
Nitric Oxide Metabolism Affects Germination in Botrytis cinerea and Is Connected to Nitrate Assimilation


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Germination Assays
2.3. Nuclei Staining
2.4. Microscopy
2.5. Statistical Analysis
2.6. Differential Expression Analysis
3. Results
3.1. Pharmacological Analysis
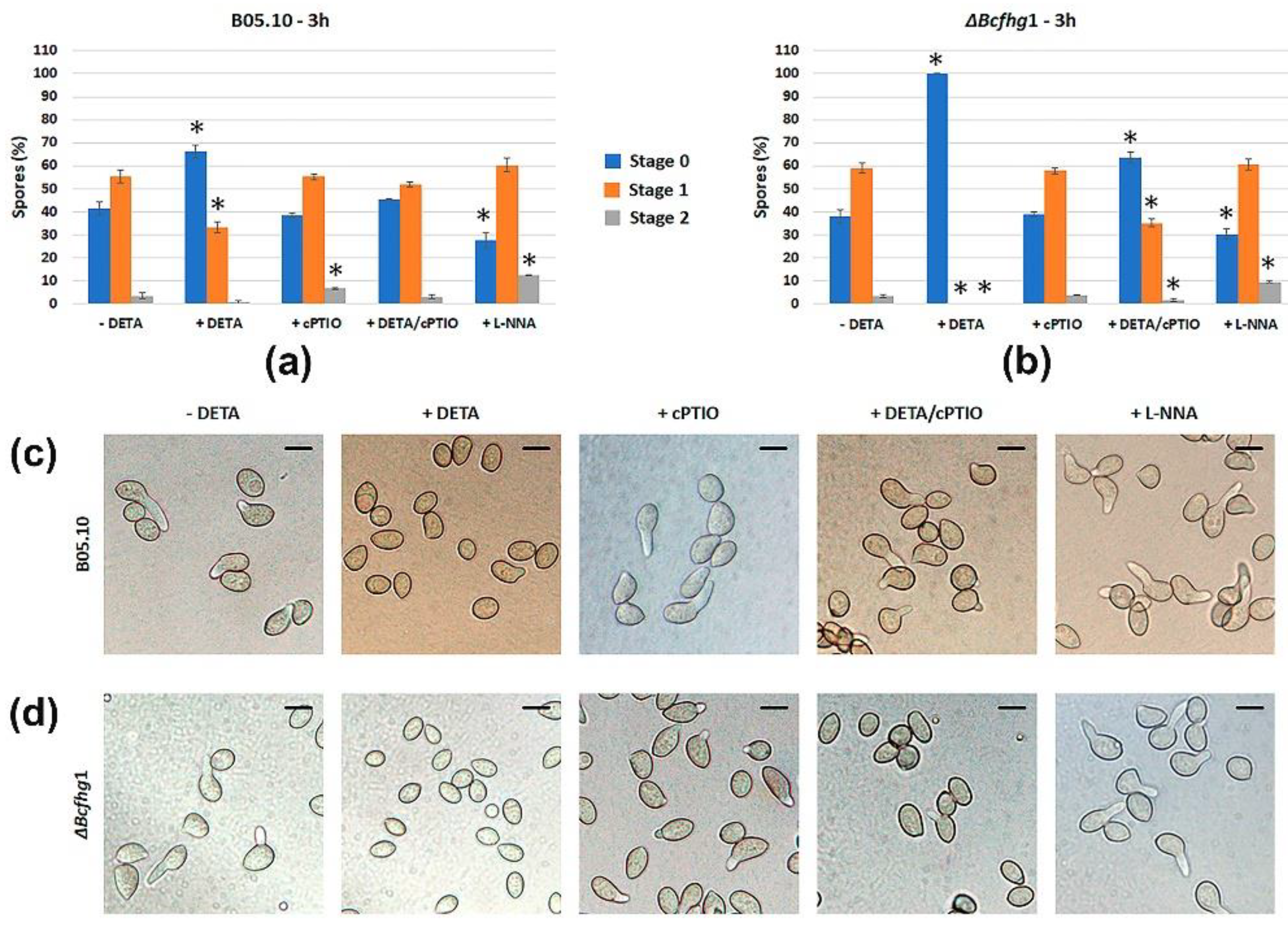
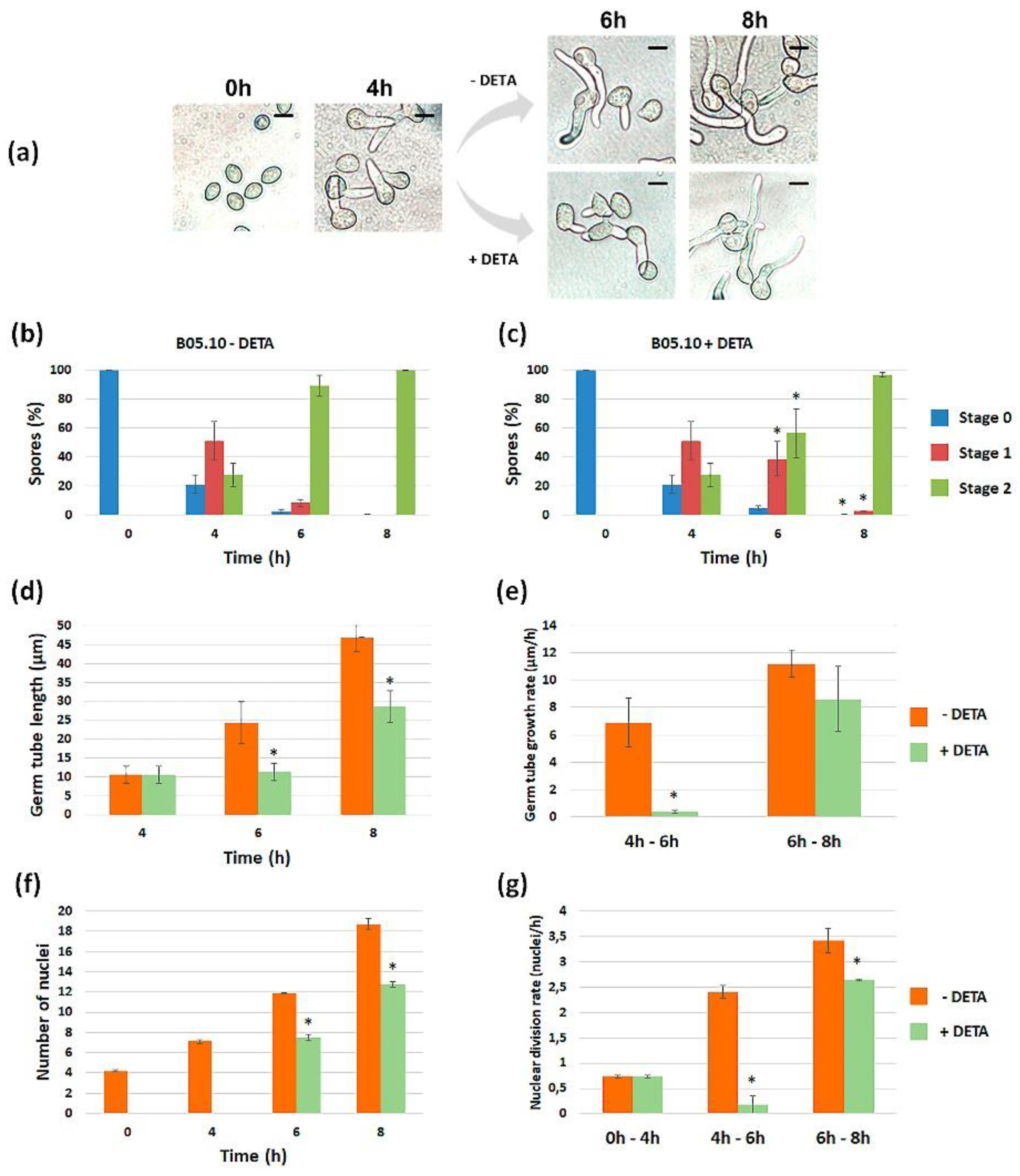
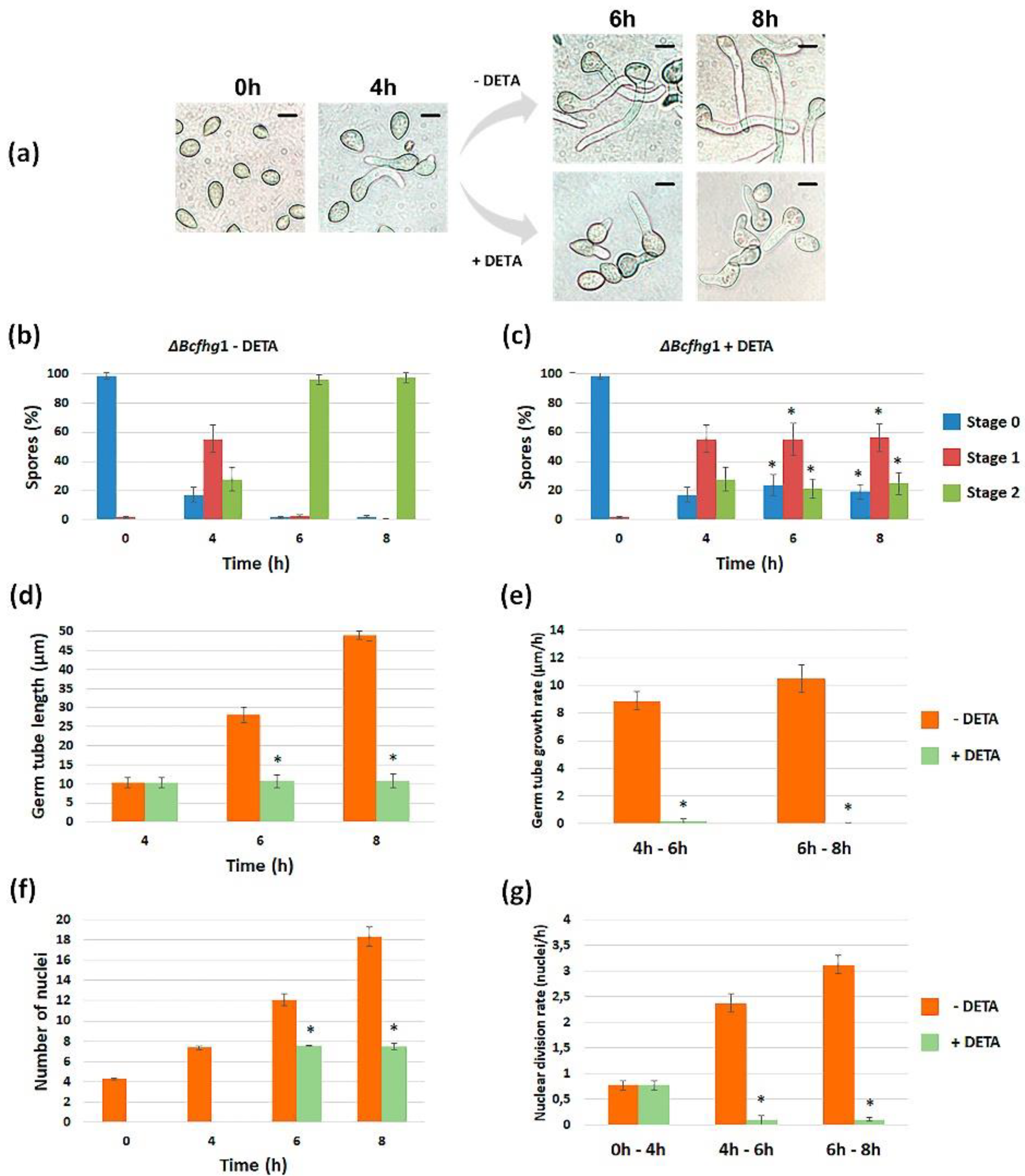
3.1.1. Modulating NO Levels Affects Germination of B. cinerea
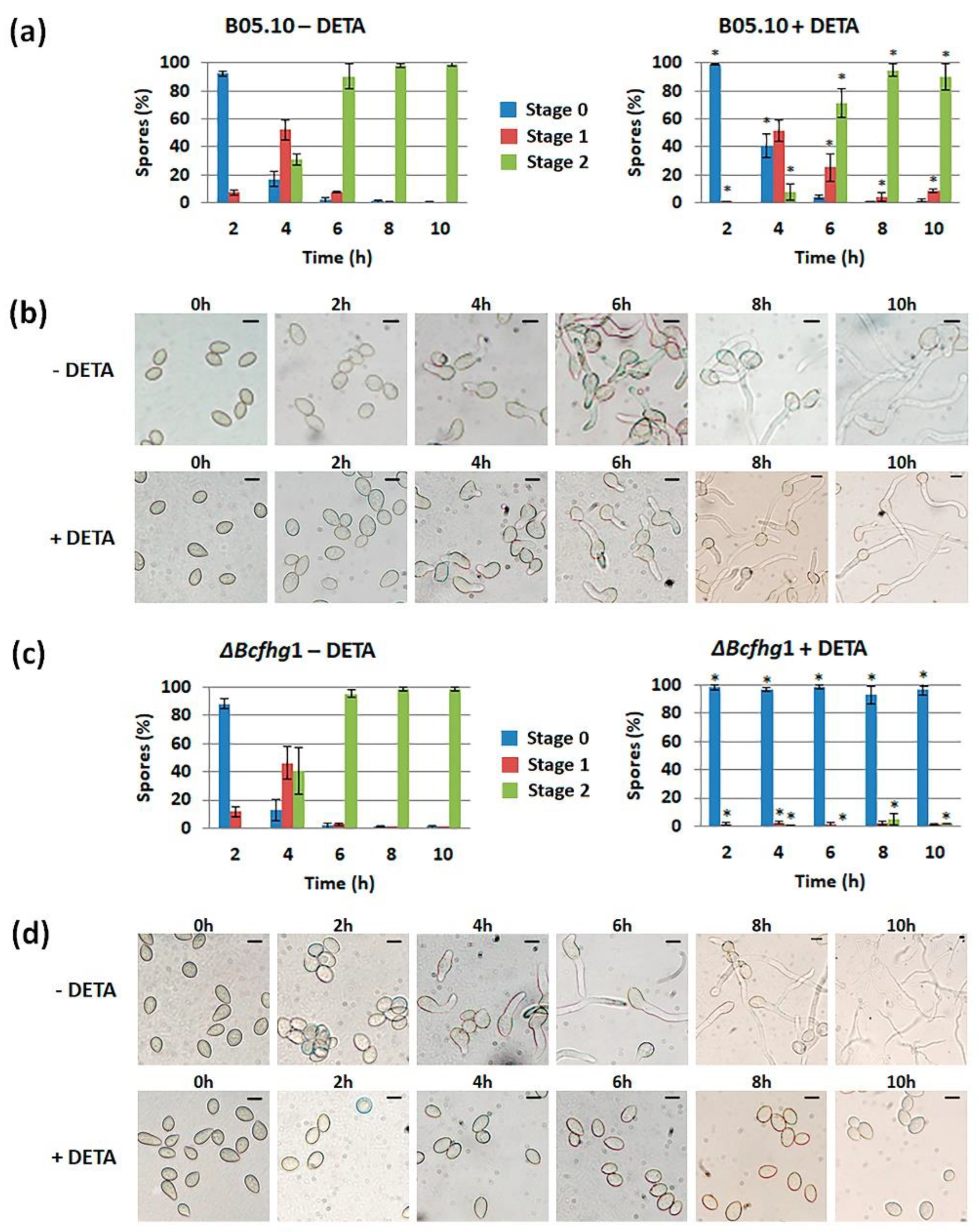
3.1.2. Quantification of the Effect of NO along Time
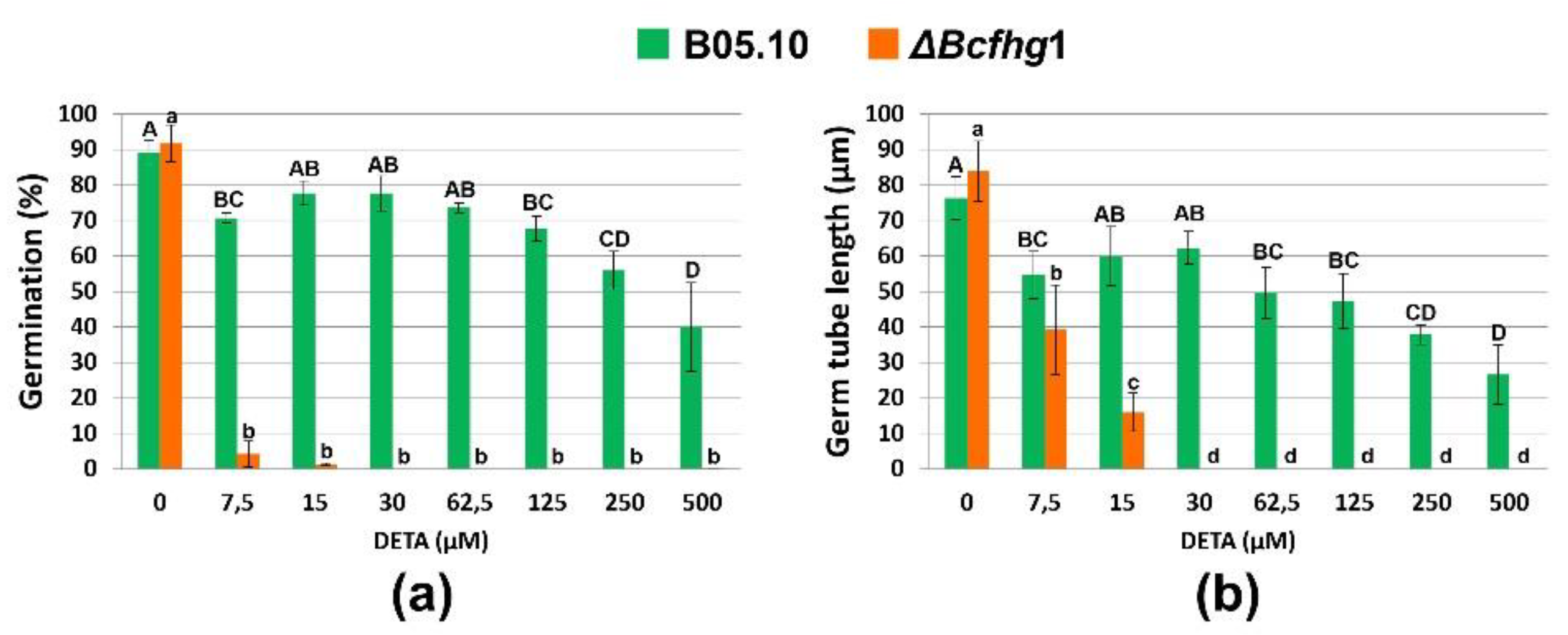
3.1.3. The NO Influence Depends on Its Concentration
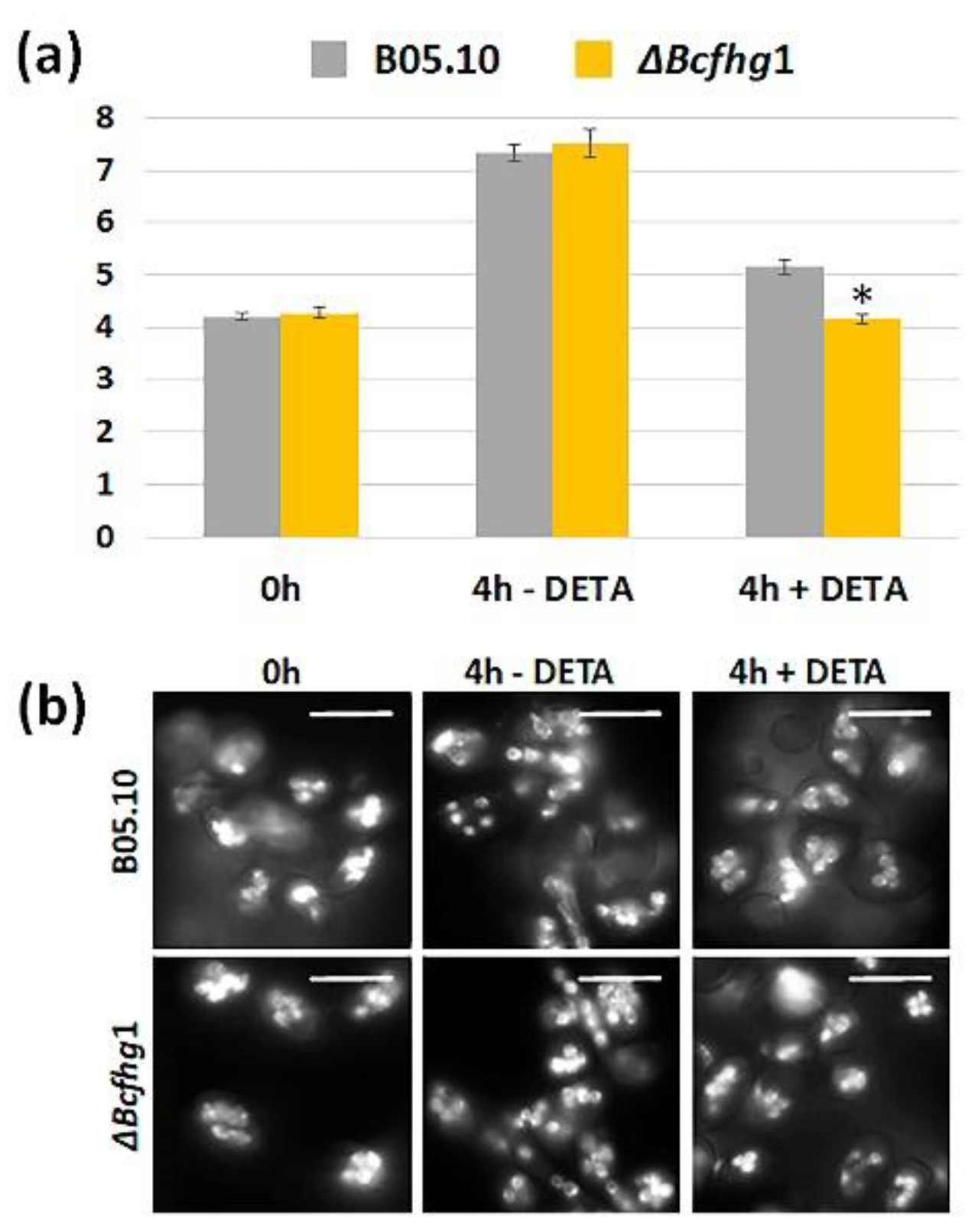
3.1.4. Effect of NO on Nuclear Division during Germination of B. cinerea B05.10 and ΔBcfhg1 Strains
3.1.5. Effect of Exposure to NO Once the Germination Program Has Been Launched
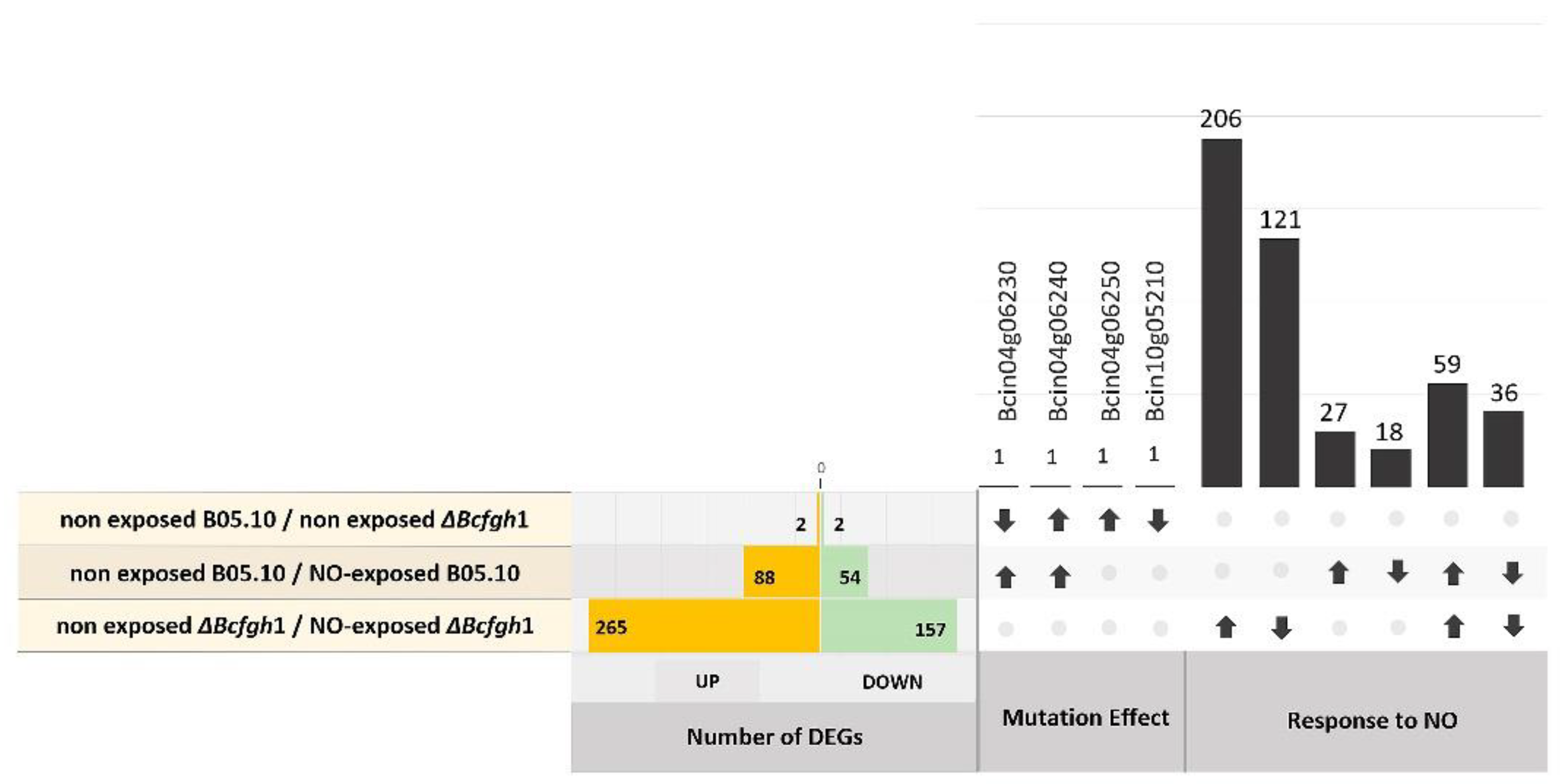
3.2. Differential Gene Expression Analysis in Response to NO Exposure
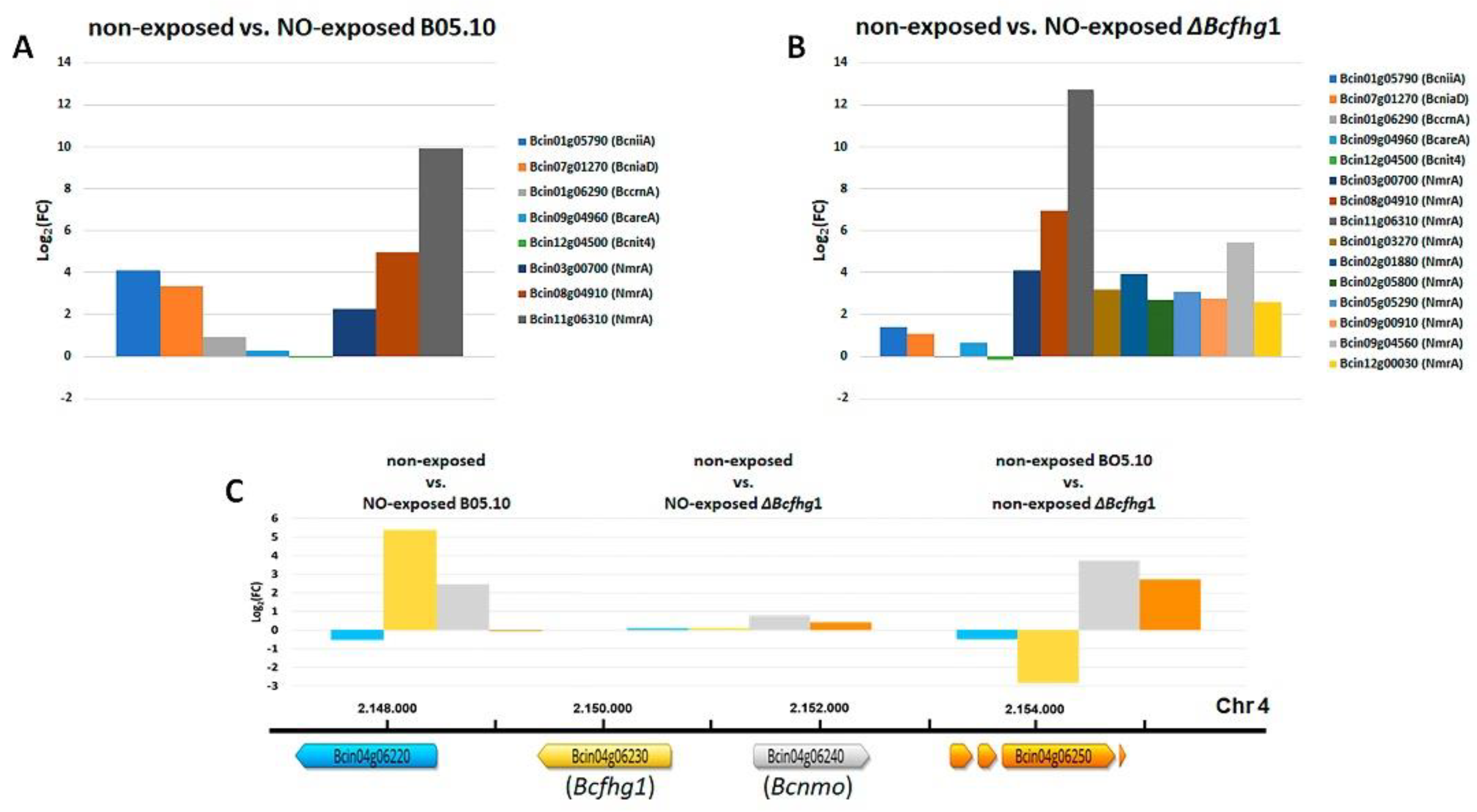
3.2.1. Differential Gene Expression Analysis of the B05.10 Strain Response to NO
3.2.2. Differential Gene Expression Analysis of the ΔBcfhg1 Strain Response to NO
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crane, B.R.; Sudhamsu, J.; Patel, B.A. Bacterial nitric oxide synthases. Annu. Rev. Biochem. 2010, 79, 445–470. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Marcos, J.F.; Marcos, A.T.; Strauss, J. Nitric oxide in fungi: Is there NO light at the end of the tunnel? Curr. Genet. 2016, 62, 513–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippovich, S.Y.; Bachurina, G.P. Nitric Oxide in Fungal Metabolism (Review). Appl. Biochem. Microbiol. 2021, 57, 694–705. [Google Scholar] [CrossRef]
- Mur, L.A.; Mandon, J.; Persijn, S.; Cristescu, S.M.; Moshkov, I.E.; Novikova, G.V.; Hall, M.A.; Harren, F.J.; Hebelstrup, K.H.; Gupta, K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AoB Plants 2013, 5, pls052. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, H.H.; Shonle, H.A.; Grindley, H.S. The origin of the nitrates in the urine. J. Biol. Chem. 1916, 24, 461–490. [Google Scholar] [CrossRef]
- Schmidt, H.; Walter, U. NO at work. Cell 1994, 78, 919–925. [Google Scholar] [CrossRef]
- Astier, J.; Mounier, A.; Santolini, J.; Jeandroz, S.; Wendehenne, D. The evolution of nitric oxide signalling diverges between animal and green lineages. J. Exp. Bot. 2019, 70, 4355–4364. [Google Scholar] [CrossRef]
- Zhao, Y.; Lim, J.; Xu, J.; Yu, J.H.; Zheng, W. Nitric oxide as a developmental and metabolic signal in filamentous fungi. Mol. Microbiol. 2020, 113, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Fu, Y.; Jiang, D.; Xie, J.; Cheng, J.; Li, G.; Hamid, M.I.; Yi, X. Cyclic GMP as a second messenger in the nitric oxide-mediated conidiation of the mycoparasite Coniothyrium minitans. Appl. Environ. Microbiol. 2010, 76, 2830–2836. [Google Scholar] [CrossRef] [Green Version]
