
New Species of Large-Spored Alternaria in Section Porri Associated with Compositae Plants in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Morphological Observations
2.3. DNA Extraction and PCR Amplification
2.4. Phylogenetic Analyses
3. Results
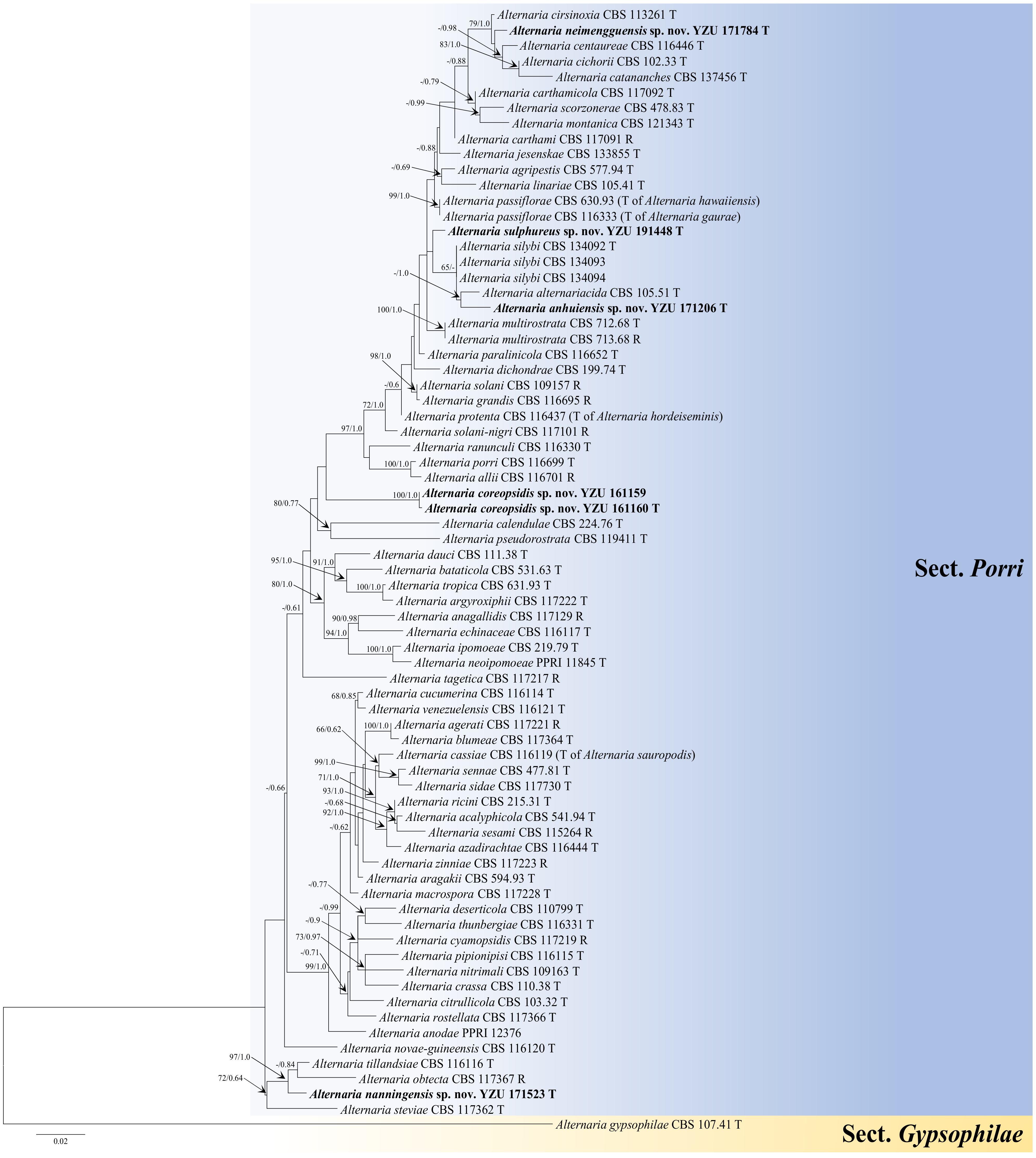
3.1. Phylogenetic Analysis
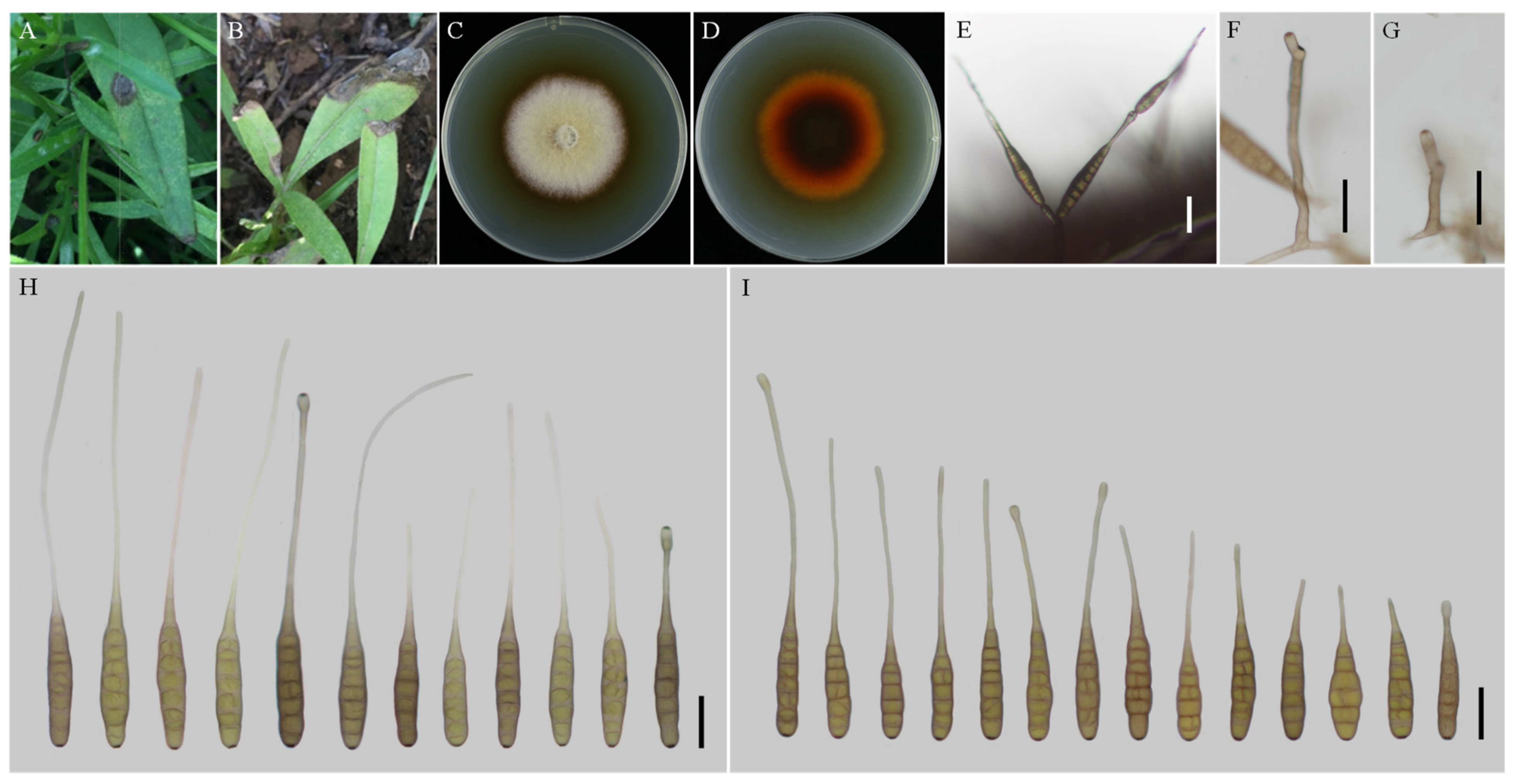
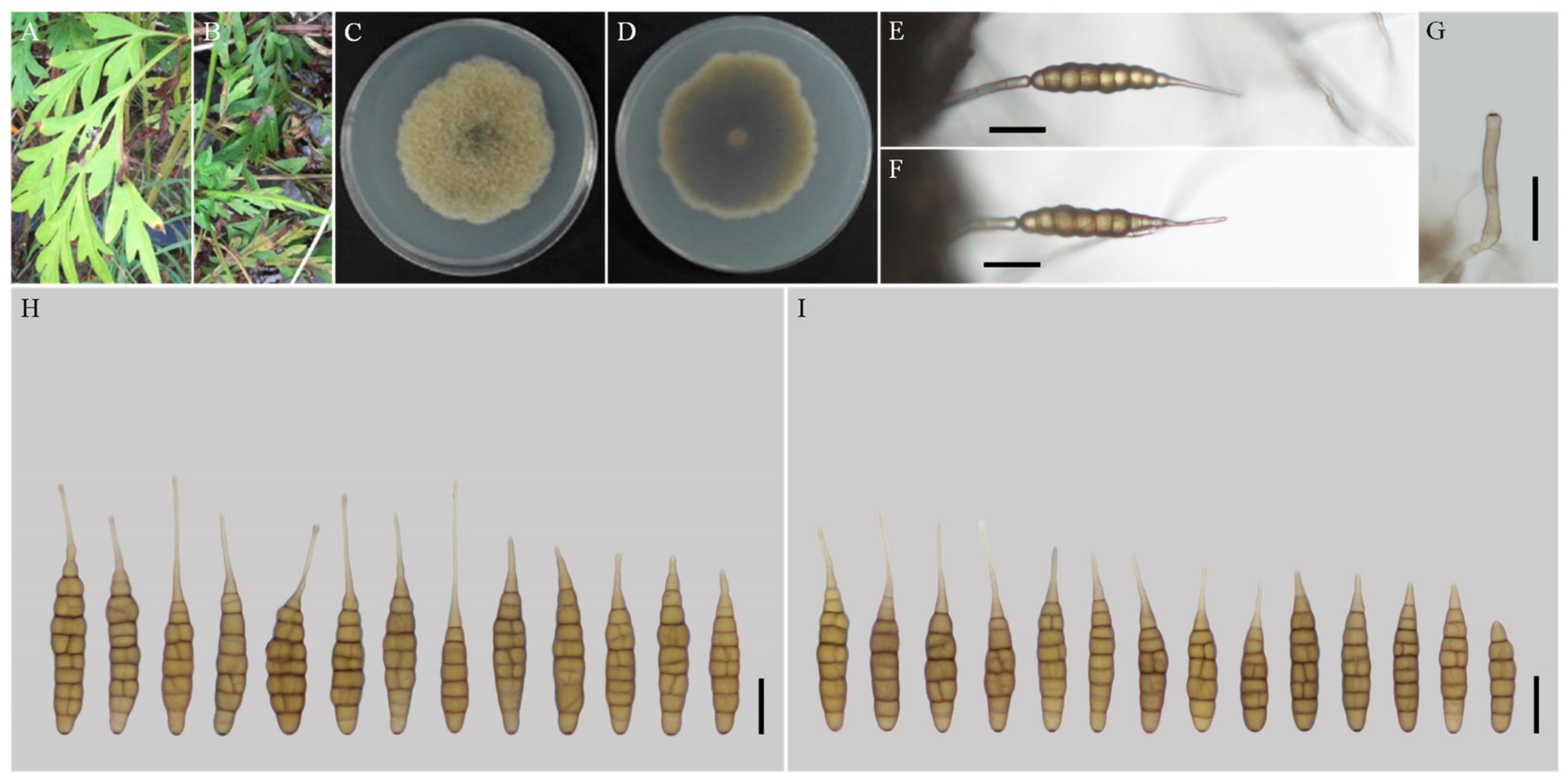
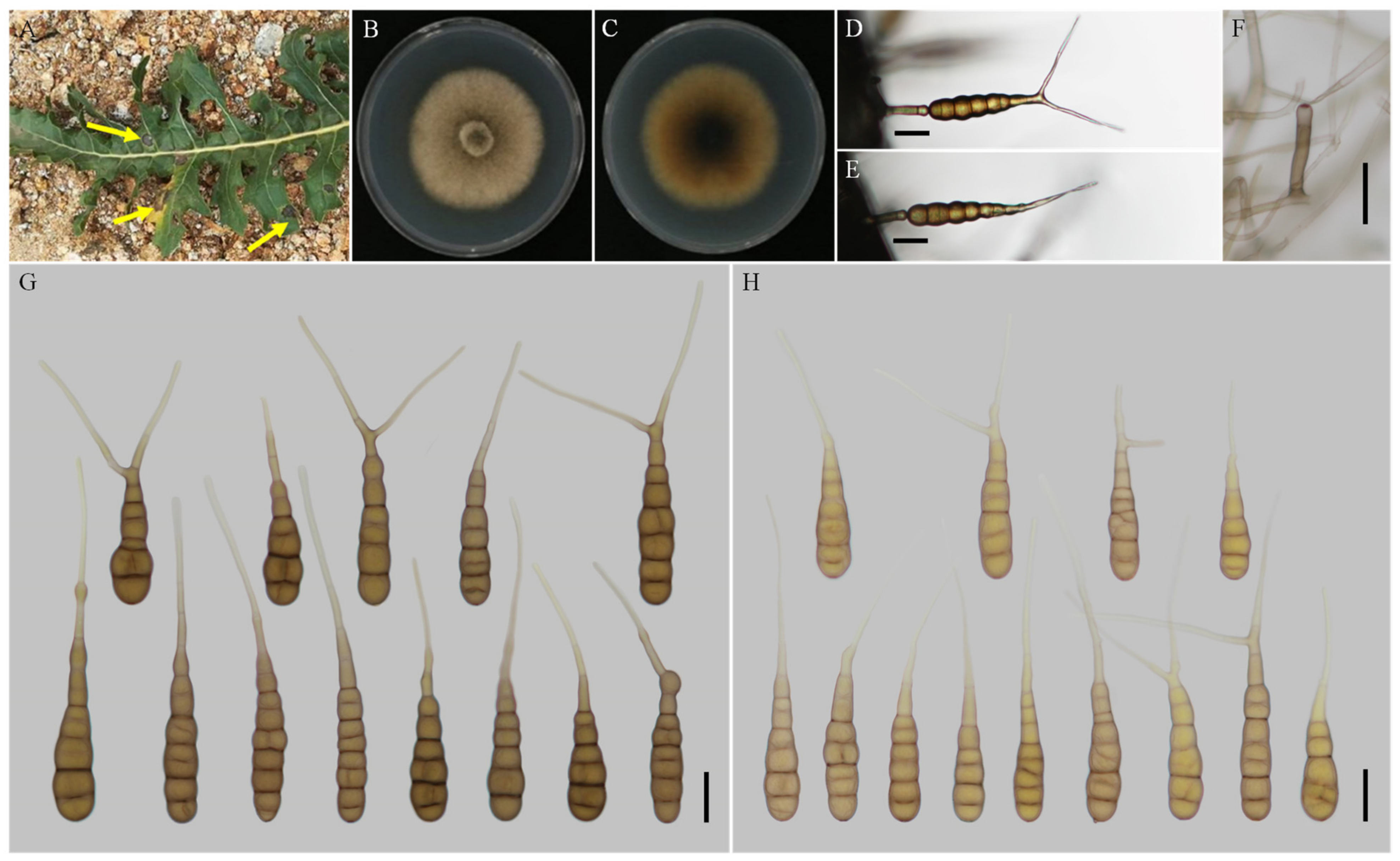
3.2. Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woudenberg, J.H.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, E.G. Alternaria: An Identification Manual; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Zhang, T.Y. Flora Fungorum Sinicorum, Alternaria; Science Press: Beijing, China, 2003; Volume 16. [Google Scholar]
- Woudenberg, J.H.C.; Truter, M.; Groenewald, J.Z.; Crous, P.W. Large-spored Alternaria pathogens in section Porri disentangled. Stud. Mycol. 2014, 79, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Gannibal, P.B. Distribution of Alternaria species among sections. 1. Section Porri. Mycotaxon 2015, 130, 207–213. [Google Scholar] [CrossRef]
- Pinto, V.E.F.; Patriarca, A. Alternaria Species and Their Associated Mycotoxins; Mycotoxigenic Fungi Humana Press: New York, NY, USA, 2017; pp. 13–32. [Google Scholar] [CrossRef]
- Pryor, B.M.; Bigelow, D.M. Molecular characterization of Embellisia and Nimbya species and their relationship to Alternaria, Ulocladium and Stemphylium. Mycologia 2003, 95, 1141–1154. [Google Scholar] [CrossRef]
- Park, M.S.; Romanoski, C.E.; Pryor, B.M. A re-examination of the phylogenetic relationship between the causal agents of carrot black rot, Alternaria radicina and A. carotiincultae. Mycologia 2008, 100, 511–527. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.F.; Liao, J.; Chen, X.Y.; Liu, Q.K.; Yu, Z.H.; Deng, J.X. A novel species and a new record of Alternaria isolated from two Solanaceae plants in China. Mycol. Prog. 2019, 18, 1005–1012. [Google Scholar] [CrossRef]
- Htun, A.A.; Liu, H.F.; He, L.; Zhou, X.Z.; Aung, S.L.L.; Deng, J.X. New species and new record of Alternaria from onion leaf blight in Myanmar. Mycol. Prog. 2022, 21, 59–69. [Google Scholar] [CrossRef]
