
Three Small Cysteine-Free Proteins (CFP1–3) Are Required for Insect-Pathogenic Lifestyle of Metarhizium robertsii


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recognition and Sequence Analysis of CFP Homologs in M. robertsii
2.2. Generation of Targeted Gene Mutants
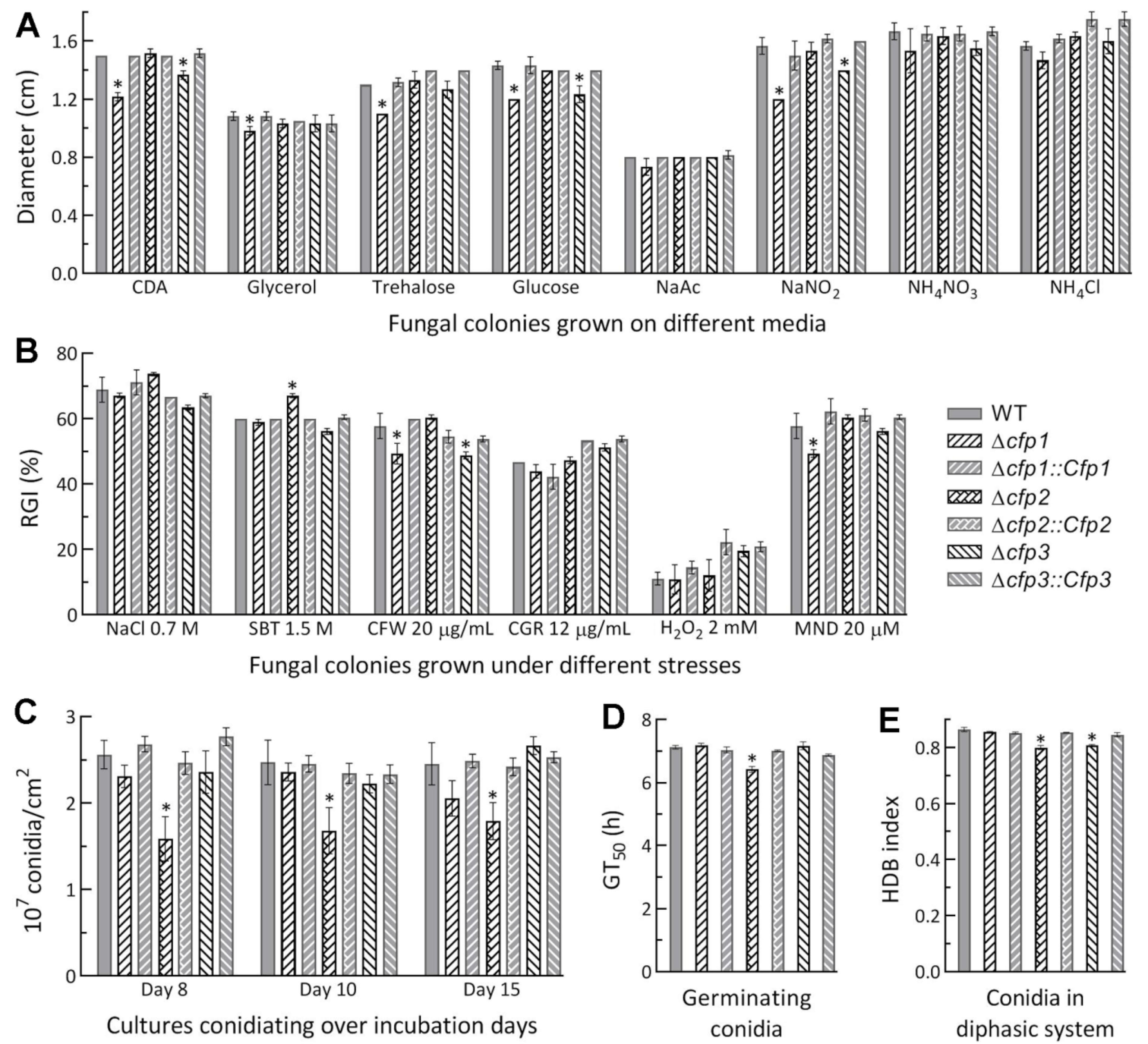
2.3. Assessments of Growth Rates under Different Culture Conditions
2.4. Assessments of Conidial Yield and Quality
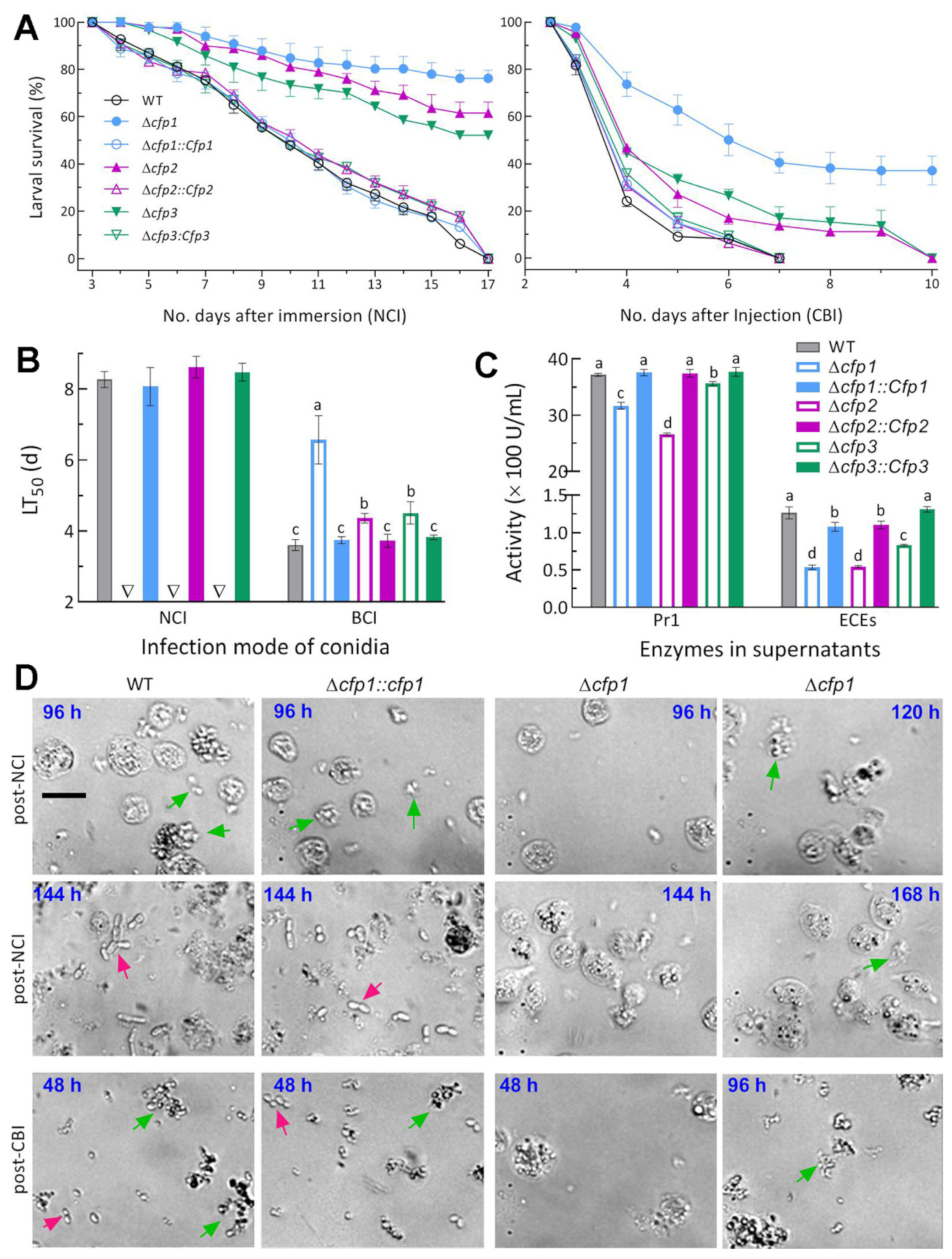
2.5. Fungal Virulence Bioassays
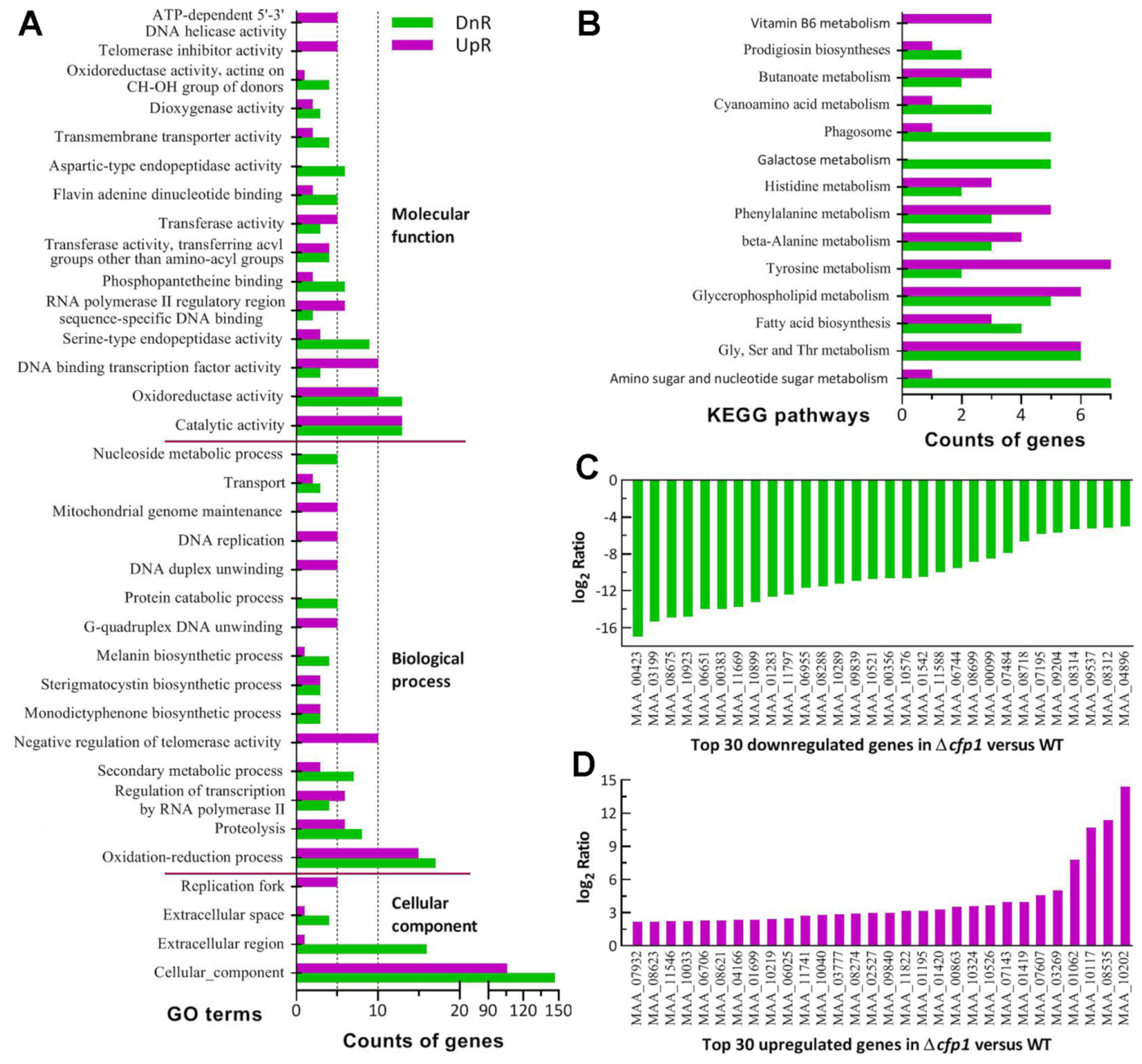
2.6. Transcriptomic Analysis
3. Results
3.1. Sequence Features of CFP1–3
3.2. Limited Roles of cfp1–3 in Asexual Cycle In Vitro
3.3. Indispensable Roles of cfp1–3 in Insect Pathogenicity and Virulence
3.4. Transcriptomic Insight into Indispensable Role of cfp1 in Infection Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Faria, M.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Luo, Z.B.; Keyhani, N.O. Improving mycoinsecticides for insect biological control. Appl. Microbiol. Biotechnol. 2015, 99, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Wang, S.B. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Gao, Q.; Jin, K.; Ying, S.H.; Zhang, Y.J.; Xiao, G.H.; Shang, Y.F.; Duan, Z.B.; Hu, X.; Xie, X.Q.; Zhou, G.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.H.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.W.; Xie, X.Q.; Shang, Y.F.; Zheng, H.J.; Zhou, Y.; St Leger, R.J.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Xiao, G.H.; Zheng, P.; Shang, Y.F.; Su, Y.; Zhang, X.Y.; Liu, X.Z.; Zhan, S.; St Leger, R.J.; Wang, C. