
Three New Species of Microdochium (Sordariomycetes, Amphisphaeriales) on Miscanthus sinensis and Phragmites australis from Hainan, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphology
2.2. DNA Extraction and Amplification
2.3. Phylogenetic Analyses
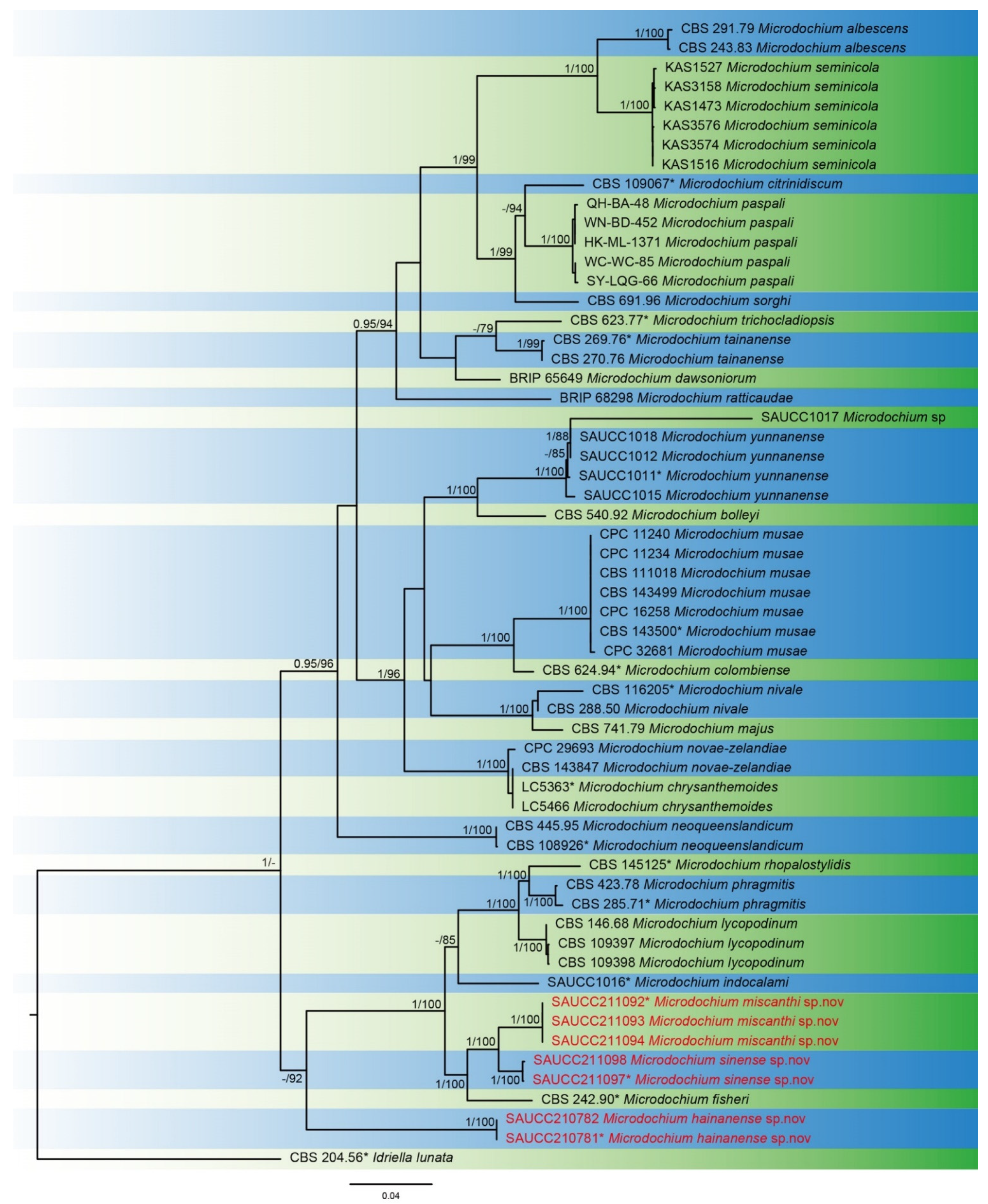
3. Results
3.1. Phylogenetic Analyses
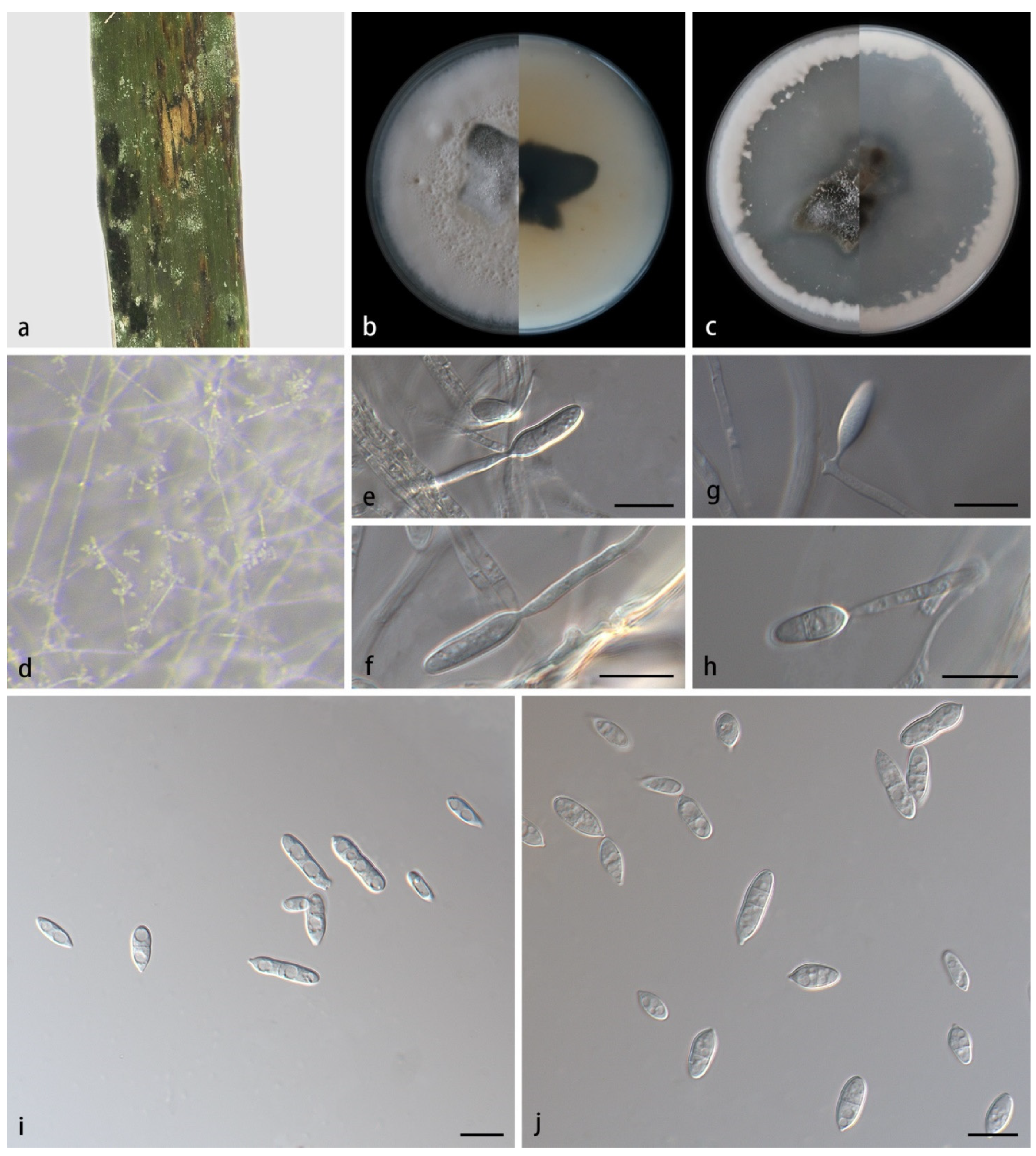
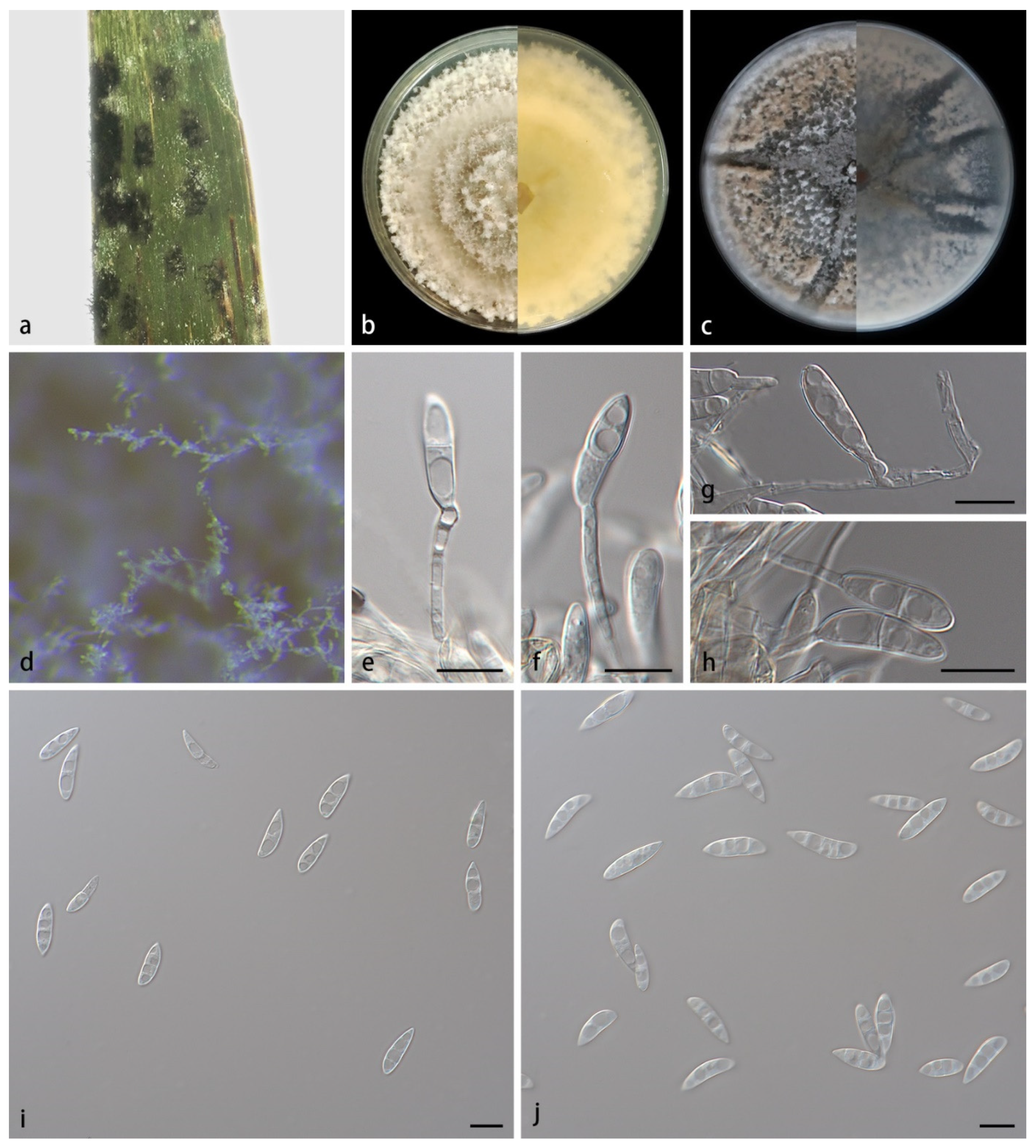
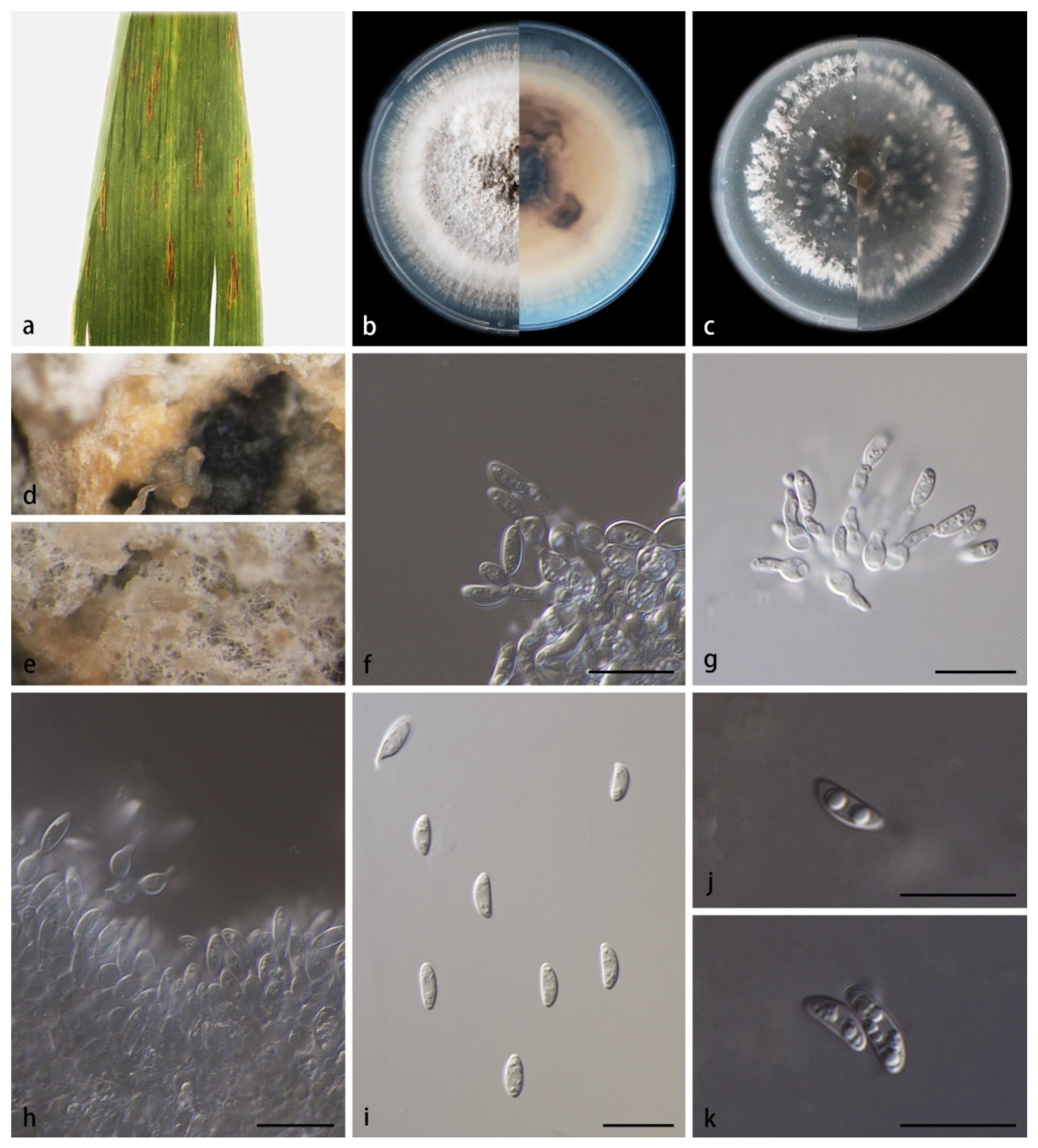
3.2. Taxonomy
3.3. Key to the Species of Microdochium
- 1. Sexual morph known------------------------------------------------------------------------------------2
- 1. Sexual morph unknown-------------------------------------------------------------------------------13
- 2. Perithecia maximum diameter > 200 μm------------------------------------------------------------3
- 2. Perithecia maximum diameter < 200 μm------------------------------------------------------------9
- 3. Maximum number of septa of ascospores > 3----------------------------------------------------- 4
- 3. Maximum number of septa of ascospores ≤ 3------------------------------------------------------5
- 4. Asci size 90.0–120.0 × 21.0–25.0 μm-------------------------------------------------M. consociatum