- Pryor, B.M.; Gilbertson, R.L. Molecular phylogenetic relationships amongst Alternaria species and related fungi based upon analysis of nuclear ITS and mt SSU rDNA sequences. Mycol. Res. 2000, 104, 1312–1321. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.S.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Park, M.S.; Pryor, B.M. Nimbya and embellisia revisited, with nov.comb. for Alternaria celosiae and A. perpunctulata. Mycol. Prog. 2012, 11, 799–815. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: Formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, D.P.; Gannibal, P.B.; Dugan, F.M.; Pryor, B.M. Characterization of Alternaria isolates from the infectoria species-group and a new taxon from Arrhenatherum, Pseudoalternaria arrhenatheria sp. nov. Mycol. Prog. 2014, 13, 257–276. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; Vries, M.D.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Pei, D.F.; Aung, S.L.L.; Liu, H.F.; Liu, Q.K.; Yu, Z.H.; Deng, J.X. Alternaria hydrangeae sp. nov. (Ascomycota: Pleosporaceae) from Hydrangea paniculata in China. Phytotaxa 2019, 401, 287–295. [Google Scholar] [CrossRef]
- He, L.; Cheng, H.; Htun, A.A.; Ge, H.; Xia, Z.Z.; Guo, J.J.; Deng, J.X.; Du, T. Phylogeny and taxonomy of two new Alternaria (Ascomycota: Pleosporaceae) species in section Gypsophilae from China. Mycol. Prog. 2021, 20, 355–363. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and taxonomy of the pleomorphic genus Alternaria. Mycol. Prog. 2016, 15, 3. [Google Scholar] [CrossRef]
- Ghafri, A.A.; Maharachchikunbura, S.S.; Hyde, K.D.; Nadiya, A.A.S.; Abdullah, M.A.S. A new section and a new species of Alternaria encountered from Oman. Phytotaxa 2019, 405, 279–289. [Google Scholar] [CrossRef]
- Gannibal, P.B.; Orina, A.S.; Gasich, E.L. A new section for Alternaria helianthiinficiens found on sunflower and new asteraceous hosts in Russia. Mycol. Prog. 2022, 21, 34. [Google Scholar] [CrossRef]
- Hind, N. Introduction to the compositae, the largest family of flowering plants. Curtis’s Bot. Mag. 2018, 35, 332–338. [Google Scholar] [CrossRef]
- Luo, H.; Yu, Z.; Lu, H.; Zhao, M.; Zhou, Y.; Deng, J. Taxonomy of Alternaria from Compositae: Research progress. Chin. Agric. Sci. Bull. 2018, 34, 63–70. [Google Scholar]
- Mackinnon, S.L.; Keifer, P.; Ayer, W.A. Components from the phytotoxic extract of Alternaria brassicicola, a black spot pathogen of canola. Phytochemistry 1999, 51, 215–221. [Google Scholar] [CrossRef]
- Andersen, B.; Krøger, E.; Roberts, R.G. Chemical and morphological segregation of Alternaria alternata, A. gaisen and A. longipes. Mycol. Res. 2001, 105, 291–299. [Google Scholar] [CrossRef]
- Thomma, B.P. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.X.; Li, M.J.; Paul, N.C.; Oo, M.M.; Lee, H.B.; Oh, S.K.; Yu, S.H. Alternaria brassicifolii sp. nov. isolated from Brassica rapa subsp. pekinensis in Korea. Mycobiology 2018, 46, 172–176. [Google Scholar] [CrossRef] [Green Version]
