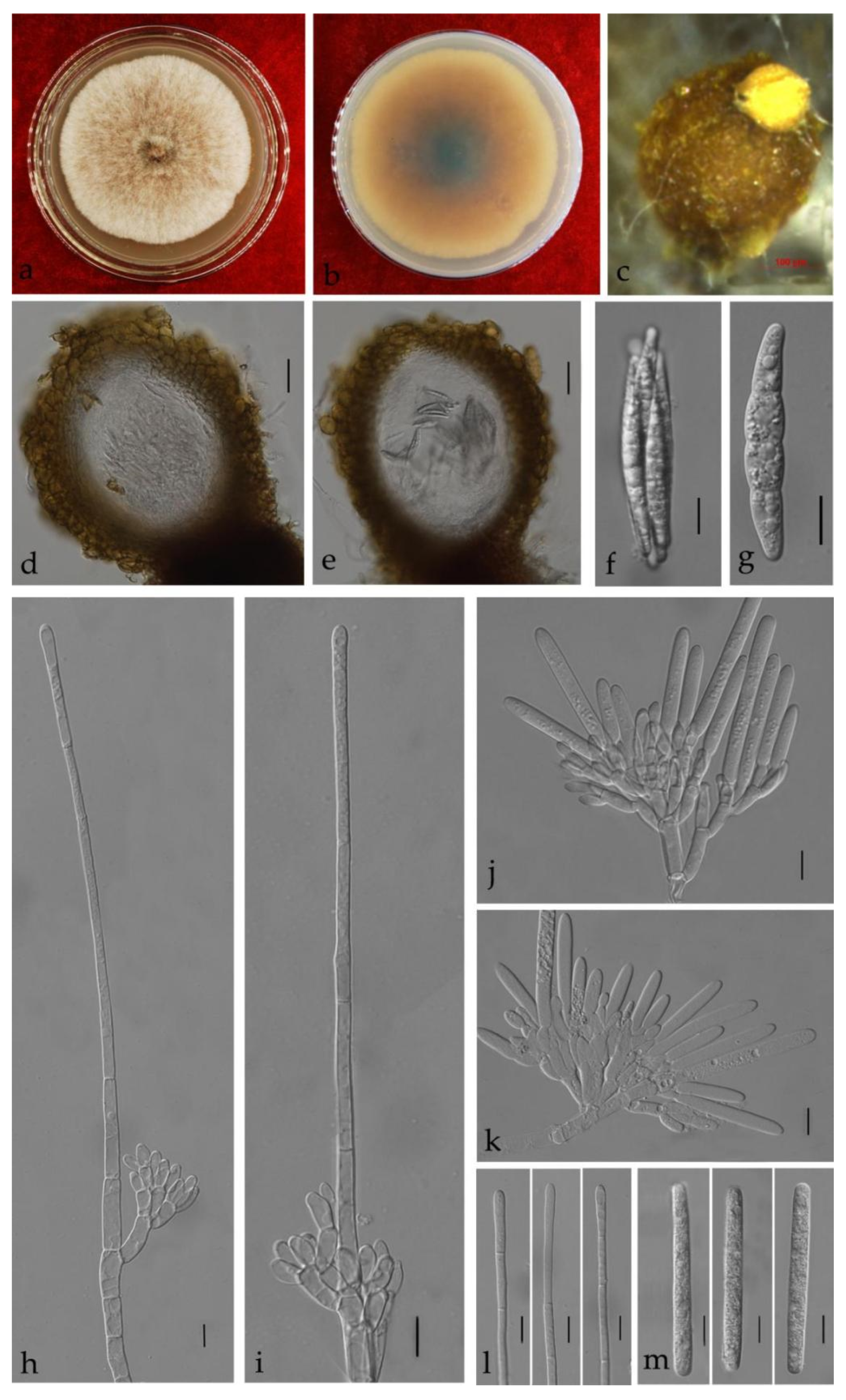
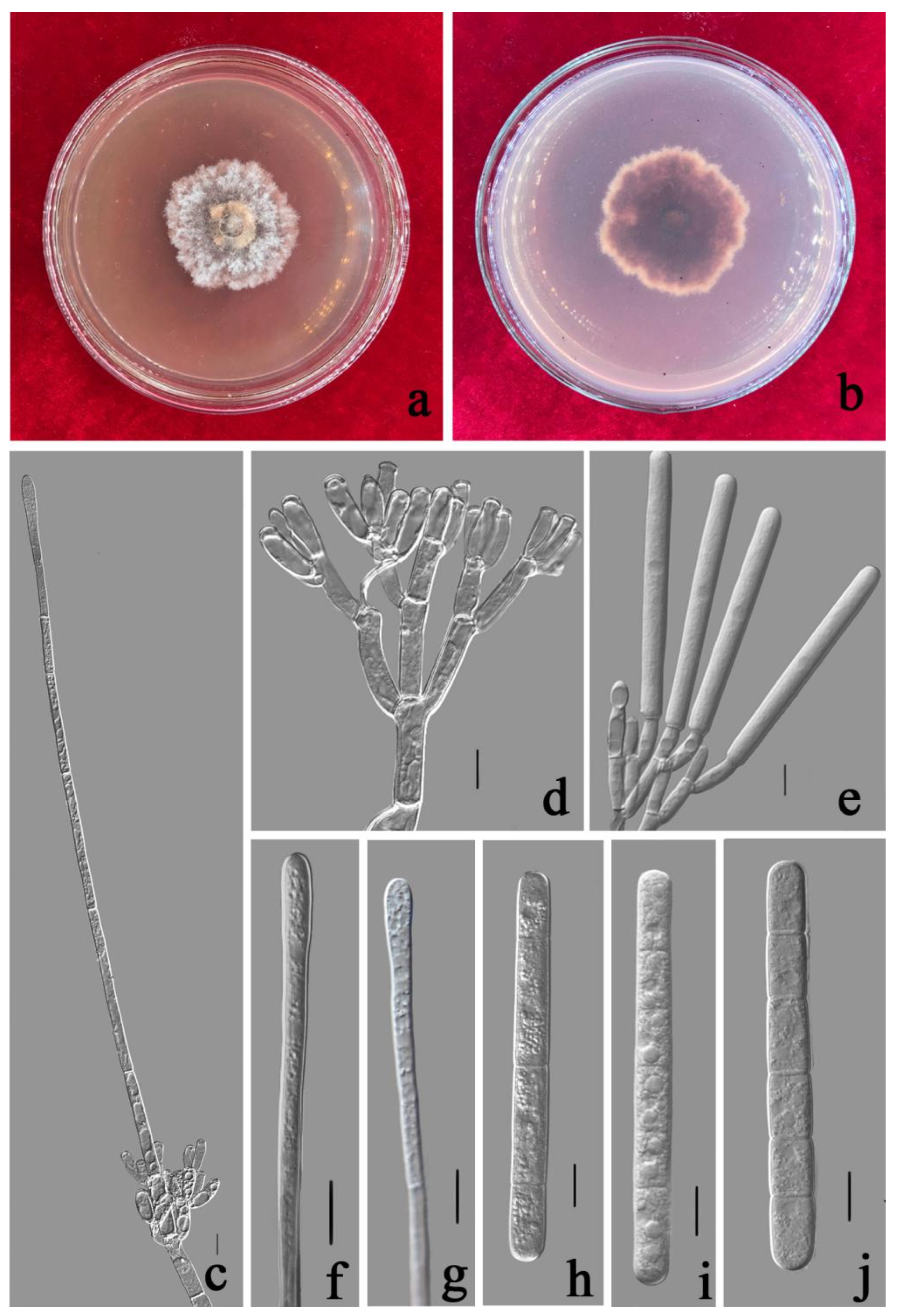
3.3. Taxonomy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Species | Ascospores | Macroconidia | Vesicle | References | |||
|---|---|---|---|---|---|---|---|---|
| Size/μm | Septa | Size/μm | Septa | Shape | Diam/μm | |||
| Ca. colhounii | Ca. eucalypti | (25–)30–36(–56) × (3–)5–6(–8) = 33 × 6 | (1–)3 | (66–)69–75(–80) × (5–)6 = 72 × 6 | 3 | broad clavate | 4–6 | [32] |
| Ca. eucalypti | / | / | (65–)70–80(–87) × (5–)6 = 75 × 6.5 | 1–3 | Clavate to broad clavate | 4–10 | (This study) | |
| Ca. shaoguanensis | (45–)50–70 × (3–)4–8(–9) = 56.5 × 6.5 | 1 | (55–)60–70(–75) × (4–)5–8 = 65 × 6.5 | 1–3 | narrowly clavate | (2–)4–7 | (This study) | |
| Ca. kyotensis | Ca. ilicicola | = 45 × 6 | 1 | = 62 × 6 | 1(–3) | sphaeropedunculate | 8–12 | [32] |
| Ca. cassiae | 40–50 × 4–8 = 45 × 6 | 1 | 40–65 × 4–8 = 54 × 6 | 1–3 | sphaeropedunculate | 8–12 | (This study) | |
| Ca. reteaudii | Ca. strelitziae | / | / | (65–)80–95(–115) × (4–)6–10(–12) = 87 × 8 | 3–5 | narrowly clavate to broad clavate | 3–8 | (This study) |