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.F.; Xiao, G.H.; Zheng, P.; Cen, K.; Zhan, S.; Wang, C.S. Divergent and convergent evolution of fungal pathogenicity. Genome Biol. Evol. 2016, 8, 1374–1387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.B.; Feng, M.G. Antioxidant enzymes and their contributions to biological control potential of fungal insect pathogens. Appl. Microbiol. Biotechnol. 2018, 102, 4995–5004. [Google Scholar] [CrossRef]
- Ying, S.H.; Feng, M.G. Insight into vital role of autophagy in sustaining biological control potential of fungal pathogens against pest insects and nematodes. Virulence 2019, 10, 429–437. [Google Scholar] [CrossRef]
- Tong, S.M.; Feng, M.G. Insights into regulatory roles of MAPK-cascaded pathways in multiple stress responses and life cycles of insect and nematode mycopathogens. Appl. Microbiol. Biotechnol. 2019, 103, 577–587. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Fuxa, J.R.; Lacey, L.A.; Onstad, D.W.; Kaya, H.K. Definitions of pathogenicity and virulence in invertebrate pathology. J. Invertebr. Pathol. 2005, 88, 1–7. [Google Scholar] [CrossRef]
- Holder, D.J.; Kirkland, B.H.; Lewis, M.W.; Keyhani, N.O. Surface characteristics of the entomopathogenic fungus Beauveria (Cordyceps) bassiana. Microbiology 2007, 153, 3448–3457. [Google Scholar] [CrossRef] [Green Version]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Gao, B.J.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. Subtilisin-like Pr1 proteases marking evolution of pathogenicity in a wide-spectrum insect-pathogenic fungus. Virulence 2020, 11, 365–380. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.L.; Du, Y.R.; Jin, K.; Xia, Y.X. The Ste12-like transcription factor MaSte12 is involved in pathogenicity by regulating the appressorium formation in the entomopathogenic fungus, Metarhizium acridum. Appl. Microbiol. Biotechnol. 2017, 101, 8571–8584. [Google Scholar] [CrossRef]