- 4. Asci size 80.0–100.0 × 17.0–22.0 μm--------------------------------------------------------M. musae
- 5. Asci size = 50.0–70.0 × 7.0–9.0 μm---------------------------------------------------------------------6
- 5. Asci size ≠ 50.0–70.0 × 7.0–9.0 μm---------------------------------------------------------------------7
- 6. Ascospores size 9.5–17.0 × 3.0–4.5 μm-----------------------------------------------------M. majus
- 6. Ascospores size 10.0–17.0 × 3.5–4.5 μm----------------------------------------------------M. nivale
- 7. Ascospores 1–3 septa-------------------------------------------------------------------------------------8
- 7. Ascospores 1–2 septa--------------------------------------------------------------------M. stevensonii
- 8. Ascospores size 20.0–32.0 × 3.0–3.5 μm------------------------------------------M. fusariisporum
- 8. Ascospores size 15.0–25.0 × 4.0–5.0 μm----------------------------------------------M. passiflorae
- 9. Perithecia maximum diameter < 150 μm----------------------------------------------------------10
- 9. Perithecia maximum diameter > 150 μm----------------------------------------------------------11
- 10. Ascospores size 20.0–22.0 × 3.5 μm----------------------------------------------------M. opuntiae
- 10. Ascospores size 12.0–22.0 × 3.0–5.0 μm---------------------------------------------M. seminicola
- 11. Chlamydospores known--------------------------------------------------------------M. ratticaudae
- 11. Chlamydospores unknown--------------------------------------------------------------------------12
- 12. Conidia falcate, 11.0–16.0 × 3.5–4.5 μm, 0–3 septa---------------------------------M. albescens
- 12. Conidia lunate, 8.0–15.0 × 2.5–3.5 μm, 0–1 septa------------------------------M. lycopodinum
- 13. Chlamydospores known-----------------------------------------------------------------------------14
- 13. Chlamydospores unknown--------------------------------------------------------------------------16
- 14. Conidia oblong---------------------------------------------------------------------M. trichocladiopsis
- 14. Conidia lunate------------------------------------------------------------------------------------------15
- 15. Chlamydospores chain or clusters--------------------------------------------------------M. bolleyi
- 15. Chlamydospores rounded or obovoid-----------------------------------------------------M. poae
- 16. Conidia aseptate----------------------------------------------------------------------------------------17
- 16. Conidia septate-----------------------------------------------------------------------------------------23
- 17. Conidiogenous cells two types-----------------------------------------------------M. yunnanense
- 17. Conidiogenous cells one type-----------------------------------------------------------------------18
- 18. Conidiogenous cells with denticulate-------------------------------------------------------------19
- 18. Conidiogenous cells not denticulate--------------------------------------------------------------21
- 19. Conidiogenous cells ampulliform------------------------------------------------------------------20
- 19. Conidiogenous cells cylindrical----------------------------------------------------M. sclerotiorum
- 20. Conidia pointed at both ends, no appendages--------------------------------------M. griseum
- 20. Conidia with straight appendages at both ends---------------------------M. queenslandicum
- 21. Conidiogenous cells monoblastic--------------------------------------M. hainanense sp. nov.
