
Leaf-Associated Epiphytic Fungi of Gingko biloba, Pinus bungeana and Sabina chinensis Exhibit Delicate Seasonal Variations

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. Quantitative PCR Analysis
2.4. High-Throughput Sequencing
2.5. Statistical Analysis
3. Results
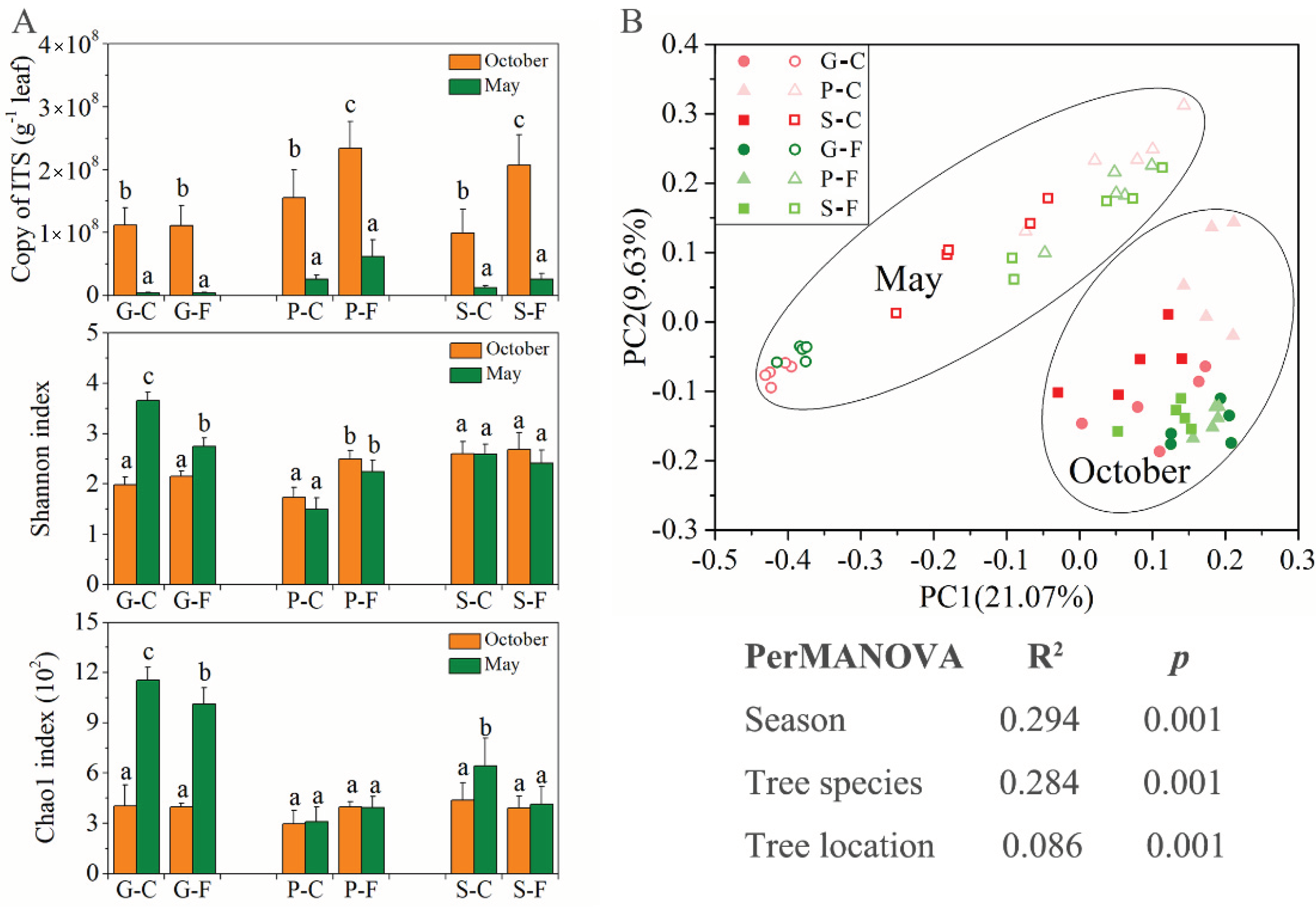
3.1. High Abundance and Diversity of Leaf-Associated Fungi
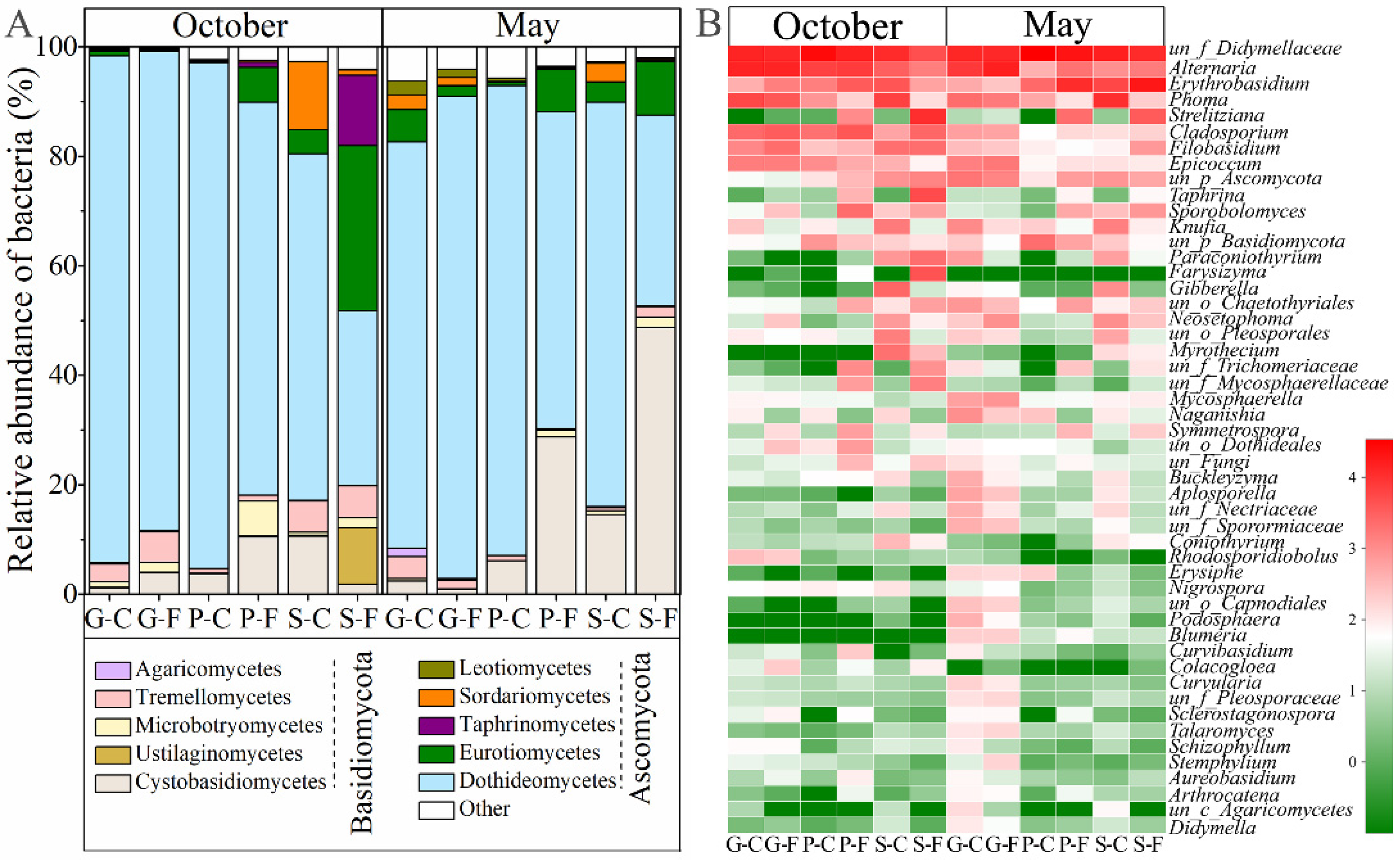
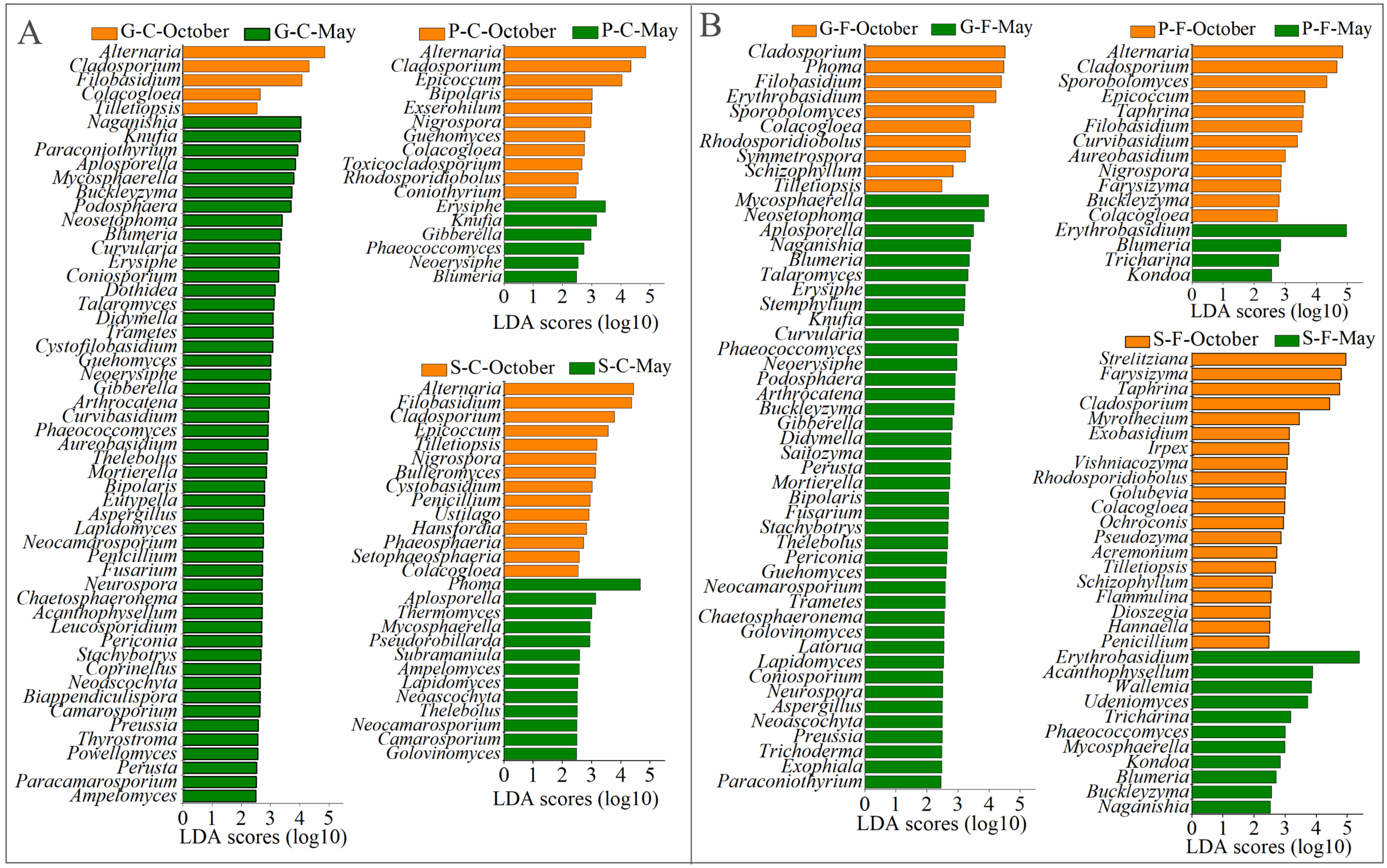
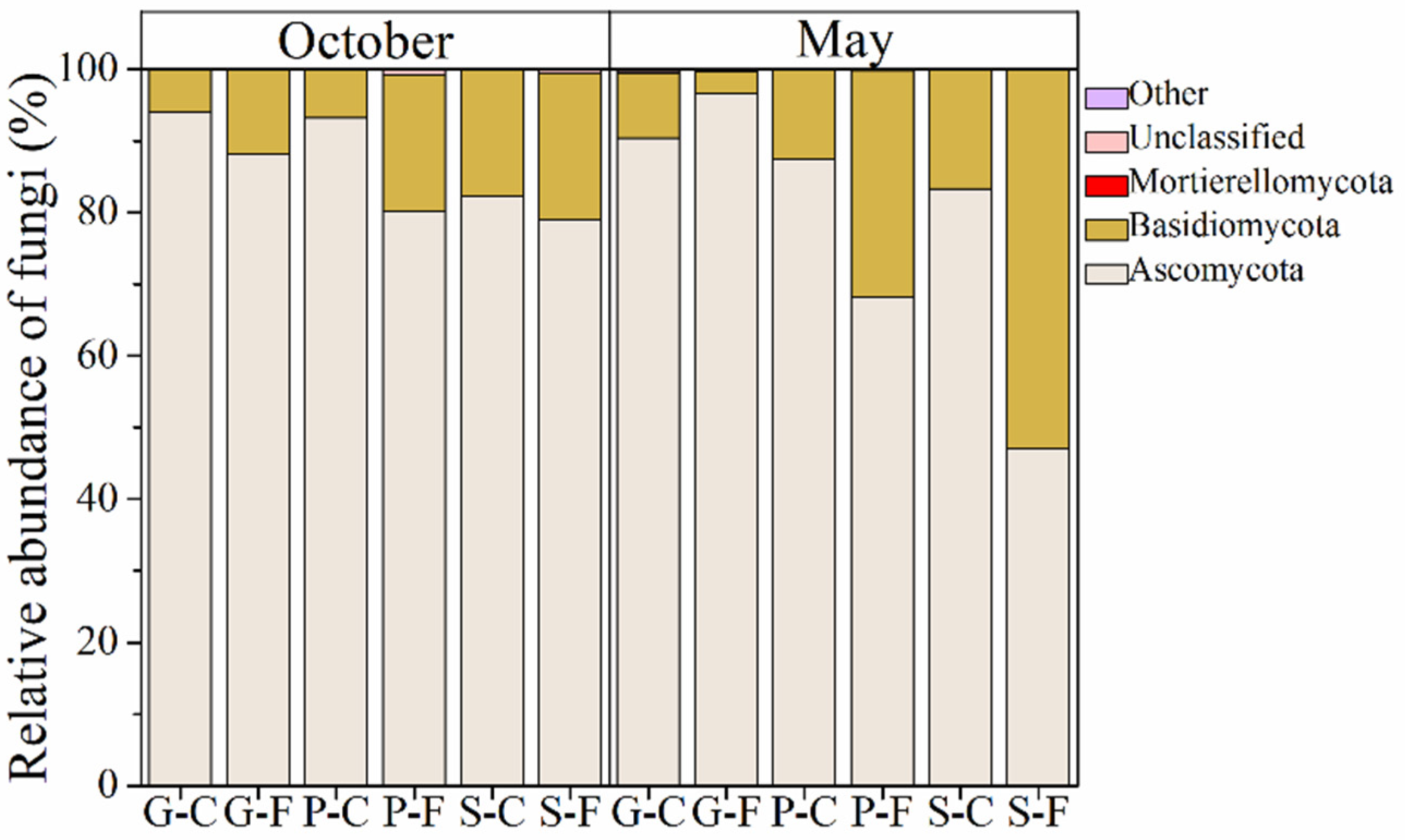
3.2. Variations in Community Composition of Leaf-Associated Fungi during October and May
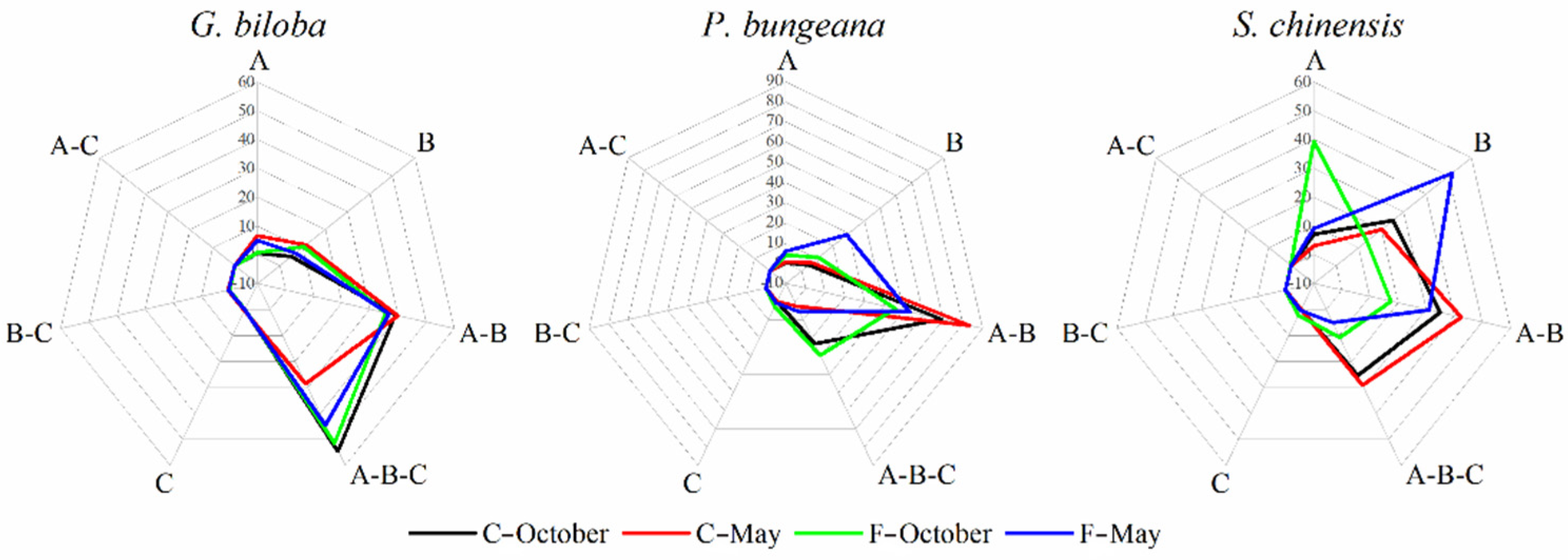
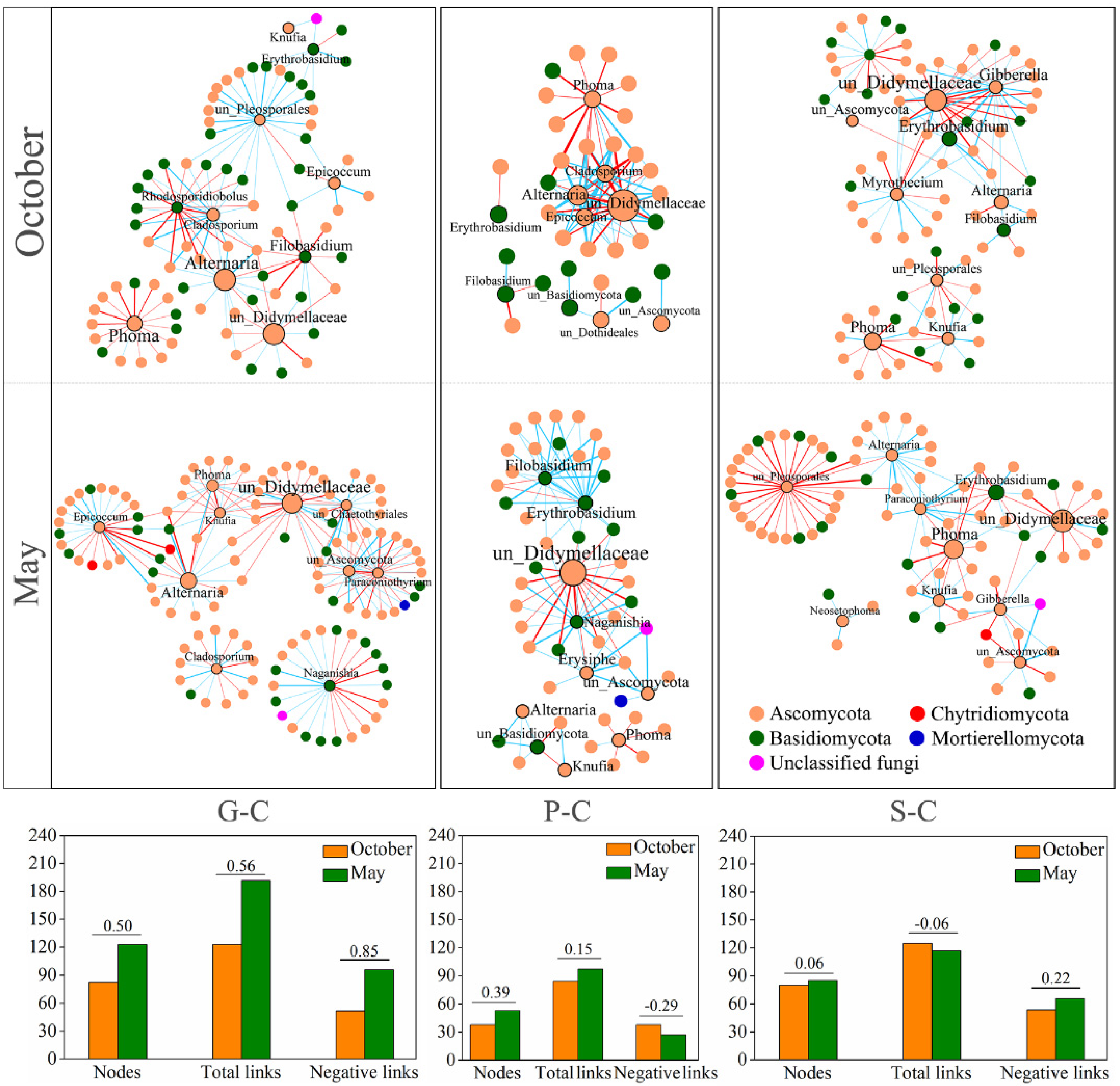
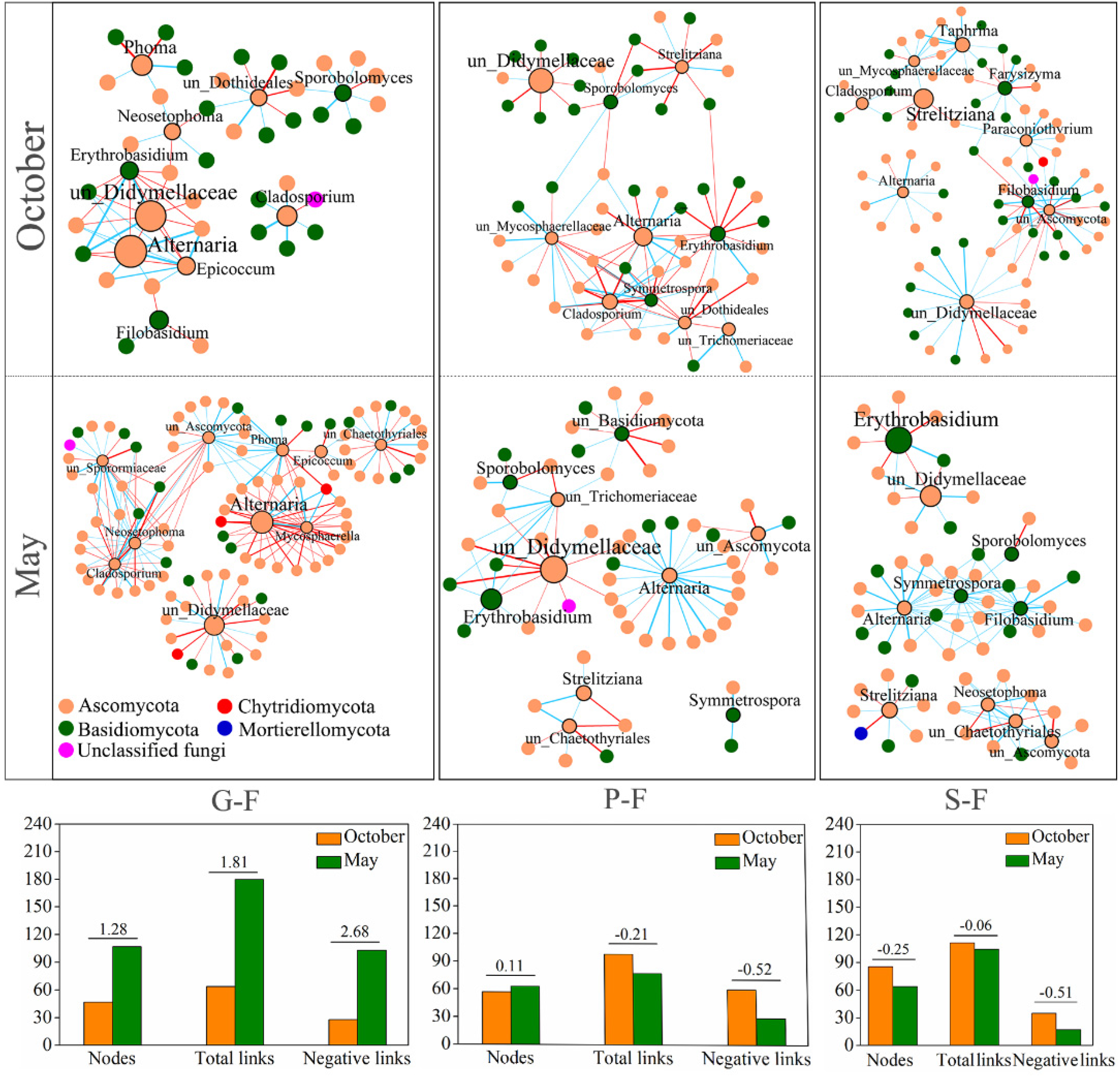
3.3. Variations in Fungal Interactions of Leaf-Associated Fungi during October and May
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Atamna-Ismaeel, N.; Finkel, O.M.; Glaser, F.; Sharon, I.; Schneider, R.; Post, A.F.; Spudich, J.L.; Mering, C.V.; Vorholt, J.A.; Iluz, D. Microbial rhodopsins on leaf surfaces of terrestrial plants. Environ. Microbiol. 2012, 14, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, N.B.; Vitousek, P.M. Fungal endophyte communities reflect environmental structuring across a Hawaiian landscape. Proc. Natl. Acad. Sci. USA 2012, 109, 13022–13027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Castillo, J.A. Influence of Light on Plant-Phyllosphere Interaction. Front. Plant Sci. 2018, 9, 1482. [Google Scholar] [CrossRef] [Green Version]
- Stone, B.W.G.; Weingarten, E.A.; Jackson, C.R. The role of the phyllosphere microbiome in plant health and function. Annu. Plant Rev. Online 2018, 1, 533–556. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Podile, A.R. Biological control of late leaf spot of peanut (Arachis hypogaea) with chitinolytic bacteria. Phytopathology 2005, 95, 1157–1165. [Google Scholar] [CrossRef] [Green Version]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Métraux, J.