
Functional Analysis of the Plasma Membrane H+-ATPases of Ustilago maydis

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Culture Media, and Growth Conditions
2.2. Mutant Sporidia Generation
2.3. Growth Kinetics and Glucose Consumption
2.4. Oxygen Consumption
2.5. H+-ATPase Activity Indirect Assay
2.6. Effect of Different Stress Conditions under Cell Growth
2.7. Isolation of Total Plasma Membrane Fraction
2.8. Enzymatic Activity and Kinetic Studies
2.9. Proton Pumping Rate
2.10. Intracellular pH
2.11. Estimation of Plasma Membrane Electrical Potential
2.12. Determination of Protein
3. Results
3.1. Growth and Basal Metabolism
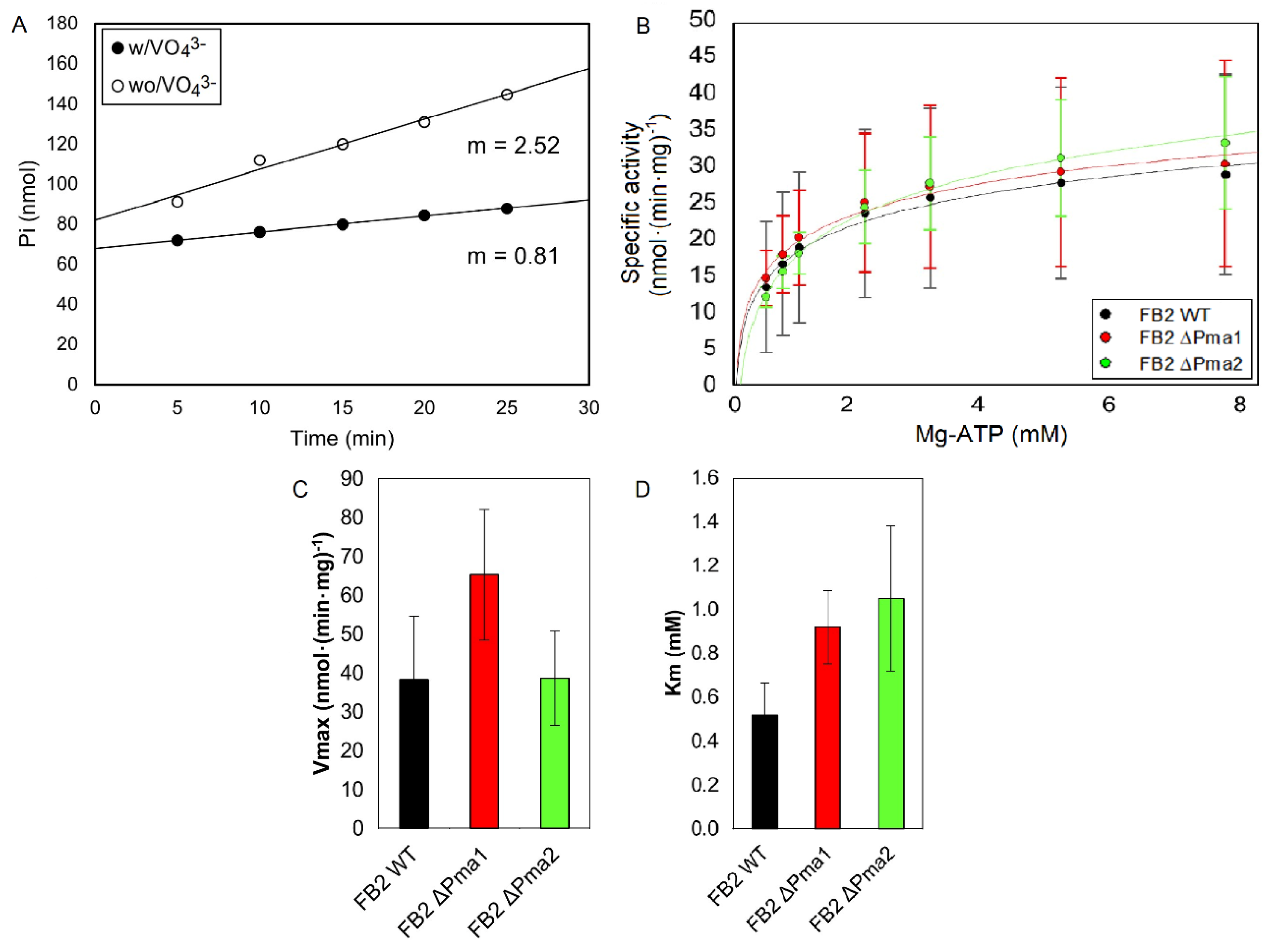
3.2. H+-ATPase Activity
3.3. Proton Pumping Rate and Internal pH
3.4. Effect of an Acidic Load and Nourseothricin on cell Growth
3.5. Effect of Osmotic/Cold Stress on Cell Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goffeau, A.; Slayman, C.W. The proton-translocating ATPase of the fungal plasma membrane. Biochim. Biophys. Acta (BBA)—Rev. Bioenerg. 1981, 639, 197–223. [Google Scholar] [CrossRef]
- Serrano, R. Structure, function and regulation of plasma membrane H (+)-ATPase. FEBS Lett. 1993, 325, 108–111. [Google Scholar] [CrossRef] [Green Version]
- Palmgren, M.; Morsomme, P. The plasma membrane H+-ATPase, a simple polypeptide with a long history. Yeast 2019, 36, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutsenko, S.; Kaplan, J.H. P-type ATPases. Trends Biochem. Sci. 1996, 21, 467. [Google Scholar] [CrossRef]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta (BBA)—Biomembr. 2000, 1465, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kane, P.M. Proton transport and pH control in fungi. Adv. Exp. Med. Biol. 2016, 892, 33–68. [Google Scholar]
- Portillo, F.; Serrano, R. Growth control strength and active site of yeast plasma membrane ATPase studied by site-directed mutagenesis. Eur. J. Biochem. 1989, 186, 501–507. [Google Scholar] [CrossRef]
- Vallejo, C.G.; Serrano, R. Physiology of mutants with reduced expression of plasma membrane H+-ATPase. Yeast 1989, 5, 307–319. [Google Scholar] [CrossRef]
- Gévaudant, F.; Duby, G.; von Stedingk, E.; Zhao, R.; Morsomme, P.; Boutry, M. Expression of a constitutively activated plasma membrane hH+-atpase alters plant development and increases salt tolerance. Plant Physiol. 2007, 144, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Struck, C.; Siebels, C.; Rommel, O.; Wernitz, M.; Hahn, M. The plasma membrane H+-ATPase from the biotrophic rust fungus Uromyces fabae: Molecular characterization of the gene (PMA1) and functional expression of the enzyme in yeast. Mol. Plant-Microbe Interact. 1998, 11, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Morsomme, P.; Slayman, C.W.; Goffeau, A. Mutagenic study of the structure, function and biogenesis of the yeast plasma membrane H(+)-ATPase. Biochim. Biophys. Acta 2000, 1469, 133–1357. [Google Scholar] [CrossRef]