- Samalova, M.; Johnson, J.; Illes, M.; Kelly, S.; Fricker, M.; Gurr, S. Nitric oxide generated by the rice blast fungus Magnaporthe oryzae drives plant infection. New Phytol. 2013, 197, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Horchani, F.; Prévot, M.; Boscari, A.; Evangelisti, E.; Meilhoc, E.; Bruand, C.; Raymond, P.; Boncompagni, E.; Aschi-Smiti, S.; Puppo, A.; et al. Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules. Plant Physiol. 2011, 155, 1023–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Higgins, V.J. Nitric oxide has a regulatory effect in the germination of conidia of Colletotrichum coccodes. Fungal Genet. Biol. 2005, 42, 284–292. [Google Scholar] [CrossRef]
- Lai, T.; Li, B.; Qin, G.; Tian, S. Oxidative damage involves in the inhibitory effect of nitric oxide on spore germination of Penicillium expansum. Curr. Microbiol. 2011, 62, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, Y.; Bi, Y.; Li, J.; Bao, G.; Liu, J.; Yu, X. Effects of nitric oxide on growth of Fusarium sulphureum and its virulence to potato tubers. Eur. Food Res. Technol. 2014, 238, 1007–1014. [Google Scholar] [CrossRef]
- Ávila, A.C.; Ochoa, J.; Proaño, K.; Martínez, M.C. Jasmonic acid and nitric oxide protects naranjilla (Solanum quitoense) against infection by Fusarium oxysporum f. sp. quitoense by eliciting plant defense responses. Physiol. Mol. Plant Pathol. 2019, 106, 129–136. [Google Scholar] [CrossRef]
- Do, Y.J.; Kim, D.H.; Jo, M.S.; Kang, D.G.; Lee, S.W.; Kim, J.-W.; Hong, J.K. In vitro Conidial Germination and Mycelial Growth of Fusarium oxysporum f. sp. fragariae Coordinated by Hydrogen Peroxideand Nitric Oxide-signalling. Korean J. Mycol. 2019, 47, 219–232. [Google Scholar]
- Kunert, J. Effect of nitric oxide donors on survival of conidia, germination and growth of Aspergillus fumigatus in vitro. Folia Microbiol. 1995, 40, 238–244. [Google Scholar] [CrossRef]
- Lazar, E.; Wills, R.; Ho, B.; Harris, A.; Spohr, L. Antifungal effect of gaseous nitric oxide on mycelium growth, sporulation and spore germination of the postharvest horticulture pathogens, Aspergillus niger, Monilinia fructicola and Penicillium italicum. Lett. Appl. Microbiol. 2008, 46, 688–692. [Google Scholar] [CrossRef]
- Huang, H.; Huang, M.; Lv, W.; Hu, Y.; Wang, R.; Zheng, X.; Ma, Y.; Chen, C.; Tang, H. Inhibition of Trichophyton rubrum by 420-nm Intense Pulsed Light. In Vitro Activity and the Role of Nitric Oxide in Fungal Death. Front. Pharmacol. 2019, 10, 1143. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Gao, Z.; Wang, C.; Huang, L.; Kang, Z.; Zhang, H. Nitric oxide and reactive oxygen species coordinately regulate the germination of Puccinia striiformis f. sp. tritici Urediniospores. Front. Microbiol. 2016, 7, 178. