
The Entomopathogenic Fungus Metarhizium anisopliae Affects Feeding Preference of Sogatella furcifera and Its Potential Targets’ Identification


Abstract
:1. Introduction
2. Methods and Materials
2.1. Insect and Plant Culture
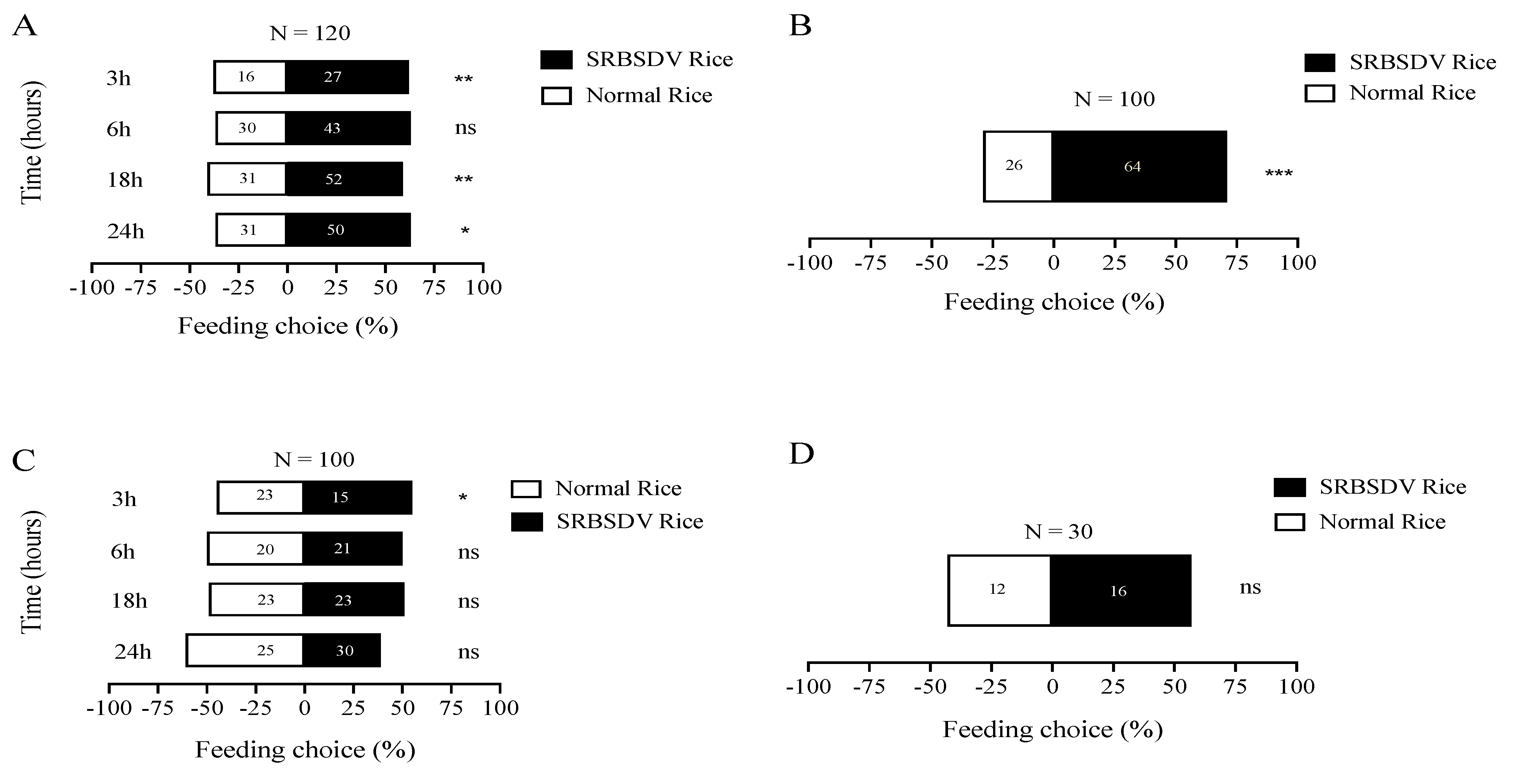
2.2. Effects of M. anisopliae on S. furcifera Feeding Behavior
2.3. Total RNA Isolation, Quantification and Sequencing
2.4. Data Analysis
2.5. Validation of DEGs Library and Olfactory-Related Genes Identification