- Shang, J.M.; Shang, Y.F.; Tang, G.R.; Wang, C.S. Identification of a key G-protein coupled receptor in mediating appressorium formation and fungal virulence against insects. Sci. China Life. Sci. 2021, 64, 466–477. [Google Scholar] [CrossRef]
- Zhou, Q.; Yu, L.; Ying, S.H.; Feng, M.G. Comparative roles of three adhesin genes (adh1–3) in insect-pathogenic lifecycle of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2021, 105, 5491–5502. [Google Scholar] [CrossRef]
- Lewis, M.W.; Robalino, I.V.; Keyhani, N.O. Uptake of the fluorescent probe FM4-64 by hyphae and haemolymph-derived in vivo hyphal bodies of the entomopathogenic fungus Beauveria bassiana. Microbiology 2009, 155, 3110–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Mas5, a homologue of bacterial DnaJ, is indispensable for the host infection and environmental adaptation of a filamentous fungal insect pathogen. Environ. Microbiol. 2016, 18, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.T.; Peng, H.; Tong, S.M.; Ying, S.H.; Feng, M.G. Distinctive role of fluG in the adaptation of Beauveria bassiana to insect- pathogenic lifecycle and environmental stresses. Environ. Microbiol. 2021, 23, 5184–5199. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal effector proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, M.S.; Carvunis, A.R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Toruño, T.T.; Stergiopoulos, I.; Coaker, G. Plant-pathogen effectors: Cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [Green Version]
- De Wit, P.J.G.M.; Laugé, R.; Honee, G.; Joosten, M.H.A.J.; Vossen, P.; Kooman-Gersmann, M.; Vogelsang, R.; Vervoort, J.J.M. Molecular and biochemical basis of the interaction between tomato and its fungal pathogen Cladosporium fulvum. Antonie Van Leeuwenhoek 1997, 71, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; van Esse, H.P.; Crous, P.W.; de Wit, P.J.G.M. Cladosporium fulvum (syn. Passalora fulva), a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae. Mol. Plant Pathol. 2005, 6, 379–393. [Google Scholar]