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Fu, Y.; Jiang, D.; Li, G.; Yi, X.; Peng, Y. L-arginine is essential for conidiation in the filamentous fungus Coniothyrium minitans. Fungal Genet. Biol. 2007, 44, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Abaitua, F.; Rementería, A.; Millan, R.S.; Eguzkiza, A.; Rodriguez, J.A.; Pontón, J.; Sevilla, M.J. In vitro survival and germination of Candida albicans in the presence of nitrogen compounds. Microbiology 1999, 145, 1641–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pengkit, A.; Jeon, S.S.; Son, S.J.; Shin, J.H.; Baik, K.Y.; Choi, E.H.; Park, G. Identification and functional analysis of endogenous nitric oxide in a filamentous fungus. Sci. Rep. 2016, 6, 30037. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.; Hecker, R.; Rockel, P.; Ninnemann, H. Role of nitric oxide synthase in the light-induced development of sporangiophores in Phycomyces blakesleeanus. Plant Physiol. 2001, 126, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Prado, A.M.; Porterfield, D.M.; Feijo, J.A. Nitric oxide is involved in growth regulation and re-orientation of pollen tubes. Development 2004, 131, 2707–2714. [Google Scholar] [CrossRef] [Green Version]
- Prado, A.M.; Colaço, R.; Moreno, N.; Silva, A.C.; Feijo, J.A. Targeting of pollen tubes to ovules is dependent on nitric oxide (NO) signaling. Mol. Plant 2008, 1, 703–714. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, X.C.; Zhu-Ge, Q.; Jiang, X.; Wang, W.D.; Fang, W.P.; Chen, X.; Li, X.H. Nitric oxide participates in cold-inhibited Camellia sinensis pollen germination and tube growth partly via cGMP in vitro. PLoS ONE 2012, 7, e52436. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, T.; Zhang, C.; Hao, H.; Liu, P.; Zheng, M.; Baluška, F.; Šamaj, J.; Lin, J. Nitric oxide modulates the influx of extracellular Ca2+ and actin filament organization during cell wall construction in Pinus bungeana pollen tubes. New Phytol. 2009, 182, 851–862. [Google Scholar] [CrossRef]
- Hausladen, A.; Gow, A.J.; Stamler, J.S. Nitrosative stress: Metabolic pathway involving the flavohemoglobin. Proc. Natl. Acad. Sci. USA 1998, 95, 14100–14105. [Google Scholar] [CrossRef] [Green Version]
- Gardner, P.R.; Gardner, A.M.; Martin, L.A.; Salzman, A.L. Nitric oxide dioxygenase: An enzymic function for flavohemoglobin. Proc. Natl. Acad. Sci. USA 1998, 95, 10378–10383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausladen, A.; Gow, A.; Stamler, J.S. Flavohemoglobin denitrosylase catalyzes the reaction of a nitroxyl equivalent with molecular oxygen. Proc. Natl. Acad. Sci. USA 2001, 98, 10108–10112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durner, J.; Gow, A.J.; Stamler, J.S.; Glazebrook, J. Ancient origins of nitric oxide signaling in biological systems. Proc. Natl. Acad. Sci. USA 1999, 96, 14206–14207. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.O.; Orii, Y.; Lloyd, D.; Hughes, M.N.; Poole, R.K. Anoxic function for the Escherichia coli flavohaemoglobin (hmp): Reversible binding of nitric oxide and reduction to nitrous oxide. FEBS Lett. 1999, 445, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Turrion-Gomez, J.L.; Eslava, A.P.; Benito, E.P. The flavohemoglobin BCFHG1 is the main NO detoxification system and confers protection against nitrosative conditions but is not a virulence factor in the fungal necrotroph Botrytis cinerea. Fungal Genet. Biol. 2010, 47, 484–496. [Google Scholar] [CrossRef]