- 21. Conidiogenous cells sympodial--------------------------------------------------------------------22
- 22. Conidia filiform, 7.0–16.0 × 1.0 μm----------------------------------------------------M. palmicola
- 22. Conidia lunate, 7.5.0–11.0 × 1.8–2.0 μm--------------------------------------M. queenslandicum
- 23. Conidiogenous cells two types------------------------------------------------------M. colombiense
- 23. Conidiogenous cells one type-----------------------------------------------------------------------24
- 24. Conidia relatively narrow, acicular, filiform, falcate or lunate-----------------------------25
- 24. Conidia relatively rounded, ellipsoid, fusiform, cylindrical or obovoid-----------------32
- 25. Conidia with long appendages at both ends------------------------------------------M. linariae
- 25. Conidia without appendages at both ends------------------------------------------------------26
- 26. Conidia with conspicuous rhachides-----------------------------------------------M. tainanense
- 26. Conidia without conspicuous rhachides---------------------------------------------------------27
- 27. Conidiogenous cells ampulliform------------------------------------------------------------------28
- 27. Conidiogenous cells cylindrical--------------------------------------------------------------------31
- 28. Maximum number of septa of conidia = 10---------------------------------------------M. sorghi
- 28. Maximum number of septa of conidia < 10------------------------------------------------------29
- 29. Conidia lunate-----------------------------------------------------------------M. neoqueenslandicum
- 29. Conidia falcate------------------------------------------------------------------------------------------30
- 30. Conidia size 25.0–30.0 × 1.5–2.0 μm, 0–1 septa----------------------------------M. caespitosum
- 30. Conidia size 7.0– 20.5 ×2.5–4.5 μm, 0–3 septa-----------------------------------------M. paspali
- 31. Conidia size 25.0–75.0 × 1.0–2.0 μm, 0–3 septa--------------------------------M. dawsoniorum
- 31. Conidia size 5.5–10.0 × 2.0–2.5 μm, 0–1 septa-------------------------------M. novae-zelandiae
- 32. Conidia with guttulate--------------------------------------------------------------------------------33
- 32. Conidia no guttulate----------------------------------------------------------------------------------35
- 33. Conidiogenous cells solitary-------------------------------------------------M. chrysanthemoides
- 33. Conidiogenous cells sympodial--------------------------------------------------------------------34
- 34. Conidia size 10.0–14.5 × 2.0–3.0 μm, 0–1 septa-----------------------------------M. phragmitis
- 34. Conidia size 13.0–23.0 × 2.5–4.0 μm, 1–3 septa------------------------------M. rhopalostylidis
- 35. Conidia cylindrical-------------------------------------------------------------------------------------36
- 35. Conidia fusiform---------------------------------------------------------------------------------------42
- 36. Conidiogenous cells denticulate--------------------------------------------------------------------37
- 36. Conidiogenous cells not denticulate--------------------------------------------------------------40
- 37. Conidiogenous cells blastic-sympodial-------------------------------------------M. cylindricum
- 37. Conidiogenous cells mono- or polyblastic------------------------------------------------------38
- 38. Conidia spindle-to-rod-shaped-------------------------------------------M. miscanthi sp. nov.
- 38. Conidia clavate to obovoid--------------------------------------------------------------------------39
- 39. Conidia size 7.0–31.0 × 2.0–3.0 μm, 0–3 septa----------------------------------M. citrinidiscum
- 39. Conidia size 13.0–15.5 × 3.5–5.5 μm, 1–3 septa-----------------------------------M. indocalami
- 40. Conidiogenous cells ampulliform------------------------------------------------------M. maydis
- 40. Conidiogenous cells cylindrical--------------------------------------------------------------------41
- 41. Conidiogenous cells monoblastic, 16.3–22.4 × 4.1–5.7 μm-------------M. sinense sp. nov.
- 41. Conidiogenous cells sympodial, 6.5–15.0 × 2.5–3.5 μm-----------------------------M. stoveri
- 42. Conidiogenous cells ampulliform------------------------------------------------------------------43
- 42. Conidiogenous cells cylindrical--------------------------------------------------------------------44
- 43. Conidiogenous cells solitary------------------------------------------------------------M. punctum
- 43. Conidiogenous cells sympodial-------------------------------------------------------M. triticicola
- 44. Conidiogenous cells mono- or polyblastic---------------------------------------M. maculosum
- 44. Conidiogenous cells sympodial--------------------------------------------------------------------45
- 45. Conidiogenous cells not denticulate------------------------------------------M. panattonianum
- 45. Conidiogenous cells denticulate--------------------------------------------------------------------46
- 46. Conidia size 7.0–12.0 × 3.0–4.0 μm, 0–1 septa------------------------------------------M. fisheri
- 46. Conidia size 8.0–15.0 × 3.0–4.5 μm, 1–2 septa----------------------------------M. intermedium
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sydow, H. Mycotheca germanic. Fasc. XLII–XLV (No. 2051–2250). Annls Mycol. 1924, 22, 257–268. [Google Scholar]
- Hernández-Restrepo, M.; Groenewald, J.Z.; Crous, P.W. Taxonomic and phylogenetic re-evaluation of Microdochium, Monographella and Idriella. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 57–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable Mycobiota from karst caves in China, with descriptions of 20 new species. Pers. Mol. Phylogeny Evol. Fungi 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Akulov, A.; Denman, S.; Roux, J.; Braun, U.; Burgess, T.I.; Carnegie, A.J.; Váczy, K.Z. New and interesting fungi 1. Fuse 2018, 1, 169–215. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W. New and interesting fungi 2. Fuse 2019, 3, 57–134. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Huang, S.T.; Xia, J.W.; Zhang, X.G.; Sun, W.X.; Li, Z. Two new species of Microdochium from Indocalamus longiauritus in south-western China. MycoKeys 2020, 72, 93–108. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Pers. Mol. Phylogeny Evol. Fungi 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/ (accessed on 27 March 2022).