-P.; L’Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Meckes, N.; Pervaiz, Z.H.; Traw, M.B. Microbial Interactions in the Phyllosphere Increase Plant Performance under Herbivore Biotic Stress. Front. Microbiol. 2017, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.G.; Miller, R.M. Carbon cycling by arbuscular mycorrhizal fungi in soil-plant systems. Trends Plant Sci. 2003, 8, 407–409. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.C.; Jiang, Y.; Li, M.B.; Zhu, W.X.; Xu, N.; Zhang, H.H. Improving plant growth and alleviating photosynthetic inhibition from salt stress using amf in alfalfa seedlings. J. Plant Interact. 2019, 14, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, E.R.; Ballhorn, D.J. Do foliar endophytes matter in litter decomposition? Microorganisms 2020, 8, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of host manipulation by intracellular effectors from plant pathogenic fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef] [PubMed]
- Mahnert, A.; Ortega, R.A.; Berg, C.; Grebe, M.; Berg, G. Leaves of Indoor Ornamentals Are Biodiversity and Functional Hotspots for Fungi. Front. Microbiol. 2018, 9, 2343. [Google Scholar] [CrossRef] [PubMed]
- Bashir, I.; War, A.F.; Rafiq, I.; Reshi, Z.A.; Rashid, I.; Shouche, Y.S. Phyllosphere microbiome: Diversity and functions. Microbiol. Res. 2022, 254, 126888. [Google Scholar] [CrossRef] [PubMed]
- Ryder, L.S.; Talbot, N.J. Regulation of appressorium development in pathogenic fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Sarsaiya, S.; Wu, Q.; Lu, Y.; Shi, J. A review of plant leaf fungal diseases and its environment speciation. Bioengineered 2019, 10, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Khan, M.A.; Sharif, M.; Shah, J.H.; Rehman, M.H.U.; Javed, K. An automated detection and classification of citrus plant diseases using image processing techniques: A review. Comput. Electron. Agric. 2018, 153, 12–32. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks. New Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef] [Green Version]