- Laugé, R.; Goodwin, P.H.; de Wit, P.J.G.M.; Joosten, M.H.A.J. Specific HR-associated recognition of secreted proteins from Cladosporium fulvum occurs in both host and non-host plants. Plant J. 2000, 23, 735–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kock, M.J.D.; Brandwagt, B.F.; Bonnema, G.; de Wit, P.J.G.M.; Lindhout, P. The tomato orion locus comprises a unique class of Hcr9 genes. Mol. Breed. 2005, 15, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Chu, Z.J.; Wang, Y.J.; Ying, S.H.; Wang, X.W.; Feng, M.G. Genome-wide host-pathogen interaction unveiled by transcriptomic response of diamondback moth to fungal infection. PLoS ONE 2016, 11, e0152908. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.J.; Sun, H.H.; Zhu, X.G.; Ying, S.H.; Feng, M.G. Discovery of a new intravacuolar protein required for the autophagy, development and virulence of Beauveria bassiana. Environ. Microbiol. 2017, 19, 2806–2818. [Google Scholar] [CrossRef]
- Chu, Z.J.; Sun, H.H.; Ying, S.H.; Feng, M.G. Vital role for cyclophilin B (CypB) in asexual development, dimorphic transition and virulence of Beauveria bassiana. Fungal Genet. Biol. 2017, 105, 8–15. [Google Scholar] [CrossRef]
- Zhu, X.G.; Chu, Z.J.; Ying, S.H.; Feng, M.G. Lysyl-tRNA synthetase (Krs) acts a virulence factor of Beauveria bassiana by its vital role in conidial germination and dimorphic transition. Fungal Biol. 2017, 121, 956–965. [Google Scholar] [CrossRef]