- Cenis, J.L. Rapid extraction of fungal DNA for PCR amplification. Nucleic Acids Res. 1992, 20, 2380. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi–gene phylogeny of Clavicipitaceae (Ascomycota, fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenetics Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Chun, J. Computer Assisted Classification and Identification of Actinomycetes. Ph.D. Thesis, Unversity of Newcastle, Newcastle upon Tyne, UK, 1995. Available online: http://theses.ncl.ac.uk/jspui/handle/10443/410 (accessed on 1 May 2022).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A. FigTree v.1.3.1. Institute of Evolutionary Biology; University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Gannibal, P.B. Taxonomic studies of Alternaria from Russia: New species on Asteraceae. Mycotaxon 2010, 114, 109–114. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Luo, H.; Xia, Z.Z.; Chen, Y.Y.; Zhou, Y.; Deng, J.X. Morphology and Molecular Characterization of Alternaria argyranthemi on Chrysanthemum coronarium in China. Mycobiology 2018, 46, 278–282. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Liu, H.F.; Htun, A.A.; Ge, H.; Deng, J.X.; Du, T. First Report of Alternaria cinerariae Causing Leaf Spot on Tussilago farfara in China. Plant Dis. 2020, 104, 3264. [Google Scholar] [CrossRef]
- Lee, Y.H.; Cho, W.D.; Kim, W.K.; Jin, K.S.; Lee, E.J. Report on host-unrecorded diseases identified from economical crops in Korea. Res. Rep. Rural Developm. Admin. 1991, 33, 15–19. [Google Scholar]
- Zhao, G.; Zhang, T.Y.; Cao, A.X.; Wang, H.K. Phylogenetic relationships of Alternaria and related genera and taxonomic status of A. leucanthemi inferred from ITS rDNA sequence analysis. Mycosystema 2006, 25, 184–191. [Google Scholar] [CrossRef]
- Tomioka, K.; Sato, T.; Koganezawa, H. Marigold leaf spot caused by Alternaria tagetica new to Japan. J. Gen. Plant Pathol. 2000, 664, 294–298. [Google Scholar] [CrossRef]
- Li, M.J.; Deng, J.X.; Paul, N.C.; Lee, H.B.; Yu, S.H. Characterization and pathogenicity of Alternaria vanuatuensis, a new record from Allium plants in Korea and China. Mycobiology 2014, 42, 412–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, N.C.; Deng, J.X.; Lee, H.B.; Yu, S.H. Characterization and pathogenicity of Alternaria burnsii from seeds of Cucurbita maxima (Cucurbitaceae) in Bangladesh. Mycobiology 2015, 43, 384–391. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section | Species | Strain | Locality | Substrate | ITS | GAPDH | Alt a 1 | TEF1 | RPB2 |
|---|---|---|---|---|---|---|---|---|---|