- Turrion-Gomez, J.L.; Benito, E.P. Flux of nitric oxide between the necrotrophic pathogen Botrytis cinerea and the host plant. Mol. Plant Pathol. 2011, 12, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Büttner, P.; Koch, F.; Voigt, K.; Quidde, T.; Risch, S.; Blaich, R.; Brückner, B.; Tudzynski, P. Variations in ploidy among isolates of Botrytis cinerea: Implications for genetic and molecular analyses. Curr. Genet. 1994, 25, 445–450. [Google Scholar] [CrossRef]
- Benito, E.P.; Have, A.; Klooster, J.W.; van Kan, J.A.L. Fungal and plant gene expression during synchronized infection of tomato leaves by Botrytis cinerea. Eur. J. Plant Pathol. 1998, 104, 207–220. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Novodvorska, M.; Hayer, K.; Pullan, S.T.; Wilson, R.; Blythe, M.J.; Stam, H.; Stratford, M.; Archer, D.B. Trancriptional landscape of Aspergillus niger at breaking of conidial dormancy revealed by RNA-sequencing. BMC Genom. 2013, 14, 246. [Google Scholar] [CrossRef] [Green Version]
- Collado, I.G.; Viaud, M. Secondary Metabolism in Botrytis cinerea: Combining Genomic and Metabolomic Approaches. In Botrytis—The Fungus, the Pathogen and its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer: Cham, Switzerland, 2016; pp. 291–313. [Google Scholar]
- Wendehenne, D.; Pugin, A.; Klessig, D.F.; Durner, J. Nitric oxide: Comparative synthesis and signaling in animal and plant cells. Trends Plant Sci. 2001, 6, 177–183. [Google Scholar] [CrossRef]
- Beligni, M.V.; Lamattina, L. Nitric oxide in plants: The history is just beginning. Plant Cell Environ. 2001, 24, 267–278. [Google Scholar] [CrossRef] [Green Version]
- León, J.; Costa-Broseta, Á. Present knowledge and controversies, deficiencies, and misconceptions on nitric oxide synthesis, sensing, and signaling in plants. Plant Cell Environ. 2020, 43, 1–15. [Google Scholar] [CrossRef]
- Poole, R.K. Flavohaemoglobin: The pre-eminent nitric oxide-detoxifying machine of microorganisms. F1000Research 2020, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Astuti, R.I.; Nasuno, R.; Takagi, H. Nitric oxide signaling in yeast. Appl. Microbiol. Biotechnol. 2016, 100, 9483–9497. [Google Scholar] [CrossRef]
- VAN Baarlen, P.; Staats, M.; VAN Kan, J.A. Induction of programmed cell death in lily by the fungal pathogen Botrytis elliptica. Mol. Plant Pathol. 2004, 5, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U.; Amoroso, G.; Köhle, H.; Sültemeyer, D.F. Non-invasive online detection of nitric oxide from plants and some other organisms by mass spectrometry. Plant J. 2004, 38, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Floryszak-Wieczorek, J.; Arasimowicz, M.; Milczarek, G.; Jelen, H.; Jackowiak, H. Only an early nitric oxide burst and the following wave of secondary nitric oxide generation enhanced effective defence responses of pelargonium to a necrotrophic pathogen. New Phytol. 2007, 175, 718–730. [Google Scholar] [CrossRef]