- Bánki, O.; Roskov, Y.; Döring, M.; Ower, G.; Vandepitte, L.; Hobern, D.; Remsen, D.; Schalk, P.; DeWalt, R.E.; Keping, M.; et al. Catalogue of Life Checklist (Version 2022-03-21). Cat. Life 2022. [Google Scholar] [CrossRef]
- Glynn, N.C.; Hare, M.C.; Parry, D.W.; Edwards, S.G. Phylogenetic analysis of EF-1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties Majus and Nivale to species status. Fungal Biol. 2005, 109, 872–880. [Google Scholar] [CrossRef]
- Jewell, L.E.; Hsiang, T. Multigene differences between Microdochium nivale and Microdochium majus. Botany 2013, 91, 99–106. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W. Fungi in agricultural soils. J. Appl. Ecol. 1972, 160. [Google Scholar] [CrossRef]
- Mouchacca, J.; Samson, R.A. Deux nouvelles espèces du genre Microdochium Sydow. Rev. Mycol. 1973, 37, 267–275. [Google Scholar]
- Sutton, B.C. Eucalyptus microfungi: Microdochium and Phaeoisaria species from Brazil. Nova Hedwig. 1976, 27, 215–222. [Google Scholar]
- Von, A.J.A. Notes on Monographella and Microdochium. Trans. Br. Mycol. Soc. 1984, 83, 373–374. [Google Scholar]
- Harris, D.C. Microdochium fusarioides sp.nov. from oospores of Phytophthora syringae. Trans. Br. Mycol. Soc. 1985, 84, 358–361. [Google Scholar] [CrossRef]
- Lombard, L.; Van, D.M.N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.M.; Li, G.S.; Zhao, M.Q.; Cai, L. A new leaf blight disease of turfgrasses caused by Microdochium poae, sp. nov. Mycologia 2019, 111, 265–273. [Google Scholar] [CrossRef]
- Von Arx, J.A. Plant pathogenic fungi. Nova Hedwig. 1987, 87, 1–288. [Google Scholar]
- Hiruma, K.; Kobae, Y.; Toju, H. Beneficial associations between Brassicaceae plants and fungal endophytes under nutrient-limiting conditions: Evolutionary origins and host–symbiont molecular mechanisms. Curr. Opin. Plant Biol. 2018, 44, 145–154. [Google Scholar] [CrossRef]
- Mandyam, K.G.; Roe, J.; Jumpponen, A. Arabidopsis thaliana model system reveals a continuum of responses to root endophyte colonization. Fungal Biol. 2013, 117, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, K.; Loughin, T.; Jumpponen, A. Isolation and morphological and metabolic characterization of common endophytes in annually burned tallgrass prairie. Mycologia 2010, 102, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadmani, L.; Jamali, S.; Fatemi, A. Biocontrol activity of endophytic fungus of barley, Microdochium bolleyi, against Gaeumannomyces graminis var. tritici. Mycol. Iran. 2018, 5, 7–14. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M. The sooty moulds. Fungal Divers 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and Phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from Fagaceae Leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in the Netherlands. MycoKeys 2021, 85, 31–56. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.; Waller, J.; Abang, M.M.; Zhang, Z.J.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers 2009, 39, 183–204. [Google Scholar]
- Crous, P.W.; Verkleij, G.J.M.; Groenewald, J.Z.; Houbraken, J. Fungal Biodiversity; CBS Laboratory Manuals Series 1; CBS-KNAW Fungal Biodiversity Centre Utrecht: Utrecht, The Netherlands, 2009. [Google Scholar]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; Lee, S.H.; Taylor, L.; Shawe-Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.; Tedersoo, L.; Abarenkov, K.; Ryberg, M.; Kristiansson, E.; Hartmann, M.; Schoch, C.; Nylander, J.; Bergsten, J.; Porter, T.; et al. Five simple guidelines for establishing basic authenticity and reliability of newly generated fungal ITS sequences. MycoKeys 2012, 4, 37–63. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Phylogenet. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment. Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: San Diego, CA, USA, 2012; p. 8. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van, D.M.P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModelTest v. 2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Parkinson, V.O.; Sivanesan, A.; Booth, C. The perfect state of the rice leaf-scald fungus and the taxonomy of both the perfect and imperfect states. Trans. Br. Mycol. Soc. 1981, 76, 59–69. [Google Scholar] [CrossRef]