- Serrano, R.; Kielland-Brandt, M.C.; Fink, G.R. Yeast plasma membrane ATPase is essential for growth and has homology with (Na+ + K+), K+- and Ca2+-ATPases. Nature 1986, 319, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Schlesser, A.; Ulaszewski, S.; Ghislain, M.; Goffeau, A. A second transport ATPase gene in Saccharomyces cerevisiae. J. Biol. Chem. 1988, 263, 19480–19487. [Google Scholar] [CrossRef]
- Serrano, R. Characterization of the plasma membrane ATPase of Saccharomyces cerevisiae. Mol. Cell. Biochem. 1978, 22, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Bowman, B.J.; Slayman, C.W. Characterization of plasma membrane adenosine triphosphatase of Neurospora crassa. J. Biol. Chem. 1977, 252, 3357–3363. [Google Scholar] [CrossRef]
- Scarborough, G.A. Isolation and characterization of Neurospora crassa plasma membranes. J. Biol. Chem. 1975, 250, 1106–1111. [Google Scholar] [CrossRef]
- Dufour, J.P.; Goffeau, A. Molecular and kinetic properties of the purified plasma membrane ATPase of the yeast Schizosaccharomyces pombe. Eur. J. Biochem. 1980, 105, 145–154. [Google Scholar] [CrossRef]
- Both, M.; Eckert, S.E.; Csukai, M.; Müller, E.; Dimopoulos, G.; Spanu, P.D. Transcript profiles of Blumeria graminis development during infection reveal a cluster of genes that are potential virulence determinants. Mol. Plant-Microbe Interact. 2005, 18, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Robles-Martínez, L.; Pardo, J.P.; Miranda, M.; Mendez, T.L.; Matus-Ortega, M.G.; Mendoza-Hernández, G.; Guerra-Sánchez, G. The basidiomycete Ustilago maydis has two plasma membrane H+-ATPases related to fungi and plants. J. Bioenerg. Biomembr. 2013, 45, 477–490. [Google Scholar] [CrossRef]
- Holliday, R. Ustilago maydis. In Bacteria, Bacteriophages, and Fungi; Springer: Boston, MA, USA, 1974; pp. 575–595. [Google Scholar]
- Terfrüchte, M.; Joehnk, B.; Fajardo-Somera, R.; Braus, G.H.; Riquelme, M.; Schipper, K.; Feldbrügge, M. Establishing a versatile Golden Gate cloning system for genetic engineering in fungi. Fungal Genet. Biol. 2014, 62, 1–10. [Google Scholar] [CrossRef]
- Punt, P.J.; Van den Hondel, C.A. Transformation of Filamentous Fungi Based on Hygromycin B and Phleomycin Resistance Markers. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 216, pp. 447–457. [Google Scholar]
- Sierra-Campos, E.; Velázquez, I.; Matuz-Mares, D.; Villavicencio-Queijeiro, A.; Pardo, J. Functional properties of the Ustilago maydis alternative oxidase under oxidative stress conditions. Mitochondrion 2009, 9, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Goossens, A.; De la Fuente, N.; Forment, J.; Serrano, R.; Portillo, F. Regulation of yeast H+-ATPase by protein kinases belonging to a family dedicated to activation of plasma membrane transporters. Mol. Cell. Biol. 2000, 