- Almeida, B.; Buttner, S.; Ohlmeier, S.; Silva, A.; Mesquita, A.; Sampaio-Marques, B.; Osório, N.S.; Kollau, A.; Mayer, B.; Leão, C.; et al. NO-mediated apoptosis in yeast. J. Cell Sci. 2007, 120, 3279–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindermayr, C.; Saalbach, G.; Durner, J. Proteomic Identification of S-Nitrosylated Proteins in Arabidopsis. Plant Physiol. 2005, 137, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Guo, K.; Andrés, V.; Walsh, K. Nitric oxide-induced downregulation of Cdk2 activity and cyclin A gene transcription in vascular smooth muscle cells. Circulation 1998, 97, 2066–2072. [Google Scholar] [CrossRef] [Green Version]
- Tanner, F.C.; Meier, P.; Greutert, H.; Champion, C.; Nabel, E.G.; Lüscher, T.F. Nitric oxide modulates expression of cell cycle regulatory proteins: A cytostatic strategy for inhibition of human vascular smooth muscle cell proliferation. Circulation 2000, 101, 1982–1989. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, P.; Heintz, N.H. S-phase arrest by reactive nitrogen species is bypassed by okadaic acid, an inhibitor of protein phosphatases PP1/PP2A. Free. Radic. Biol. Med. 2006, 40, 247–259. [Google Scholar] [CrossRef]
- Majumdar, U.; Biswas, P.; Subhra, S.T.; Maiti, D.; Ghosh, S. Regulation of cell cycle and stress responses under nitrosative stress in Schizosaccharomyces pombe. Free Radic. Biol. Med. 2012, 52, 2186–2200. [Google Scholar] [CrossRef]
- Lai, T.; Chen, Y.; Li, B.; Qin, G.; Tian, S. Mechanism of Penicillium expansum in response to exogenous nitric oxide based on proteomics analysis. J. Proteom. 2014, 103, 47–56. [Google Scholar] [CrossRef]
- Marroquin-Guzman, M.; Hartline, D.; Wright, J.D.; Elowsky, C.; Bourret, T.J.; Wilson, R.A. The Magnaporthe oryzae nitrooxidative stress response suppresses rice innate immunity during blast disease. Nat. Microbiol. 2017, 2, 17054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’reilly, C.; Turner, P.D. The nitrilase family of CN hydrolysing enzymes–A comparative study. J. Appl. Microbiol. 2003, 95, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Pace, H.C.; Brenner, C. The nitrilase superfamily: Classification, structure and function. Genome Biol. 2001, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Punt, P.J.; Strauss, J.; Smit, R.; Kinghorn, J.R.; Van Den Hondel, C.A.; Scazzocchio, C. The intergenic region between the divergently transcribed niiA and niaD genes of Aspergillus nidulans contains multiple NirA binding sites which act bidirectionally. Mol. Cell. Biol. 1995, 15, 5688–5699. [Google Scholar] [CrossRef] [Green Version]
- Gronover, C.S.; Schumacher, J.; Hantsch, P.; Tudzynski, B. A novel seven-helix transmembrane protein BTP1 of Botrytis cinerea controls the expression of GST-encoding genes, but is not essential for pathogenicity. Mol. Plant Pathol. 2005, 6, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Marzluf, G.A. Regulation of nitrogen metabolism and gene expression in fungi. Microbiol. Rev. 1981, 45, 437–461. [Google Scholar] [CrossRef] [PubMed]