- Shao, W.; Cai, Q.; Tong, S.M.; Ying, S.H.; Feng, M.G. Rei1-like protein regulates nutritional metabolism and transport required for the asexual cycle in vitro and in vivo of a fungal insect pathogen. Environ. Microbiol. 2019, 21, 2772–2786. [Google Scholar] [CrossRef]
- Shao, W.; Cai, Q.; Tong, S.M.; Ying, S.H.; Feng, M.G. Nuclear Ssr4 is required for the in vitro and in vivo asexual cycles and global gene activity of Beauveria bassiana. mSystems 2020, 5, e00677-19. [Google Scholar] [CrossRef] [Green Version]
- Mou, Y.N.; Fu, B.; Ren, K.; Tong, S.M.; Ying, S.H.; Feng, M.G. A small cysteine-free protein acts as a novel regulator of fungal insect-pathogenic lifecycle and genomic expression. mSystems 2021, 6, e00098-21. [Google Scholar] [CrossRef]
- Mou, Y.N.; Ren, K.; Tong, S.M.; Ying, S.H.; Feng, M.G. Fungal insecticidal activity elevated by non-risky markerless overexpression of an endogenous cysteine-free protein gene. Pest Manag. Sci. 2022, 78. [Google Scholar] [CrossRef]
- Liu, Q.; Ying, S.H.; Li, J.G.; Tian, C.G.; Feng, M.G. Insight into the transcriptional regulation of Msn2 required for conidiation, multi-stress responses and virulence of two entomopathogenic fungi. Fungal Genet. Biol. 2013, 54, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.M.; Feng, M.G. Phenotypic and molecular insights into heat tolerance of formulated cells as active ingredients of fungal insecticides. Appl. Microbiol. Biotechnol. 2020, 104, 5711–5724. [Google Scholar] [CrossRef]
- Tong, S.M.; Feng, M.G. Molecular basis and regulatory mechanisms underlying fungal insecticides’ resistance to solar ultraviolet irradiation. Pest Manag. Sci. 2022, 78, 30–42. [Google Scholar] [CrossRef]