- Busby, P.E.; Peay, K.G.; Newcombe, G. Common foliar fungi of Populus trichocarpa modify Melampsora rust disease severity. New Phytol. 2016, 209, 1681–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordier, T.; Robin, C.; Capdevielle, X.; Fabreguettes, O.; Desprez-Loustau, M.L.; Vacher, C. The composition of phyllosphere fungal assemblages of European beech (Fagus sylvatica) varies significantly along an elevation gradient. New Phytol. 2012, 196, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Izuno, A.; Kanzaki, M.; Artchawakom, T.; Wachrinrat, C.; Isagi, Y. Vertical Structure of Phyllosphere Fungal Communities in a Tropical Forest in Thailand Uncovered by High-Throughput Sequencing. PLoS ONE 2016, 11, e0166669. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Li, S.C.; Wu, B.W.; Wang, Y.L.; Ji, N.N.; Yao, H.; Cai, H.Y.; Shi, M.M.; Zhang, D.X. Mainland and island populations of mussaenda kwangtungensis differ in their phyllosphere fungal community composition and network structure. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Bowsher, A.W.; Benucci, G.M.N.; Bonito, G.; Shade, A. Seasonal dynamics of core fungi in the switchgrass phyllosphere, and co-occurrence with leaf bacteria. Phytobiomes J. 2021, 5, 60–68. [Google Scholar] [CrossRef]
- Gong, T.Y.; Xin, X.F. Phyllosphere microbiota: Community dynamics and its interaction with plant hosts. J. Integr. Plant Biol. 2021, 63, 297–304. [Google Scholar] [CrossRef]
- Bao, L.J.; Gu, L.K.; Sun, B.; Cai, W.Y.; Zhang, S.W.; Zhuang, G.Q.; Bai, Z.H.; Zhuang, X.L. Seasonal variation of epiphytic bacteria in the phyllosphere of Gingko biloba, Pinus bungeana and Sabina chinensis. FEMS Microbiol. Ecol. 2020, 96, fiaa017. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslov, S.; Sneppen, K. Specificity and stability in topology of protein networks. Science 2002, 296, 910–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Xu, S.; Jiang, C.; Zhang, Y.; Bai, N.; Zhuang, G.; Bai, Z.; Zhuang, X. Impacts of Human Activities on the Composition and Abundance of Sulfate-Reducing and Sulfur-Oxidizing Microorganisms in Polluted River Sediments. Front. Microbiol. 