20, 7654–7661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, R. In vivo glucose activation of the yeast plasma membrane ATPase. FEBS Lett. 1983, 156, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, R.K.; Rao, R.; Slayman, C.W. Expression of the yeast plasma membrane [H+] ATPase in secretory vesicles. A new strategy for directed mutagenesis. J. Biol. Chem. 1991, 266, 7940–7949. [Google Scholar] [CrossRef]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Peña, A.; Ramírez, J.; Rosas, G.; Calahorra, M. Proton pumping and the internal pH of yeast cells, measured with pyranine introduced by electroporation. J. Bacteriol. 1995, 177, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Calahorra, M.; Martínez, G.A.; Hernández-Cruz, A.; Peña, A. Influence of monovalent cations on yeast cytoplasmic and vacuolar ph. Yeast 1998, 14, 501–515. [Google Scholar] [CrossRef]
- Peña, A.; Uribe, S.; Pardo, J.P.; Borbolla, M. The use of a cyanine dye in measuring membrane potential in yeast. Arch. Biochem. Biophys. 1984, 231, 217–225. [Google Scholar] [CrossRef]
- Bensadoun, A.; Weinstein, D. Assay of proteins in the presence of interfering materials. Anal. Biochem. 1976, 70, 241–250. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Drew, B.; Leeuwenburgh, C. Method for measuring ATP production in isolated mitochondria: ATP production in brain and liver mitochondria of Fischer-344 rats with age and caloric restriction. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2003, 285, 1259–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponce-Pineda, I.G.; Carmona-Salazar, L.; Saucedo-García, M.; Cano-Ramírez, D.; Morales-Cedillo, F.; Peña-Moral, A.; Guevara-Garcia, A.A.; Sanchez-Nieto, S.; Gavilanez-Ruiy, M. MPK6 Kinase regulates plasma membrane H+-ATPase activity in cold acclimation. Int. J. Mol. Sci. 2021, 22, 6338. [Google Scholar] [CrossRef] [PubMed]
- Scalla, R.; Amory, A.; Rigaud, J.; Goffeau, A. Phosphorylated intermediate of a transport ATPase and activity of protein kinase in membranes from corn roots. Eur. J. Biochem. 1983, 132, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Vara, F.; Serrano, R. Partial purification and properties of the proton-translocating ATPase of plant plasma membranes. J. Biol. Chem. 1982, 257, 12826–12830. [Google Scholar] [CrossRef]
- Villalobo, A.; Boutry, M.; Goffeau, A. Electrogenic proton translocation coupled to ATP hydrolysis by the plasma membrane Mg2+-dependent ATPase of yeast in reconstituted proteoliposomes. J. Biol. Chem. 1981, 256, 12081–12087. [Google Scholar] [CrossRef]
- Haupt, I.; Jonák, J.; Rychlík, I.; Thrum, H. Action of streptothricin F on ribosomal functions. J. Antibiot. 1980, 33, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Olfatmiri, H.; Alemzadeh, A.; Zakipour, Z. Up-regulation of plasma membrane H+-ATPase under salt stress may enable Aeluropus littoralis to cope with stress. Mol. Biol. Res. Commun. 2014, 3, 67–75. [Google Scholar]