- Marzluf, G.A. Genetic regulation of nitrogen metabolism in the fungi. Microbiol. Mol. Biol. Rev. 1997, 61, 17–32. [Google Scholar]
- Pfannmüller, A.; Boysen, J.M.; Tudzynski, B. Nitrate Assimilation in Fusarium fujikuroi Is Controlled by Multiple Levels of Regulation. Front. Microbiol. 2017, 8, 381. [Google Scholar] [CrossRef] [Green Version]
- Burger, G.; Tilburn, J.; Scazzocchio, C. Molecular cloning and functional characterization of the pathway-specific regulatory gene nirA, which controls nitrate assimilation in Aspergillus nidulans. Mol. Cell. Biol. 1991, 11, 795–802. [Google Scholar]
- Kudla, B.; Caddick, M.X.; Langdon, T.; Martinez-Rossi, N.M.; Bennett, C.F.; Sibley, S.; Davies, R.W.; Arst, H.N., Jr. The regulatory gene areA mediating nitrogen metabolite repression in Aspergillus nidulans. Mutations affecting specificity of gene activation alter a loop residue of a putative zinc finger. EMBO J. 1990, 9, 1355–1364. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Transcriptional control of nmrA by the bZIP transcription factor MeaB reveals a new level of nitrogen regulation in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 534–551. [Google Scholar] [CrossRef]
- Schönig, B.; Brown, D.W.; Oeser, B.; Tudzynski, B. Cross-species hybridization with Fusarium verticillioides microarrays reveals new insights into Fusarium fujikuroi nitrogen regulation and the role of AreA and NMR. Eukaryot. Cell 2008, 7, 1831–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinko, T.; Berger, H.; Lee, W.; Gallmetzer, A.; Pirker, K.; Pachlinger, R.; Buchner, I.; Reichenauer, T.; Güldener, U.; Strauss, J. Transcriptome analysis of nitrate assimilation in Aspergillus nidulans reveals connections to nitric oxide metabolism. Mol. Microbiol. 2010, 78, 720–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, H. Nitrite-dependent nitric oxide production pathway: Implications for involvement of active nitrogen species in photoinhibition in vivo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1477–1488. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Shi, Y. Metabolic enzymes and coenzymes in transcription–a direct link between metabolism and transcription? Trends Genet. 2004, 20, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Lamb, H.K.; Ren, J.; Park, A.; Johnson, C.; Leslie, K.; Cocklin, S.; Thompson, P.; Mee, C.; Cooper, A.; Stammers, D.K.; et al. Modulation of the ligand binding properties of the transcription repressor NmrA by GATA-containing DNA and site-directed mutagenesis. Protein Sci. 2004, 13, 3127–3138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrianopoulos, A.; Kourambas, S.; Sharp, J.A.; Davis, M.A.; Hynes, M.J. Characterization of the Aspergillus nidulans nmrA gene involved in nitrogen metabolite repression. J. Bacteriol. 1998, 180, 1973–1977. [Google Scholar] [CrossRef] [Green Version]
- Stammers, D.K.; Ren, J.; Leslie, K.; Nichols, C.E.; Lamb, H.K.; Cocklin, S.; Doods, A.; Hawkins, A.R. The structure of the negative transcriptional regulator NmrA reveals a structural superfamily which includes the short-chain dehydrogenase/reductases. EMBO J. 2001, 20, 6619–6626. [Google Scholar] [CrossRef]