- Samuels, G.J.; Hallett, I.C. Microdochium stoveri and Monographella stoveri, new combinations for Fusarium stoveri and Micronectriella stoveri. Trans. Br. Mycol. Soc. 1983, 81, 473–483. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Phylogenetic relationships of five genera of Xylariales and Rosasphaeria gen. nov. (Hypocreales). Fungal Divers 2012, 52, 75–98. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, O.; Aime, C.; et al. The Amsterdam declaration on fungal nomenclature. IMA Fungus 2011, 2, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Læssøe, T.; Srikitikulchai, P.; Luangsa-ard, J.J.D.; Stadler, M. Theissenia reconsidered, including molecular phylogeny of the type species T Pyrenocrata and a new genus Durotheca (Xylariaceae, Ascomycota). IMA Fungus 2013, 4, 57–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Host/Substrate | Country | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|---|
| LSU | ITS | BTUB | RPB2 | ||||
| Idriella. lunata | CBS 204.56 * | Fragaria chiloensis | USA | KP858981 | KP859044 | – | – |
| Microdochium. albescens | CBS 291.79 | Oryza sativa | Ivory Coast | KP858932 | KP858996 | KP859059 | KP859105 |
| CBS 243.83 | Oryza sativa | Unknown | KP858930 | KP858994 | KP859057 | KP859103 | |
| M. bolleyi | CBS 540.92 | Hordeum vulgare | Syria | KP858946 | KP859010 | KP859073 | KP859119 |
| M. chrysanthemoides | CGMCC3.17929 * | Unnamed Karst Cave | China | KU746736 | KU746690 | – | – |
| CGMCC3.17930 * | Unnamed Karst Cave | China | KU746735 | KU746689 | – | – | |
| M. citrinidiscum | CBS 109067 * | Eichhornia crassipes | Peru | KP858939 | KP859003 | KP859066 | KP859112 |
| M. colombiense | CBS 624.94 * | Musa sapientum | Colombia | KP858935 | KP858999 | KP859062 | KP859108 |
| M. dawsoniorum | BRIP 65649 | Sporobolus | Australia | – | MK966337 | – | – |
| M. fisheri | CBS 242.90 * | Oryza sativa | UK | KP858951 | KP859015 | KP859079 | KP859124 |
| M. hainanense | SAUCC210781 * | Phragmites australis | China | OM959323 | OM956295 | OM981146 | OM981153 |
| SAUCC210782 | Phragmites australis | China | OM959324 | OM956296 | OM981147 | OM981154 | |
| M. indocalami | SAUCC1016 * | Indocalamus longiauritus | China | MT199878 | MT199884 | MT435653 | MT510550 |
| M. lycopodinum | CBS 146.68 | Air samples | The Netherlands | KP858929 | KP858993 | KP859056 | KP859102 |
| CBS 109397 | Phragmites australis | Germany | KP858940 | KP859004 | KP859067 | KP859113 | |
| CBS 109398 | Phragmites australis | Germany | KP858941 | KP859005 | KP859068 | KP859114 | |
| M. majus | CBS 741.79 | Triticum aestivum | Germany | KP858937 | KP859001 | KP859064 | KP859110 |
| M. miscanthi | SAUCC211092 * | Miscanthus sinensis | China | OM957532 | OM956214 | OM981141 | OM981148 |
| SAUCC211093 | Miscanthus sinensis | China | OM957533 | OM956215 | OM981142 | OM981149 | |
| SAUCC211094 | Miscanthus sinensis | China | OM957534 | OM956216 | OM981143 | OM981150 | |
| M. musae | CBS 111018 = CPC 5380 | Musa cv. cavendish | Costa Rica | – | AY293061 | – | – |
| CBS 143499 = CPC 32809 | Musa sp. | Malaysia | MH107941 | MH107894 | – | – | |
| CBS 143500 * = CPC 32689 | Musa sp. | Malaysia | MH107942 | MH107895 | – | MH108003 | |
| CPC 11234 | Musa sp. | Mauritius | MH107943 | MH107896 | – | – | |
| CPC 11240 | Musa sp. | Mauritius | MH107944 | MH107897 | – | – | |
| CPC 16258 | Musa sp. | Mexico | MH107945 | MH107898 | – | – | |
| CPC 32681 | Musa sp. | Malaysia | MH107946 | MH107899 | – | – | |
| M. neoqueenslandicum | CBS 445.95 | Juncus effusus | The Netherlands | KP858933 | KP858997 | KP859060 | KP859106 |