2019, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Salles, J.F.; Poly, F.; Schmid, B.; Le Roux, X. Community niche predicts the functioning of denitrifying bacterial assemblages. Ecology 2009, 90, 3324–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, Q.; Wang, Y.; Xiong, S.; Yu, H.; Zhao, X.; Tan, W.; Cui, D.; Xi, B. Untangling the response of fungal community structure, composition and function in soil aggregate fractions to food waste compost addition. Sci. Total Environ. 2021, 769, 145248. [Google Scholar] [CrossRef]
- Kembel, S.W.; Mueller, R.C. Plant traits and taxonomy drive host associations in tropical phyllosphere fungal communities. Botany 2014, 92, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Sun, X.; He, C.; Maitra, P.; Li, X.C.; Guo, L.D. Phyllosphere epiphytic and endophytic fungal community and network structures differ in a tropical mangrove ecosystem. Microbiome 2019, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Katsoula, A.; Vasileiadis, S.; Karamanoli, K.; Vokou, D.; Karpouzas, D.G. Factors structuring the epiphytic archaeal and fungal communities in a semi-arid mediterranean ecosystem. Microb. Ecol. 2021, 82, 638–651. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and epiphytic phyllosphere fungal communities are shaped by different environmental factors in a mediterranean ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef] [Green Version]
- Haridas, S.; Albert, R.; Binder, M.; Bloem, J.; LaButti, K.; Salamov, A.; Andreopoulos, B.; Baker, S.E.; Barry, K.; Bills, G.; et al. 101 Dothideomycetes genomes: A test case for predicting lifestyles and emergence of pathogens. Stud. Mycol. 2020, 96, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lucking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of dothideomycetes: Dothideomycetidae and pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Qian, X.; Duan, T.T.; Sun, X.; Zheng, Y.; Wang, Y.L.; Hu, M.L.; Yao, H.; Ji, N.N.; Lv, P.P.; Chen, L.; et al. Host genotype strongly influences phyllosphere fungal communities associated with Mussaenda pubescens var. alba (Rubiaceae). Fungal Ecol. 2018, 36, 141–151. [Google Scholar] [CrossRef]