- Muzi, C.; Camoni, L.; Visconti, S.; Aducci, P. Cold stress affects H+-ATPase and phospholipase D activity in Arabidopsis. Plant Physiol. Biochem. 2016, 108, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Sigler, K.; Höfer, M. Mechanisms of acid extrusion in yeast. Biochim. Biophys. Acta 1991, 1071, 375–391. [Google Scholar] [CrossRef]
- Kotyk, A.; Georghiou, G. Effects of the physiological state of five yeast species on H+-ATPase-related processes. Folia Microbiol. 1993, 38, 467–472. [Google Scholar] [CrossRef]
- Sigler, K.; Knotková, A.; Páca, J.; Wurst, M. Extrusion of metabolites from baker’s yeast during glucose-induced acidification. Folia Microbiol. 1980, 25, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Wurst, M.; Sigler, K.; Knotková, A. Gas chromatographic determination of extracellular metabolites produced by baker’s yeast during glucose-induced acidification. Folia Microbiol. 1980, 25, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, E.; Ramos-Casillas, L.E.; Marín-Hernández, A.; Moreno-Sánchez, R.; Guerra-Sánchez, G. Glycolysis in Ustilago maydis. FEMS Yeast Res. 2008, 8, 1313–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucena, R.M.; Dolz-Edo, L.; Brul, S.; De Morais, M.A.; Smits, G. Extreme low cytosolic pH is a signal for cell survival in acid stressed yeast. Genes 2020, 11, 656. [Google Scholar] [CrossRef]
- Eraso, P.; Gancedo, C. Activation of yeast plasma membrane ATPase by acid pH during growth. FEBS Lett. 1987, 224, 187–192. [Google Scholar] [CrossRef]
- Holyoak, C.D.; Stratford, M.; McMullin, Z.; Cole, M.B.; Crimmins, K.; Brown, A.J.; Coote, P.J. Activity of the plasma membrane H+-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl. Environ. Microbiol. 1996, 62, 3158–3164. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Phenotype | Source |
|---|---|---|---|
| FB2 WT | a2b2; pma1, pma2 | Pma1, Pma2 | [19] |
| FB2 ΔPma1 | a2b2; pma2 | Pma2 | This study |
| FB2 ΔPma2 | a2b2; pma1 | Pma1 | This study |
| Primer | Sequence (5′–3′) | Use |
|---|---|---|
| ΔPma1 | ||
| Upstream flank, Forward primer | CGTAGGCCTCGCTTGTTG | Flank construction |
| Upstream flank, Reverse primer | GGTCTCGCCTGCAATATTTGTTCTTGCCTCGTCCTGTC | Flank construction |
| Downstream flank, Forward primer | GGTCTCCAGGCCGATGAAAGAAAAAAGACTACCG | Flank construction |
| Downstream flank, Reverse primer | GGTCTCCGGCCACCGAGATGCATGCTCACATTC | Flank construction |
| Diagnostic P1, Forward primer | CGGTGTTGCCATGAACACCGATGGCCAGTG | Diagnostic PCR |
| Diagnostic P2, Reverse primer | GAGGGCAACGGATTCGAGCTTCTTGGTCTT | Diagnostic PCR |
| DIG-probe 1, Forward primer | ACGACGTTGTAAAACGACGGCCAG | DIG-probe 1 for Southern blot |
| DIG-probe 1, Reverse primer | GGTCTCCAGGCCGATGAAAGAAAAAAGACTAC CG | DIG-probe 1 for Southern blot |