3. Results
3.1. Effects of M. anisopliae on Feeding Choice of S. furcifera
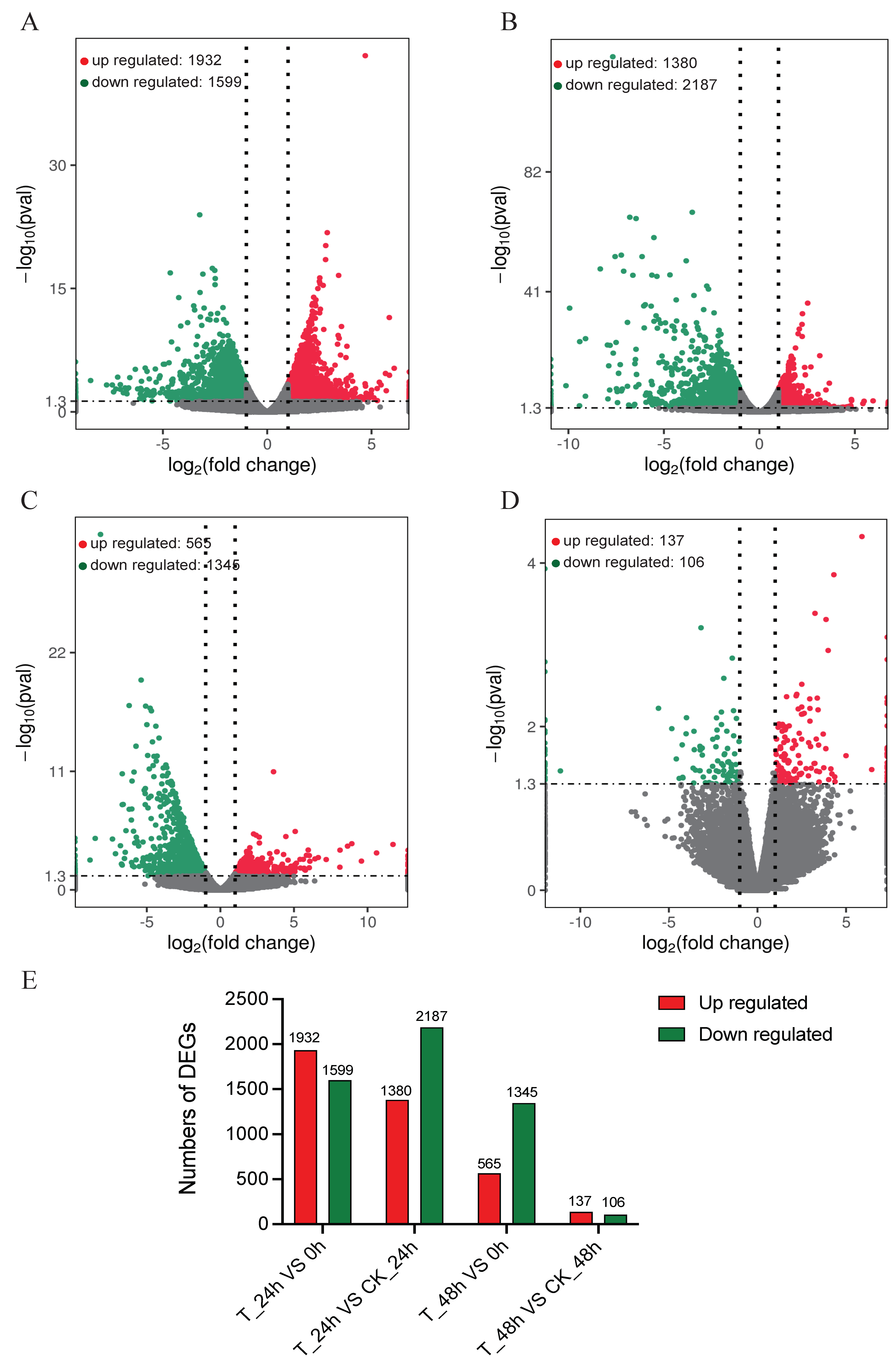
3.2. Summary of Digital Gene Expression by Transcriptomic Profiling
3.3. Transcriptomic Comparison and Analysis of M. anisopliae Inoculation
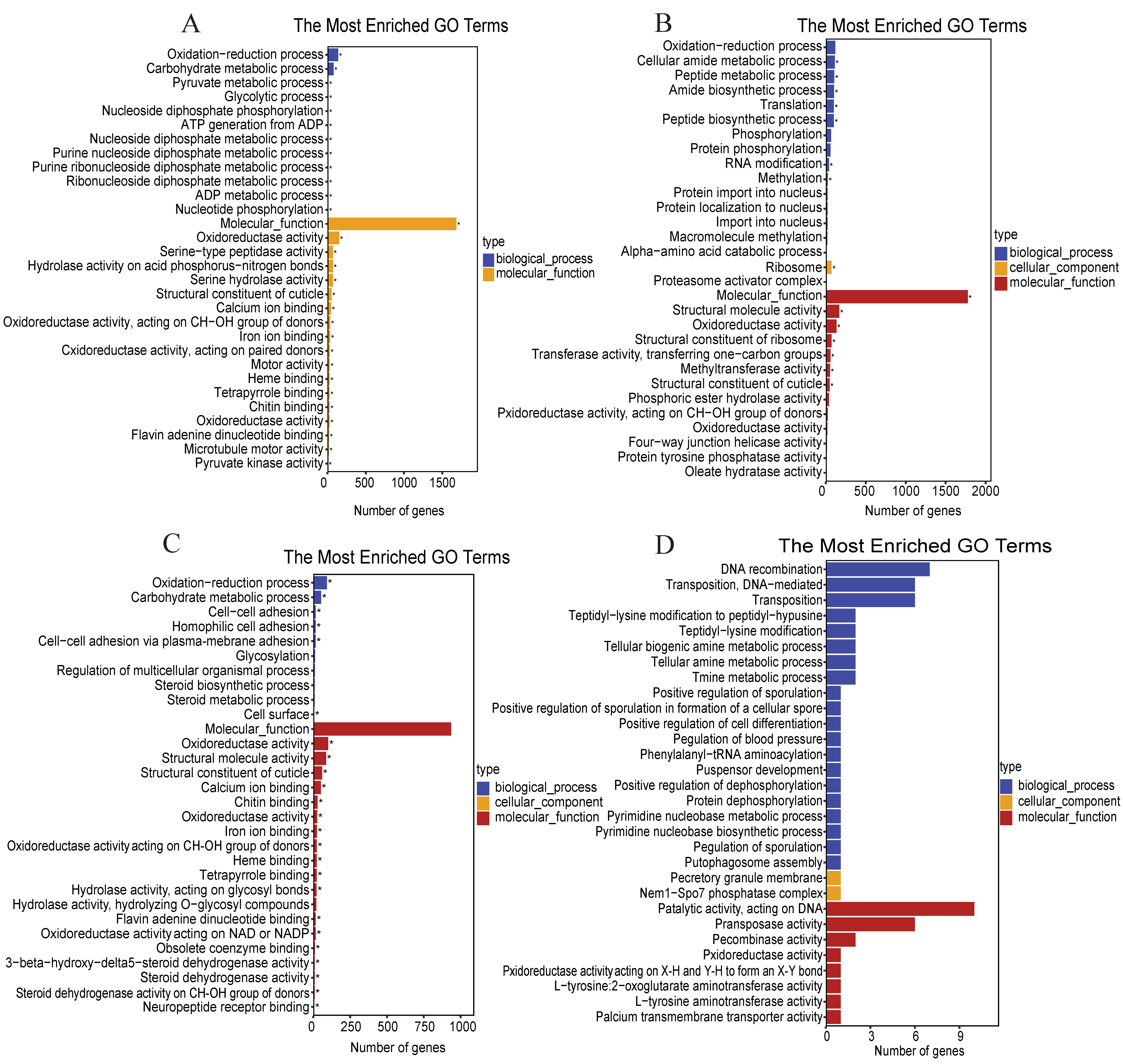
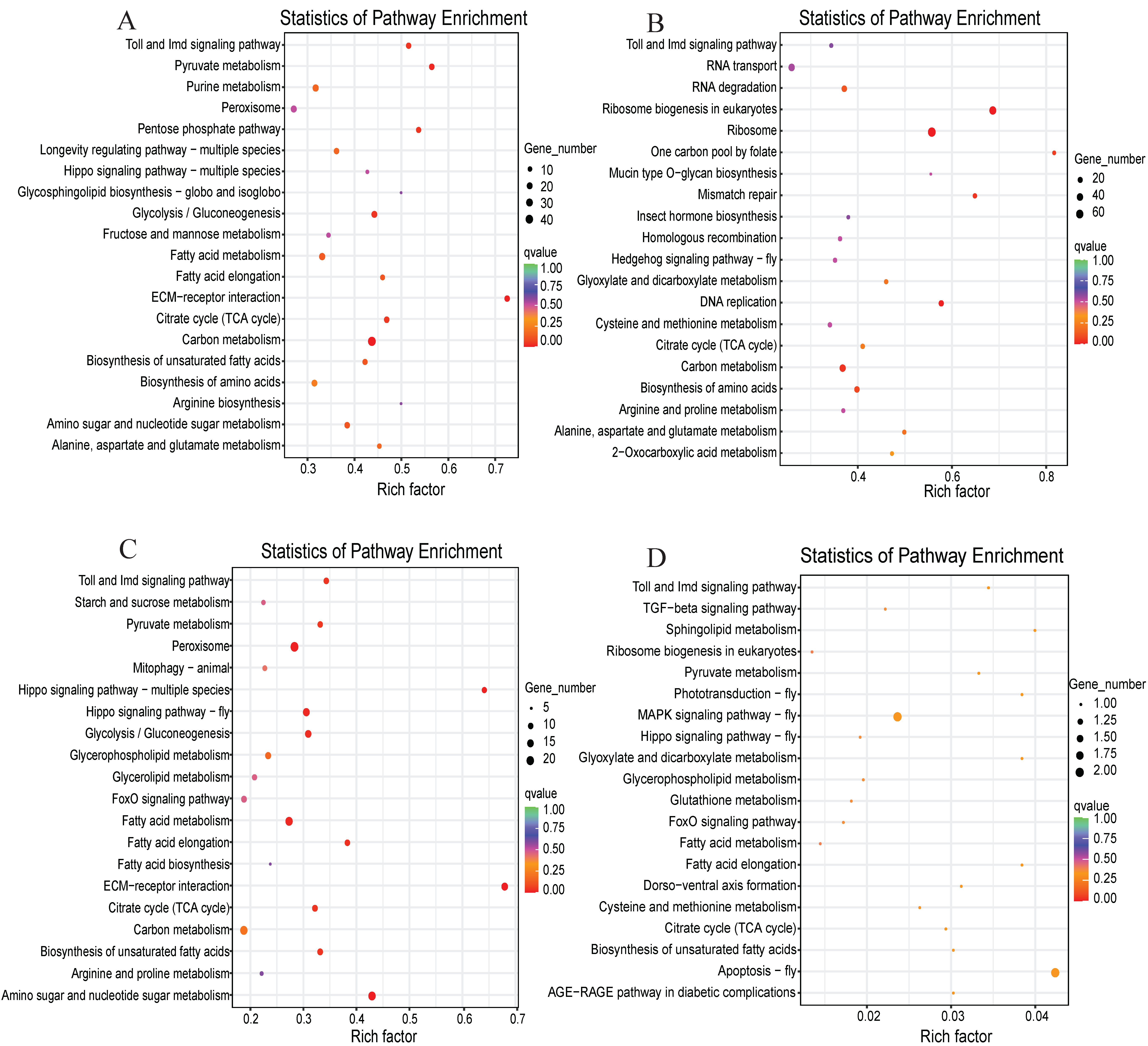
3.4. Functional Classification and Pathway Analysis
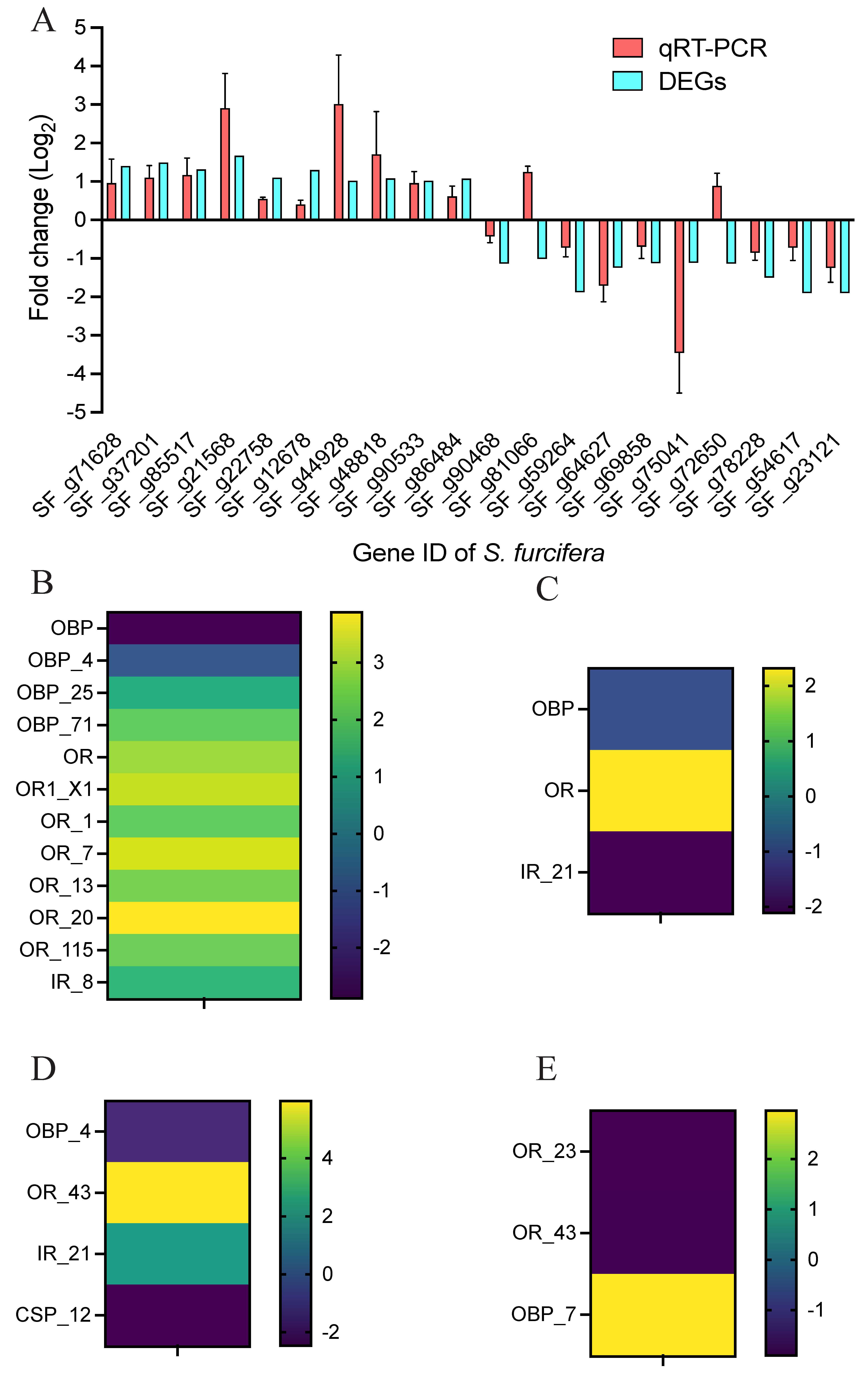