| Ca. pseudoreteaudii | / | / | (88–)96–112(–119) × 7–9(–10) = 104 × 8 | 5–8 | narrowly clavate | 3–5 | [14] | |
| Ca. acaciicola | / | / | (85–)90–98(–105) × (6–)6.5–7.5 = 94 × 7 | 5 | narrowly clavate | 4–7 | [31] | |
| Ca. reteaudii | / | / | (50–)75–95(–120) × (5–)6–7 = 84 × 6.5 | (1–) 5(–6) | clavate | 3–5(–6) | [9] | |
| Ca. guangdongensis | / | / | 55–70 × 5–7(–9) = 64 × 6 | 1–3 | narrowly clavate | 3–7 | (This study) | |
| Ca. melaleucae | / | / | 80–95(–100) × (5–)7–10 = 88 × 8 | 3–5 | narrowly clavate | 3–7 | (This study) | |
| Ca. queenslandica | / | / | (61–)65–73(–78) × (4–)5–6(–7) = 69 × 6 | 4–6 | narrowly clavate | 3–4 | [14] | |
| Ca. lombardiana | / | / | (64–)74–86(–98) × (5–)5.5–6.5(–7.5) = 80 × 6 | 5 | narrowly clavate | 2–4 | [3] | |
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossman, A.Y. Calonectria and its type species, Calonectria daldiniana, a later synonym of Calonectria pyrochroa. Mycotaxon 1979, 8, 321–328. [Google Scholar]