- Lamb, H.K.; Leslie, K.; Dodds, A.L.; Nutley, M.; Cooper, A.; Johnson, C.; Thompson, P.; Stammers, D.K.; Hawkins, A.R. The negative transcriptional regulator NmrA discriminates between oxidized and reduced dinucleotides. J. Biol. Chem. 2003, 278, 32107–32114. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Qiu, M.; Wang, B.; Yin, W.B.; Nie, X.; Qin, Q.; Ren, S.; Yang, K.; Zhang, F.; Zhuang, Z.; et al. Functional Analysis of the Nitrogen Metabolite Repression Regulator Gene nmrA in Aspergillus flavus. Front. Microbiol. 2016, 7, 1794. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, P.; Willmann, A.; Straeten, M.; Kleigrewe, K.; Beyer, M.; Humpf, H.U.; Tudzynski, B. Biosynthesis of the red pigment bikaverin in Fusarium fujikuroi: Genes, their function and regulation. Mol. Microbiol. 2009, 72, 931–946. [Google Scholar] [CrossRef] [PubMed]
- Amselem, J.; Cuomo, C.A.; van Kan, J.A.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; de Vries, R.P.; Dyer, P.S.; Fillinger, S.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, M.E.; Borowsky, A.T.; Joehnk, B.; Steenwyk, J.L.; Shen, X.X.; Sil, A.; Rokas, A. Recurrent loss of abaA, a master regulator of asexual development in filamentous fungi, correlates with changes in genomic and morphological traits. Genome Biol. Evol. 2020, 12, 1119–1130. [Google Scholar] [CrossRef]
- Yu, J.-H. Regulation of development in Aspergillus nidulans and Aspergillus fumigatus. Mycobiology 2010, 38, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.S.; Lee, S.-L.; Wolkow, T.D.; McGuire, S.-L.; Hamer, J.E.; Wood, G.; Osmani, S.A. Interaction between developmental and cell cycle regulators is required for morphogenesis in Aspergillus nidulans. EMBO J. 1999, 18, 6994–7001. [Google Scholar] [CrossRef]
- Borneman, A.R.; Hynes, M.J.; Andrianopoulos, A. The abaA homologue of Penicillium marneffei participates in two developmental programmes: Conidiation and dimorphic growth. Mol. Microbiol. 2000, 38, 1034–1047. [Google Scholar] [CrossRef] [PubMed]
- Son, H.; Kim, M.G.; Min, K.; Seo, Y.S.; Lim, J.Y.; Choi, G.J.; Kim, J.C.; Chae, S.K.; Lee, Y.W. AbaA regulates conidiogenesis in the ascomycete fungus Fusarium graminearum. PLoS ONE 2013, 8, e72915. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C. Regulation of mitochondrial respiration by nitric oxide inhibition of cytochrome c oxidase. Biochim. Biophys. Acta Bioenerg. 2001, 1504, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLoS Genet. 2014, 10, e1004040. [Google Scholar] [CrossRef] [Green Version]
- Badaruddin, M.; Holcombe, L.J.; Wilson, R.A.; Wang, Z.Y.; Kershaw, M.J.; Talbot, N.J. Glycogen metabolic genes are involved in trehalose-6-phosphate synthase-mediated regulation of pathogenicity by the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2013, 9, e1003604. [Google Scholar] [CrossRef] [Green Version]