| Porri | A. acalyphicola | CBS 541.94 T | Seychelles | Acalypha indica | KJ718097 | KJ717952 | KJ718617 | KJ718446 | KJ718271 |
| Porri | A. agerati | CBS 117221 R | USA | Ageratum houstonianum | KJ718098 | KJ717953 | KJ718618 | KJ718447 | KJ718272 |
| Porri | A. agripestis | CBS 577.94 T | Canada | Euphorbia esula, stem lesion | KJ718099 | JQ646356 | KJ718619 | KJ718448 | KJ718273 |
| Porri | A. allii | CBS 116701 R | USA | Allium cepa var. viviparum | KJ718103 | KJ717957 | KJ718623 | KJ718452 | KJ718277 |
| Porri | A. alternariacida | CBS 105.51 T | UK | Solanum lycopersicum, fruit | KJ718105 | KJ717959 | KJ718625 | KJ718454 | KJ718279 |
| Porri | A. anagallidis | CBS 117129 R | New Zealand | Anagallis arvensis, leaf spot | KJ718109 | KJ717962 | KJ718629 | KJ718457 | KJ718283 |
| Porri | A. anhuiensis sp. nov. | YZU 171206 T | China | Coreopsis basalis, leaf | MK264916 | MK303949 | MK303953 | MK303958 | MK303960 |
| Porri | A. anodae | PPRI 12376 | South Africa | Anoda cristata, leaf | KJ718110 | KJ717963 | KJ718630 | KJ718458 | KJ718284 |
| Porri | A. aragakii | CBS 594.93 T | USA | Passiflora edulis | KJ718111 | KJ717964 | KJ718631 | KJ718459 | KJ718285 |
| Porri | A. argyroxiphii | CBS 117222 T | USA | Argyroxiphium sp. | KJ718112 | JQ646350 | KJ718632 | KJ718460 | KJ718286 |
| Porri | A. azadirachtae | CBS 116444 T | Australia | Azadirachta indica, leaf spot | KJ718115 | KJ717967 | KJ718635 | KJ718463 | KJ718289 |
| Porri | A. bataticola | CBS 531.63 T | Japan | Ipomoea batatas | KJ718117 | JQ646349 | JQ646433 | KJ718465 | KJ718291 |
| Porri | A. blumeae | CBS 117364 T | Thailand | Blumea aurita | KJ718126 | AY562405 | AY563291 | KJ718474 | KJ718300 |
| Porri | A. calendulae | CBS 224.76 T | Germany | Calendula officinalis | KJ718127 | KJ717977 | KJ718648 | KJ718475 | KJ718301 |
| Porri | A. calendulae | CBS 101498 | New Zealand | Calendula officinalis, leaf | KJ718128 | KJ717978 | KJ718645 | KJ718476 | KJ718302 |
| Porri | CBS 116439 T | New Zealand | Rosa sp., leaf spot | KJ718129 | KJ717979 | KJ718646 | KJ718477 | KJ718303 | |
| Porri | CBS 116650 R | Japan | Calendula officinalis, leaf spot | KJ718130 | KJ717980 | KJ718647 | KJ718478 | KJ718304 | |
| Porri | A. carthami | CBS 117091 R | USA | Carthamus tinctorius, leaf spot | KJ718133 | KJ717983 | KJ718651 | KJ718481 | KJ718307 |
| Porri | A. carthamicola | CBS 117092 T | Iraq | Carthamus tinctorius | KJ718134 | KJ717984 | KJ718652 | KJ718482 | KJ718308 |
| Porri | A. cassiae | CBS 116119 T | Malaysia | Sauropus androgynus | KJ718136 | KJ717986 | KJ718654 | KJ718484 | KJ718310 |