- Crous, P.W.; Wingfield, M.J. A monograph of Cylindrocladium, including anamorphs of Calonectria. Mycotaxon 1994, 51, 393–404. [Google Scholar]
- Liu, Q.L.; Li, J.; Wingfield, M.J.; Duong, T.A.; Wingfield, B.D.; Crous, P.W.; Chen, S.F. Reconsideration of species boundaries and proposed DNA barcodes for Calonectria. Stud. Mycol. 2020, 97, 1–73. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Wingfield, M.J.; Yilmaz, N.; Adan, O.C.G.; Akulov, A.; et al. New and interesting fungi. 4. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef] [PubMed]
- Mohali, S.R.; Stewart, J.E. Calonectria vigiensis sp. nov. (Hypocreales, Nectriaceae) associated with dieback and sudden-death symptoms of Theobroma cacao from Mérida state, Venezuela. Botany 2021, 99, 683–693. [Google Scholar] [CrossRef]
- Pham, N.Q.; Marincowitz, S.; Chen, S.; Yaparudin, Y.; Wingfield, M.J. Calonectria species, including four novel taxa, associated with Eucalyptus in Malaysia. Mycol. Prog. 2022, 21, 181–197. [Google Scholar] [CrossRef]
- Index Fungorum. 2022. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 11 March 2022).