| CBS 108926 * | Agrostis sp. | New Zealand | KP858938 | KP859002 | KP859065 | KP859111 | |
| M. nivale | CBS 116205 * | Triticum aestivum | UK | KP858944 | KP859008 | KP859071 | KP859117 |
| CBS 288.50 | Unknown | Unknown | MH868135 | MH856626 | – | – | |
| M. novae-zelandiae | CBS 143847 | Turf leaves (Poaceae) | New Zealand | – | LT990655 | LT990608 | LT990641 |
| CPC 29693 | Turf leaves (Poaceae) | New Zealand | – | LT990656 | LT990609 | LT990642 | |
| M. paspali | HK-ML-1371 | Paspalum vaginatum | China | – | KJ569509 | KJ569514 | – |
| QH-BA-48 | Paspalum vaginatum | China | – | KJ569510 | KJ569515 | – | |
| SY-LQG66 | Paspalum vaginatum | China | – | KJ569511 | KJ569516 | – | |
| WC-WC-85 | Paspalum vaginatum | China | – | KJ569512 | KJ569517 | – | |
| WN-BD-452 | Paspalum vaginatum | China | – | KJ569513 | KJ569518 | – | |
| M. phragmitis | CBS 285.71 * | Phragmites australis | Poland | KP858949 | KP859013 | KP859077 | KP859122 |
| CBS 423.78 | Phragmites communis | Germany | KP858948 | KP859012 | KP859076 | KP859121 | |
| M. ratticaudae | BRIP 68298 | introduced giant rat’s tail grasses | Australia | MW481666 | MW481661 | – | MW626890 |
| M. rhopalostylidis | CPC 34449 = CBS 145125 * | Rhopalostylis sapida | New Zealand | MK442532 | MK442592 | – | MK442667 |
| M. seminicola | KAS3576 = CBS 139951 * | Maize kernels | Switzerland | KP858974 | KP859038 | KP859101 | KP859147 |
| KAS1516 = CPC 26001 | Grain | Canada | KP858961 | KP859025 | KP859088 | KP859134 | |
| KAS3574 = DAOM 250155 | Maize kernels | Switzerland | KP858973 | KP859037 | KP859100 | KP859146 | |
| KAS3158 = DAOM 250161 | Triticum aestivum | Canada | KP858970 | KP859034 | KP859097 | KP859143 | |
| KAS1527 = DAOM 250165 | Grain | Canada | KP858966 | KP859030 | KP859093 | KP859139 | |
| KAS1473 = DAOM 250176 | Triticum aestivum | Canada | KP858955 | KP859019 | KP859082 | KP859128 | |
| M. sinense | SAUCC211097 * | Miscanthus sinensis | China | OM959225 | OM956289 | OM981144 | OM981151 |
| SAUCC211098 | Miscanthus sinensis | China | OM959226 | OM956290 | OM981145 | OM981152 | |
| M. sorghi | CBS 691.96 | Sorghum halepense | Cuba | KP858936 | KP859000 | KP859063 | KP859109 |
| M. sp. indet. | SAUCC1017 | Indocalamus longiauritus | China | MT199879 | MT199885 | MT435654 | – |
| M. tainanense | CBS 269.76 * | Saccharum officinarum | Taiwan | KP858945 | KP859009 | KP859072 | KP859118 |
| CBS 270.76 | Saccharum officinarum | Taiwan | KP858931 | KP858995 | KP859058 | KP859104 | |
| M. trichocladiopsis | CBS 623.77 * | Triticum aestivum | Unknown | KP858934 | KP858998 | KP859061 | KP859107 |
| M. yunnanense | SAUCC1011 * | Indocalamus longiauritus | China | MT199875 | MT199881 | MT435650 | MT510547 |
| SAUCC1012 | Indocalamus longiauritus | China | MT199876 | MT199882 | – | MT510548 | |
| SAUCC1015 | Indocalamus longiauritus | Chima | MT199877 | MT199883 | MT435652 | MT510549 | |
| SAUCC1018 | Indocalamus longiauritus | Chima | MT199880 | MT199886 | MT435655 | – | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Liu, X.; Zhang, Z.; Xia, J.; Zhang, X.; Meng, Z. Three New Species of Microdochium (Sordariomycetes, Amphisphaeriales) on Miscanthus sinensis and Phragmites australis from Hainan, China. J. Fungi 2022, 8, 577. https://doi.org/10.3390/jof8060577
Liu S, Liu X, Zhang Z, Xia J, Zhang X, Meng Z. Three New Species of Microdochium (Sordariomycetes, Amphisphaeriales) on Miscanthus sinensis and Phragmites australis from Hainan, China. Journal of Fungi. 2022; 8(6):577. https://doi.org/10.3390/jof8060577
Chicago/Turabian StyleLiu, Shubin, Xiaoyong Liu, Zhaoxue Zhang, Jiwen Xia, Xiuguo Zhang, and Zhe Meng. 2022. "Three New Species of Microdochium (Sordariomycetes, Amphisphaeriales) on Miscanthus sinensis and Phragmites australis from Hainan, China" Journal of Fungi 8, no. 6: 577. https://doi.org/10.3390/jof8060577