| Porri | A. catananches | CBS 137456 T | Netherlands | Catananche caerulea | KJ718139 | KJ717989 | KJ718657 | KJ718487 | KJ718313 |
| Porri | A. centaureae | CBS 116446 T | USA | Centaurea solstitialis, leaf spot | KJ718140 | KJ717990 | KJ718658 | KJ718488 | KJ718314 |
| Porri | A. cichorii | CBS 102.33 T | Cyprus | Cichorium intybus, leaf spot | KJ718141 | KJ717991 | KJ718659 | KJ718489 | KJ718315 |
| Porri | A. cirsinoxia | CBS 113261 T | Canada | Cirsium arvense, stem lesion | KJ718143 | KJ717993 | KJ718661 | KJ718491 | KJ718317 |
| Porri | A. citrullicola | CBS 103.32 T | Cyprus | Citrullus vulgaris, fruit | KJ718144 | KJ717994 | KJ718662 | KJ718492 | KJ718318 |
| Porri | A. coreopsidis sp. nov. | YZU 161159 | China | Coreopsis basalis, leaf | MK264914 | MK303947 | MK303951 | MK303955 | MK303971 |
| Porri | YZU 161160 T | China | Coreopsis basalis, leaf | ON130144 | ON229924 | ON229926 | ON229928 | ON229930 | |
| Porri | A. crassa | CBS 110.38 T | Cyprus | Datura stramonium, leaf spot | KJ718147 | KJ717997 | KJ718665 | KJ718495 | KJ718320 |
| Porri | CBS 122590 R | USA | Datura stramonium, leaf spot | KJ718152 | GQ180072 | GQ180088 | KJ718500 | KJ718325 | |
| Porri | A. cucumerina | CBS 116114 T | USA | Luffa acutangula | KJ718153 | KJ718000 | KJ718668 | KJ718501 | KJ718326 |
| Porri | CBS 117225 R | USA | Cucumis melo, leaf spot | KJ718154 | KJ718001 | KJ718669 | KJ718502 | KJ718327 | |
| Porri | A. cyamopsidis | CBS 117219 R | USA | Cyamopsis tetragonoloba, leaf spot | KJ718157 | KJ718004 | KJ718672 | KJ718505 | KJ718330 |
| Porri | A. dauci | CBS 111.38 T | Italy | Daucus carota, seed | KJ718158 | KJ718005 | KJ718673 | KJ718506 | KJ718331 |
| Porri | A. deserticola | CBS 110799 T | Namibia | desert soil | KJ718249 | KJ718077 | KJ718755 | KJ718595 | KJ718424 |
| Porri | A. dichondrae | CBS 199.74 T | Italy | Dichondra repens, leaf spot | KJ718166 | JQ646357 | JQ646441 | KJ718514 | KJ718339 |
| Porri | A. echinaceae | CBS 116117 T | New Zealand | Echinacea sp., leaf lesion | KJ718170 | KJ718015 | KJ718684 | KJ718518 | KJ718343 |
| Porri | A. grandis | CBS 116695 R | USA | Solanum tuberosum, leaf spot | KJ718241 | KJ718070 | KJ718748 | KJ718587 | KJ718416 |
| Porri | A. ipomoeae | CBS 219.79 T | Ethiopia | Ipomoea batatas, stem and petiole | KJ718175 | KJ718020 | KJ718689 | KJ718523 | KJ718348 |
| Porri | A. jesenskae | CBS 133855 T | Slovakia | Fumana procumbens, seed | KJ718177 | KJ718022 | KJ718691 | KJ718525 | KJ718350 |
| Porri | A. linariae | CBS 105.41 T | Denmark | Linaria maroccana, seedling | KJ718180 | KJ718024 | KJ718692 | KJ718528 | KJ718353 |