- Crous, P.W.; Phillips, A.J.L.; Wingfield, M.J. The genera Cylindrocladium and Cylindrocladiella in South Africa, with special reference to forest nurseries. S. Afr. For. J. 1991, 157, 69–85. [Google Scholar]
- Crous, P.W. Taxonomy and Pathology of Cylindrocladium (Calonectria) and Allied Genera; American Phytopathological Society (APS Press.): Saint Paul, MI, USA, 2002. [Google Scholar]
- Pan, R.; Guan, M.; Xu, D.; Gao, X.; Yan, X.; Liao, H. Cylindrocladium black rot caused by Cylindrocludium parasiticum newly reported on peanut in China. Plant Pathol. 2009, 58, 1176. [Google Scholar] [CrossRef]
- Alfenas, R.F.; Lombard, L.; Pereira, O.L.; Alfenas, A.C.; Crous, P.W. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Stud. Mycol. 2015, 80, 89–130. [Google Scholar] [CrossRef] [Green Version]
- Gehesquière, B.; Crouch, J.A.; Marra, R.E.; Van Poucke, K.; Rys, F.; Maes, M.; Heungens, K. Characterization and taxonomic reassessment of the box blight pathogen Calonectria pseudonaviculata, introducing Calonectria henricotiae sp. nov. Plant Pathol. 2016, 65, 37–52. [Google Scholar] [CrossRef]
- Gai, Y.; Deng, Q.; Chen, X.; Guan, M.; Xiao, X.; Xu, D.; Pan, R. Phylogenetic diversity of Calonectria ilicicola causing Cylindrocladium black rot of peanut and red crown rot of soybean in southern China. J. Gen. Plant Pathol. 2017, 83, 273–282. [Google Scholar] [CrossRef]
- Lombard, L.; Zhou, X.D.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria species associated with cutting rot of Eucalyptus. Persoonia 2010, 24, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lombard, L.; Roux, J.; Xie, Y.; Wingfield, M.J.; Zhou, X.D. Novel species of Calonectria associated with Eucalyptus leaf blight in Southeast China. Persoonia 2011, 26, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lombard, L.; Chen, S.F.; Mou, X.; Zhou, X.D.; Crous, P.W.; Wingfield, M.J. New species, hyper-diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Stud. Mycol. 2015, 80, 151–188. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Chen, S. Species diversity, mating strategy and pathogenicity of Calonectria species from diseased leaves and soils in the Eucalyptus plantation in southern China. J. Fungi 2021, 7, 73. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Donnell, K.O.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risède, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Silvestro, D.; Michalak, I. RaxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Gateway Computing Environments Workshop (GCE); Institute of Electrical and Electronics, Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute and British Mycological Society: Kew, UK, 1970. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 11 March 2022).
- Pham, N.Q.; Barnes, I.; Chen, S.; Liu, F.; Dang, Q.N.; Pham, T.Q.; Wingfield, M.J. Ten new species of Calonectria from Indonesia and Vietnam. Mycologia 2019, 111, 78–102. [Google Scholar] [CrossRef]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Phylogeny and systematics of the genus Calonectria. Stud. Mycol. 2010, 66, 31–69. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Tępień, L.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in fungal plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]
- Chethana, K.W.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal. Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Thines, M. What is a species in fungal plant pathogens? Fungal Divers. 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.K. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Ghobad-Nejhad, M.; Wanasinghe, D.N.; Liu, N.; Phillips, A.J.L.; Oliveira-Filho, J.R.C.; da Silva, G.A.; Gibertoni, T.B.; et al. One stop shop ii: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 26–50. Fungal Divers. 2019, 94, 41–129. [Google Scholar] [CrossRef]
- Li, J.; Wingfield, M.J.; Liu, Q.; Barnes, I.; Roux, J.; Lombard, L.; Crous, P.W.; Chen, S.F. Calonectria species isolated from Eucalyptus plantations and nurseries in South China. IMA fungus 2017, 8, 259–286. [Google Scholar] [CrossRef] [Green Version]













| Genes | Primers | Sequence 5′-3′ | Annealing Temperature | References |
|---|---|---|---|---|
| cmdA | CAL-228F | GAGTTCAAGGAGGCCTTCTCCC | 55 °C | [18] |
| CAL-737R | CATCTTTCTGGCCATCATGG | |||
| tef1-α | EF1-728F | CATCGAGAAGTTCGAGAAGG | 54 °C | [18] |
| EF2 | GGARGTACCAGTSATCATGTT | [19] | ||
| β-tubulin | T1 | AACATGCGTGAGATTGTAAGT | 53 °C | [20] |
| CYLTUB1R | AGTTGTCGGGACGGAAGAG | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, C.; Chen, C.; Chen, J.; Xiang, M.; Wanasinghe, D.N.; Hsiang, T.; Hyde, K.D.; Manawasinghe, I.S. Identification and Characterization of Calonectria Species Associated with Plant Diseases in Southern China. J. Fungi 2022, 8, 719. https://doi.org/10.3390/jof8070719
Zhang Y, Chen C, Chen C, Chen J, Xiang M, Wanasinghe DN, Hsiang T, Hyde KD, Manawasinghe IS. Identification and Characterization of Calonectria Species Associated with Plant Diseases in Southern China. Journal of Fungi. 2022; 8(7):719. https://doi.org/10.3390/jof8070719
Chicago/Turabian StyleZhang, Yunxia, Cantian Chen, Chao Chen, Jingwen Chen, Meimei Xiang, Dhanushka N. Wanasinghe, Tom Hsiang, Kevin D. Hyde, and Ishara S. Manawasinghe. 2022. "Identification and Characterization of Calonectria Species Associated with Plant Diseases in Southern China" Journal of Fungi 8, no. 7: 719. https://doi.org/10.3390/jof8070719