- Xiong, C.; He, J.Z.; Singh, B.K.; Zhu, Y.G.; Wang, J.T.; Li, P.P.; Zhang, Q.B.; Han, L.L.; Shen, J.P.; Ge, A.H.; et al. Rare taxa maintain the stability of crop mycobiomes and ecosystem functions. Environ. Microbiol. 2021, 23, 1907–1924. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.; Ponder, M.M.; Garcia-Diaz, J. Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact. Microorganisms 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, R.M.; Padilla, G.; Araujo, W.L. The biotechnological potential of Epicoccum spp.: Diversity of secondary metabolites. Crit. Rev. Microbiol. 2018, 44, 759–778. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.T.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [Green Version]
- Rosado, A.W.C.; Custodio, F.A.; Pinho, D.B.; Ferreira, A.P.S.; Pereira, O.L. Cladosporium species associated with disease symptoms on passiflora edulis and other crops in brazil, with descriptions of two new species. Phytotaxa 2019, 409, 239–260. [Google Scholar] [CrossRef]
- Imperato, V.; Portillo-Estrada, M.; Saran, A.; Thoonen, A.; Kowalkowski, Ł.; Gawronski, S.W.; Rineau, F.; Vangronsveld, J.; Thijs, S. Exploring the Diversity and Aromatic Hydrocarbon Degrading Potential of Epiphytic Fungi on Hornbeams from Chronically Polluted Areas. J. Fungi 2021, 7, 972. [Google Scholar] [CrossRef]
- Larena, I.; Torres, R.; De Cal, A.; Linan, M.; Melgarejo, P.; Domenichini, P.; Bellini, A.; Mandrin, J.F.; Lichou, J.; de Eribe, X.O.; et al. Biological control of postharvest brown rot (Monilinia spp.) of peaches by field applications of Epicoccum nigrum. Biol. Control 2005, 32, 305–310. [Google Scholar] [CrossRef]
- Favaro, L.C.D.; Sebastianes, F.L.D.; Araujo, W.L. Epicoccum nigrum P16, a Sugarcane Endophyte, Produces Antifungal Compounds and Induces Root Growth. PLoS ONE 2012, 7, e36826. [Google Scholar] [CrossRef] [PubMed]