- Hassan, A.H.; Prochasson, P.; Neely, K.E.; Galasinski, S.C.; Chandy, M.; Carrozza, M.J.; Workman, J.L. Function and selectivity of bromodomains in anchoring chromatin-modifying complexes to promoter nucleosomes. Cell 2002, 111, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Martens, J.A.; Winston, F. Recent advances in understanding chromatin remodeling by Swi/Snf complexes. Curr. Opin. Genet. Dev. 2003, 13, 136–142. [Google Scholar] [CrossRef]
- Lebreton, A.; Saveanu, C.; Decourty, L. Rain, J.C.; Jacquier, A.; Fromont-Racine, M. A functional network involved in the recycling of nucleocytoplasmic pre-60S factors. J. Cell Biol. 2006, 173, 349–360. [Google Scholar] [CrossRef]
- Hung, N.J.; Lo, K.Y.; Patel, S.S.; Helmke, K.; Johnson, A.W. Arx1 is a nuclear export receptor for the 60S ribosomal subunit in yeast. Mol. Biol. Cell 2007, 19, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Greber, B.J.; Gerhardy, S.; Leitner, A.; Leibundgut, M.; Salem, M.; Boehringer, D.; Leulliot, N.; Aebersold, R.; Panse, V.; Ban, N. Insertion of the biogenesis factor Rei1 probes the ribosomal tunnel during 60S maturation. Cell 2016, 164, 91–102. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mou, Y.-N.; Ren, K.; Xu, S.-Y.; Ying, S.-H.; Feng, M.-G. Three Small Cysteine-Free Proteins (CFP1–3) Are Required for Insect-Pathogenic Lifestyle of Metarhizium robertsii. J. Fungi 2022, 8, 606. https://doi.org/10.3390/jof8060606
Mou Y-N, Ren K, Xu S-Y, Ying S-H, Feng M-G. Three Small Cysteine-Free Proteins (CFP1–3) Are Required for Insect-Pathogenic Lifestyle of Metarhizium robertsii. Journal of Fungi. 2022; 8(6):606. https://doi.org/10.3390/jof8060606
Chicago/Turabian StyleMou, Ya-Ni, Kang Ren, Si-Yuan Xu, Sheng-Hua Ying, and Ming-Guang Feng. 2022. "Three Small Cysteine-Free Proteins (CFP1–3) Are Required for Insect-Pathogenic Lifestyle of Metarhizium robertsii" Journal of Fungi 8, no. 6: 606. https://doi.org/10.3390/jof8060606