| Porri | A. passiflorae | CBS 630.93 T | USA | Passiflora edulis | KJ718210 | JQ646352 | KJ718718 | KJ718556 | KJ718383 |
| Porri | CBS 116333 T | New Zealand | Gaura lindheimeri, leaf | KJ718211 | KJ718046 | KJ718719 | KJ718557 | KJ718384 | |
| Porri | A. pipionipisi | CBS 116115 T | India | Cajanus cajan, seed | KJ718214 | KJ718049 | KJ718722 | KJ718560 | KJ718387 |
| Porri | A. porri | CBS 116699 T | USA | Allium cepa, leaf spot | KJ718218 | KJ718053 | KJ718727 | KJ718564 | KJ718391 |
| Porri | A. protenta | CBS 116437 T | New Zealand | Hordeum vulgare, seed | KJ718220 | KJ718055 | KJ718729 | KJ718566 | KJ718393 |
| Porri | A. pseudorostrata | CBS 119411 T | USA | Euphorbia pulcherrima | JN383483 | AY562406 | AY563295 | KC584680 | KC584422 |
| Porri | A. ranunculi | CBS 116330 T | Israel | Ranunculus asiaticus, seed | KJ718225 | KJ718058 | KJ718732 | KJ718571 | KJ718398 |
| Porri | A. ricini | CBS 215.31 T | Japan | Ricinus communis | KJ718226 | KJ718059 | KJ718733 | KJ718572 | KJ718399 |
| Porri | A. rostellata | CBS 117366 T | USA | Euphorbia pulcherrima, leaf | KJ718229 | JQ646332 | KJ718736 | KJ718575 | KJ718402 |
| Porri | A. scorzonerae | CBS 478.83 T | Netherlands | Scorzonera hispanica, leaf spot | KJ718191 | JQ646334 | KJ718699 | KJ718538 | KJ718364 |
| Porri | A. sennae | CBS 477.81 T | India | Senna corymbosa, leaf | KJ718230 | JQ646344 | JQ646428 | EU130543 | KJ718403 |
| Porri | A. sesami | CBS 115264 R | India | Sesamum indicum, seedling | JF780939 | KJ718061 | KJ718738 | KJ718577 | KJ718405 |
| Porri | A. sidae | CBS 117730 T | Kiribati | Sida fallax, leaf spot | KJ718232 | KJ718062 | KJ718739 | KJ718578 | KJ718406 |
| Porri | A. silybi | CBS 134092 T | Russia | Silybum marianum, leaf | KJ718233 | KJ718063 | KJ718740 | KJ718579 | KJ718407 |
| Porri | CBS 134093 | Russia | Silybum marianum, leaf | KJ718234 | KJ718064 | KJ718741 | KJ718580 | KJ718408 | |
| Porri | CBS 134094 | Russia | Silybum marianum, leaf | KJ718235 | KJ718065 | KJ718742 | KJ718581 | KJ718409 | |
| Porri | A. solani | CBS 109157 R | USA | Solanum tuberosum, leaf spot | KJ718238 | GQ180080 | KJ718746 | KJ718585 | KJ718413 |
| Porri | A. solani-nigri | CBS 117101 R | New Zealand | Solanum nigrum, leaf spot | KJ718247 | KJ718075 | KJ718753 | KJ718593 | KJ718422 |
| Porri | A. steviae | CBS 117362 T | Japan | Stevia rebaudiana, leaf spot | KJ718252 | KJ718079 | KJ718758 | KJ718598 | KJ718427 |
| Porri | A. tagetica | CBS 117217 R | USA | Tagetes sp., leaf spot | KJ718256 | KJ718083 | KJ718763 | KJ718602 | KJ718431 |
| Porri | CBS 297.79 | UK | Tagetes sp., seed | KJ718253 | KJ718080 | KJ718759 | KJ718599 | KJ718428 | |