- Harinder, S.; Kim, H.; Song, H.; Joe, M.; Kim, D.; Bahn, Y.S.; Choi, J.I.; Lim, S. A Novel Radiation-Resistant Strain of Filobasidium sp. Isolated from the West Sea of Korea. J. Microbiol. Biotechnol. 2013, 23, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Bodenhausen, N.; Bortfeld-Miller, M.; Ackermann, M.; Vorholt, J.A. A Synthetic Community Approach Reveals Plant Genotypes Affecting the Phyllosphere Microbiota. PLoS Genet. 2014, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapkota, R.; Knorr, K.; Jorgensen, L.N.; O’Hanlon, K.A.; Nicolaisen, M. Host genotype is an important determinant of the cereal phyllosphere mycobiome. New Phytol. 2015, 207, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Costa, D.; Tavares, R.M.; Baptista, P.; Lino-Neto, T. Olive Fungal Epiphytic Communities Are Affected by Their Maturation Stage. Microorganisms 2022, 10, 376. [Google Scholar] [CrossRef]
- Preto, G.; Martins, F.; Pereira, J.A.; Baptista, P. Fungal community in olive fruits of cultivars with different susceptibilities to anthracnose and selection of isolates to be used as biocontrol agents. Biol. Control 2017, 110, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Taîbi, A.; Rivallan, R.; Broussolle, V.; Pallet, D.; Lortal, S.; Meile, J.C.; Constancias, F. Terroir is the main driver of the epiphytic bacterial and fungal communities of mango carposphere in Reunion Island. Front. Microbiol. 2021, 11, 3531. [Google Scholar] [CrossRef]
- Grady, K.L.; Sorensen, J.W.; Stopnisek, N.; Guittar, J.; Shade, A. Assembly and seasonality of core phyllosphere microbiota on perennial biofuel crops. Nat. Commun. 2019, 10, 4135. [Google Scholar] [CrossRef] [Green Version]
- Kivlin, S.N.; Kazenel, M.R.; Lynn, J.S.; Lee Taylor, D.; Rudgers, J.A. Plant identity influences foliar fungal symbionts more than elevation in the colorado rocky mountains. Microb. Ecol. 2019, 78, 688–698. [Google Scholar] [CrossRef]
- Whipps, J.M.; Hand, P.; Pink, D.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 105, 1744–1755. [Google Scholar] [CrossRef] [Green Version]