| DIG-probe 2, Forward primer | CGGTGTTGCCATGAACACCGATGGCCAGTG | DIG-probe 2 for Southern blot |
| DIG-probe 2, Reverse primer | CCAGGTGGAGACAGAGCG | DIG-probe 2 for Southern blot |
| DIG-probe 3, Forward primer | GGTCTCCGGCCACCGAGATGCATGCTCACATTC | DIG-probe 3 for Southern blot |
| DIG-probe 3, Reverse primer | TTCACACAGGAAACAGCTATGACC | DIG-probe 3 for Southern blot |
| ΔPma2 | ||
| Upstream flank, Forward primer | GGTCTCGCCTGCAATATTCAACCTCTAAGACTCGCTT | Flank construction |
| Upstream flank, Reverse primer | GGTCTCCAGGCCTCTGCCTCTTATCTTGCTCTCTTAG | Flank construction |
| Downstream flank, Forward primer | GGTCTCCGGCCGGGGAAACGTGGAGAAGGTCGCGAAA | Flank construction |
| Downstream flank, Reverse primer | GGTCTCGCTGCAATATTACCACCCTGTGCCCTCTAG | Flank construction |
| Diagnostic P1, Forward primer | ACGCTTGACAATCTCGTACTTGTGCTCGGGG | Diagnostic PCR |
| Diagnostic P2, Reverse primer | GAGGGCAACGGATTCGAGCTTCTTGGTCTT | Diagnostic PCR |
| DIG-probe 1, Forward primer | GCGCAACTGTTGGGAAGG | DIG-probe 1 for Southern blot |
| DIG-probe 1, Reverse primer | GGTCTCCAGGCCTCTGCCTCTTATCTTGCTCTCTTAG | DIG-probe 1 for Southern blot |
| DIG-probe 2, Forward primer | ACGCTTGACAATCTCGTACTTGTGCTCGGGG | DIG-probe 2 for Southern blot |
| DIG-probe 2, Reverse primer | CCCTCATTGGCTCCGACG | DIG-probe 2 for Southern blot |
| DIG-probe 3, Forward primer | GGTCTCCGGCCGGGGAAACGTGGAGAAGGTCGCGAAA | DIG-probe 3 for Southern blot |
| DIG-probe 3, Reverse primer | TGGAAAGCGGGCAGTGAG | DIG-probe 3 for Southern blot |
| Plasmid | Use | Source |
| pUMa1810 | Transforming plasmid to delete pma1 gene | This study |
| pUMa4515 | Transforming plasmid to delete pma2 gene | This study |
| pUMa1507 | Hygromycin resistance cassette | [21] |
| pUMa1467 | Destination vector | [21] |
| Strain | Duplication Time (h) |
|---|---|
| FB2 WT | 2.65 ± 0.90 |
| FB2 ΔPma1 | 3.52 ± 1.11 |
| FB2 ΔPma2 | 2.64 ± 0.16 |
| Strain | Internal pH | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| FB2 WT | 7.30 ± 0.05 | 7.11 ± 0.05 | 7.66 ± 0.08 |
| FB2 ΔPma1 | 7.19 ± 0.04 | 7.39 ± 0.03 | 7.26 ± 0.63 |
| FB2 ΔPma2 | 7.07 ± 1.23 | 7.52 ± 0.22 | 7.24 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Carrada, M.; Feldbrügge, M.; Olicón-Hernández, D.R.; Guerra-Sánchez, G.; Pardo, J.P. Functional Analysis of the Plasma Membrane H+-ATPases of Ustilago maydis. J. Fungi 2022, 8, 550. https://doi.org/10.3390/jof8060550
Vázquez-Carrada M, Feldbrügge M, Olicón-Hernández DR, Guerra-Sánchez G, Pardo JP. Functional Analysis of the Plasma Membrane H+-ATPases of Ustilago maydis. Journal of Fungi. 2022; 8(6):550. https://doi.org/10.3390/jof8060550
Chicago/Turabian StyleVázquez-Carrada, Melissa, Michael Feldbrügge, Dario Rafael Olicón-Hernández, Guadalupe Guerra-Sánchez, and Juan Pablo Pardo. 2022. "Functional Analysis of the Plasma Membrane H+-ATPases of Ustilago maydis" Journal of Fungi 8, no. 6: 550. https://doi.org/10.3390/jof8060550