| Porri | CBS 298.79 | UK | Tagetes sp., seed | KJ718254 | KJ718081 | KJ718760 | KJ718600 | KJ718429 | |
| Porri | CBS 479.81 R | UK | Tagetes erecta, seed | KC584221 | KC584143 | KJ718761 | KC584692 | KC584434 | |
| Porri | CBS 480.81 R | USA | Tagetes sp., seed | KJ718255 | KJ718082 | KJ718762 | KJ718601 | KJ718430 | |
| Porri | A. thunbergiae | CBS 116331 T | Australia | Thunbergia alata, leaf spot | KJ718257 | KJ718084 | KJ718764 | KJ718603 | KJ718432 |
| Porri | A. tillandsiae | CBS 116116 T | New Zealand | Tillandsia usneoides | KJ718260 | KJ718087 | KJ718767 | KJ718606 | KJ718435 |
| Porri | A. tropica | CBS 631.93 T | USA | Passiflora edulis, fruit | KJ718261 | KJ718088 | KJ718768 | KJ718607 | KJ718436 |
| Porri | A. venezuelensis | CBS 116121 T | Venezuela | Phaseolus vulgaris, leaf spot | KJ718263 | KJ718263 | KJ718770 | KJ718609 | KJ718438 |
| Porri | A. zinniae | CBS 117223 R | New Zealand | Zinnia elegans, leaf spot | KJ718270 | KJ718096 | KJ718777 | KJ718616 | KJ718445 |
| Porri | CBS 118.44 | Hungary | Callistephus chinensis, seed | KJ718264 | JQ646361 | KJ718771 | KJ718610 | KJ718439 | |
| Porri | CBS 117.59 | Italy | Zinnia elegans | KJ718266 | KJ718092 | KJ718773 | KJ718612 | KJ718441 | |
| Porri | CBS 299.79 | UK | Zinnia sp., seed | KJ718268 | KJ718094 | KJ718775 | KJ718614 | KJ718443 | |
| Gypsophilae | A. gypsophilae | CBS 107.41 T | Netherlands | Gypsophila elegans, seed | KC584199 | KC584118 | KJ718688 | KC584660 | KC584401 |
| Species | Strain | Conidia | Sporulation Pattern | Medium | Reference | |||
|---|---|---|---|---|---|---|---|---|
| Shape | Size (μm) | Transversesepta | Beak (μm) | |||||
| A. alternariacida | CBS 105.51 | Smooth-walled, narrowly ovoid; smooth-walled | (85–) 99–111 (–121) × (6–) 7–8 (–10) | (3–) 5–6 (–8) | (47–) 129–257 (–610) × 2 | Solitary or in unbranched chains of 2 (–3) conidia | SNA | [4] |
| A. anhuiensis sp. nov. | YZU 171206 | Long-narrow ovoid or ellipsoid; smooth-walled | 61–100 (–111.5) × (11.5–) 13–19.5 | 6–11 | (22–) 58–133 (–150.5) × 2.5–4 (–4.5) | Solitary | V8A | This study |
| A. catananches | CBS 137456 | Narrowly ovoid; ornamented in lower half of the conidium | (26–) 37–43 (–57) × (7–) 8–9 (–11) | (2–) 4 (–6) | (77–) 126–160 (–260) × 2 | Solitary | SNA | [4] |
| A. centaureae | CBS 116446 | Long narrow-ellipsoid or long-ovoid; ornamentation and punctate to pustulate | 75–100 × 15–24 | 7–10 | 140–190 × 1.5–6 | Solitary | V8A | [2] |
| A. cichorii | CBS 117218 | Narrow-ovoid or narrow-ellipsoid; smooth-walled | 60–80 × 14–18 | 7–12 | 120–240 × 2.5–7 | Terminal clumps of 4–5 conidia | V8A | [2] |