- Friesen, M.L.; Porter, S.S.; Stark, S.C.; von Wettberg, E.J.; Sachs, J.L.; Martinez-Romero, E. Microbially Mediated Plant Functional Traits. In Annual Review of Ecology, Evolution, and Systematics; Futuyma, D.J., Shaffer, H.B., Simberloff, D., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2011; Volume 42, pp. 23–46. [Google Scholar] [CrossRef] [Green Version]
- Lamit, L.J.; Busby, P.E.; Lau, M.K.; Compson, Z.G.; Wojtowicz, T.; Keith, A.R.; Zinkgraf, M.S.; Schweitzer, J.A.; Shuster, S.M.; Gehring, C.A.; et al. Tree genotype mediates covariance among communities from microbes to lichens and arthropods. J. Ecol. 2015, 103, 840–850. [Google Scholar] [CrossRef] [Green Version]
- Maignien, L.; DeForce, E.A.; Chafee, M.E.; Eren, A.M.; Simmons, S.L. Ecological Succession and Stochastic Variation in the Assembly of Arabidopsis thaliana Phyllosphere Communities. mBio 2014, 5, e00682-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelp, B.J.; Bown, A.W.; Faure, D. Extracellular gamma-aminobutyrate mediates communication between plants and other organisms. Plant Physiol. 2006, 142, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Kniskern, J.M.; Traw, M.B.; Bergelson, J. Salicylic acid and jasmonic acid signaling defense pathways reduce natural bacterial diversity on Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2007, 20, 1512–1522. [Google Scholar] [CrossRef] [Green Version]
- Horton, M.W.; Bodenhausen, N.; Beilsmith, K.; Meng, D.; Muegge, B.D.; Subramanian, S.; Vetter, M.M.; Vilhjálmsson, B.J.; Nordborg, M.; Gordon, J.I.; et al. Genome-wide association study of Arabidopsis thaliana leaf microbial community. Nat. Commun. 2014, 5, 5320. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Coaker, G.L.; Leveau, J.H. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol. Lett. 2013, 348. [Google Scholar] [CrossRef] [Green Version]
- Osono, T. Endophytic and epiphytic phyllosphere fungi of Camellia japonica: Seasonal and leaf age-dependent variations. Mycologia 2008, 100, 387–391. [Google Scholar] [CrossRef]