- Thines, E.; Weber, R.W.; Talbot, N.J. MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 2000, 12, 1703–1718. [Google Scholar] [PubMed] [Green Version]
- Leroch, M.; Kleber, A.; Silva, E.; Coenen, T.; Koppenhofer, D.; Shmaryahu, A.; Valenzuela, P.D.T.; Hahn, M. Transcriptome profiling of Botrytis cinerea conidial germination reveals upregulation of infectionrelated genes during the prepenetration stage. Eukaryot. Cell 2013, 12, 614–626. [Google Scholar] [CrossRef] [Green Version]
- Steinbach, W.J.; Cramer, R.A., Jr.; Perfect, B.Z.; Asfaw, Y.G.; Sauer, T.C.; Najvar, L.K.; Kirkpatrick, W.R.; Patterson, T.F.; Benjamin, D.K., Jr.; Heitman, J.; et al. Calcineurin controls growth, morphology, and pathogenicity in Aspergillus fumigatus. Eukaryot. Cell 2006, 5, 1091–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, F.M.E.; Heinekamp, T.; Härtl, A.; Brakhage, A.A.; Semighini, C.P.; Harris, S.D.; Savoldi, M.; de Gouvêa, P.F.; de Souza, G.M.H.; Goldman, G.H. Functional characterization of the Aspergillus fumigatus calcineurin. Fungal Genet. Biol. 2007, 44, 219–230. [Google Scholar]
- Juvvadi, P.R.; Lamoth, F.; Steinbach, W.J. Calcineurin-mediated regulation of hyphal growth, septation, and virulence in Aspergillus fumigatus. Mycopathologia 2014, 178, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, M.; Brunet-Simon, A.; Brygoo, Y.; Pradier, J.M.; Levis, C. Cyclophilin A and calcineurin functions investigated by gene inactivation, cyclosporin A inhibition and cDNA arrays approaches in the phytopathogenic fungus Botrytis cinerea. Mol. Microbiol. 2003, 50, 1451–1465. [Google Scholar] [CrossRef]
- Harren, K.; Schumacher, J.; Tudzynski, B. The Ca2+ /Calcineurin-Dependent Signaling Pathway in the Gray Mold Botrytis cinerea: The Role of Calcipressin in Modulating Calcineurin Activity. PLoS ONE 2012, 7, e41761. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Sun, C.H.; Chang, H.W.; Yang, S.; Liu, Y.; Zhang, M.Z.; Hou, J.; Zhang, H.; Li, G.H.; Qin, Q.M. Cyclophilin BcCyp2 Regulates Infection-Related Development to Facilitate Virulence of the Gray Mold Fungus Botrytis cinerea. Int. J. Mol. Sci. 2021, 22, 1694. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anta-Fernández, F.; Santander-Gordón, D.; Becerra, S.; Santamaría, R.; Díaz-Mínguez, J.M.; Benito, E.P. Nitric Oxide Metabolism Affects Germination in Botrytis cinerea and Is Connected to Nitrate Assimilation. J. Fungi 2022, 8, 699. https://doi.org/10.3390/jof8070699
Anta-Fernández F, Santander-Gordón D, Becerra S, Santamaría R, Díaz-Mínguez JM, Benito EP. Nitric Oxide Metabolism Affects Germination in Botrytis cinerea and Is Connected to Nitrate Assimilation. Journal of Fungi. 2022; 8(7):699. https://doi.org/10.3390/jof8070699
Chicago/Turabian StyleAnta-Fernández, Francisco, Daniela Santander-Gordón, Sioly Becerra, Rodrigo Santamaría, José María Díaz-Mínguez, and Ernesto Pérez Benito. 2022. "Nitric Oxide Metabolism Affects Germination in Botrytis cinerea and Is Connected to Nitrate Assimilation" Journal of Fungi 8, no. 7: 699. https://doi.org/10.3390/jof8070699