| A. cirsinoxia | CBS 113261 | Long-obclavate, short-ovoid; punctulate-walled | 70–90 × 12–22 | 7–9 | 80–165 × 2.5–4 | Solitary or tufts of 2–7 conidia | V8A | [2] |
| A. coreopsidis sp. nov. | YZU 161160 | Long-narrow ovoid or ellipsoid; smooth-walled | (48.5–) 55–80 (–85) × (9–) 10–15 | 6–9 | (20–) 30–140 (–206) × (2–) 2.5–4 | Solitary or 2–conidium chains | V8A | This study |
| A. nanningensis sp. nov. | YZU 171523 | Ovoid or ellipsoid; smooth-walled | (40.5–) 47–79 (–87) × 9–13.5 (–15) | 6–10 (–11) | 10–30 × (1–) 1.5–2 (–3) | Solitary | V8A | This study |
| A. neimengguensis sp. nov. | YZU 171784 | Ovoid or ellipsoid; smooth-walled | (70–) 77–130 (–143.5) × (13–) 15–20 (–23) | 6–11 (–12) | (24.5–) 35–65 (–92) × (1.5–) 2–3 (–4) | Solitary | V8A | This study |
| A. obtecta | CBS 134278 | Long-ovoid or ellipsoid; smooth or punctulate-walled | 65–95 × 18–22 | 7–10 | 55–150 × 2 | Solitary | PCA | [2] |
| A. porri | CBS 116698 | Ovoid, sometimes broad or nearly cylindrical; smooth or punctulate-walled | 70–105 × 19–24 | 8–12 | 95–160 × 2–6.5 | Solitary | V8A | [2] |
| A. silybi | CBS 134093 | Long-ellipsoid, subcylindrical or long-ovoid | 50–80 × 15–20 (–22) | (5–) 7–10 | 70–130 (–190) × 3 | Solitary | V4A | [40] |
| A. steviae | CBS 117362 | Long-ovoid, subellipsoid, or obovoid; smooth or punctulate-walled | 55–95 × 18–30 | 7–10 | 60–120 × 1.5–2.5 | Solitary or tiny distal clumps | V8A | [2] |
| A. sulphureus sp. nov. | YZU 191448 | Ovoid, ellipsoid, or obovoid; smooth-walled | (64–) 74–116 × (12.5–) 14–20 (–25.5) | (5–) 7–11 | (25.5–) 34–151 (–159.5) × 2.5–4.5 (–5.5) | Solitary | V8A | This study |
| A. tillandsiae | CBS 116116 | Long-ovoid, ellipsoid, long-obovoid; smooth or a minor punctulate-walled | 70–102 × 16–19 | 8–11 | 75–120 × 2 | Solitary | V8A | [2] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Luo, H.; Cheng, H.; Gou, Y.-N.; Yu, Z.-H.; Deng, J.-X. New Species of Large-Spored Alternaria in Section Porri Associated with Compositae Plants in China. J. Fungi 2022, 8, 607. https://doi.org/10.3390/jof8060607
Zhao L, Luo H, Cheng H, Gou Y-N, Yu Z-H, Deng J-X. New Species of Large-Spored Alternaria in Section Porri Associated with Compositae Plants in China. Journal of Fungi. 2022; 8(6):607. https://doi.org/10.3390/jof8060607
Chicago/Turabian StyleZhao, Lin, Huan Luo, Hong Cheng, Ya-Nan Gou, Zhi-He Yu, and Jian-Xin Deng. 2022. "New Species of Large-Spored Alternaria in Section Porri Associated with Compositae Plants in China" Journal of Fungi 8, no. 6: 607. https://doi.org/10.3390/jof8060607