- Zambell, C.B.; White, J.F. In the forest vine Smilax rotundifolia, fungal epiphytes show site-wide spatial correlation, while endophytes show evidence of niche partitioning. Fungal Divers 2015, 75, 279–297. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Duran, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175, 973–983. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Sample ID 1 | No. of Reads | No. of OTUs | Coverage (10−2) |
|---|---|---|---|---|
| October | G-C | 66,537 ± 4457 | 244 ± 68 | 99.8 ± 0.07 |
| G-F | 66,992 ± 6242 | 236 ± 21 | 99.8 ± 0.02 | |
| P-C | 67,902 ± 7380 | 159 ± 34 | 99.8 ± 0.04 | |
| P-F | 64,663 ± 4559 | 275 ± 26 | 99.8 ± 0.02 | |
| S-C | 71,866 ± 7404 | 271 ± 60 | 99.7 ± 0.07 | |
| S-F | 58,039 ± 7050 | 266 ± 43 | 99.8 ± 0.05 | |
| May | G-C | 65,209 ± 11,900 | 909 ± 75 | 99.4 ± 0.06 |
| G-F | 68,522 ± 3565 | 757 ± 57 | 99.4 ± 0.05 | |
| P-C | 62,256 ± 8731 | 171 ± 57 | 99.8 ± 0.07 | |
| P-F | 66,604 ± 6499 | 234 ± 47 | 99.8 ± 0.05 | |
| S-C | 71,771 ± 3424 | 360 ± 104 | 99.6 ± 0.10 | |
| S-F | 65,835 ± 13,643 | 254 ± 79 | 99.8 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, L.; Sun, B.; Liu, J.; Zhang, S.; Xu, N.; Zhang, X.; Bohu, T.; Bai, Z. Leaf-Associated Epiphytic Fungi of Gingko biloba, Pinus bungeana and Sabina chinensis Exhibit Delicate Seasonal Variations. J. Fungi 2022, 8, 631. https://doi.org/10.3390/jof8060631
Bao L, Sun B, Liu J, Zhang S, Xu N, Zhang X, Bohu T, Bai Z. Leaf-Associated Epiphytic Fungi of Gingko biloba, Pinus bungeana and Sabina chinensis Exhibit Delicate Seasonal Variations. Journal of Fungi. 2022; 8(6):631. https://doi.org/10.3390/jof8060631
Chicago/Turabian StyleBao, Lijun, Bo Sun, Jiayu Liu, Shiwei Zhang, Nan Xu, Xiaoran Zhang, Tsing Bohu, and Zhihui Bai. 2022. "Leaf-Associated Epiphytic Fungi of Gingko biloba, Pinus bungeana and Sabina chinensis Exhibit Delicate Seasonal Variations" Journal of Fungi 8, no. 6: 631. https://doi.org/10.3390/jof8060631