3.5. Validation of DEGs Using RT-qPCR and Olfactory-Related Genes Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.C.; Lee, S.C. Occurrence of major rice insect pests at different transplanting times and fertilizer levels in paddy field. Korean J. Appl. Entomol. 1996, 35, 132–136. [Google Scholar]
- Wei, T.Y.; Li, Y. Rice Reoviruses in insect vectors. Annu. Rev. Phytopathol. 2016, 54, 99–120. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.G.; Zhang, G.R.; Zhang, W.Q.; Hu, Y.; Zhang, J. Biological control of rice insect pests in China. Biol. Control 2013, 67, 8–20. [Google Scholar] [CrossRef]
- Cheng, J. Rice planthopper problems and relevant causes in China. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Los Baños, Philippines, 2009; Chapter 1; pp. 157–178. [Google Scholar]
- Zhou, G.H.; Xu, D.L.; Xu, D.G.; Zhang, M.X. Southern rice black-streaked dwarf virus: A white-backed planthopper-transmitted fijivirus threatening rice production in Asia. Front. Microbiol. 2013, 4, 270. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, G.; Chen, X.; Li, X.J.; Xiong, K.; Cao, S.P.; Hu, Y.Y.; Lu, M.H.; Liu, W.C.; Tuan, H.A.; et al. The influence of Sogatella furcifera (Hemiptera: Delphacidae) migratory events on the southern rice black-streaked dwarf virus epidemics. J. Econ. Entomol. 2017, 110, 854–864. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Hamiduzzaman, M.M.; Sinia, A.; Guzman-Novoa, E.; Goodwin, P.H. Entomopathogenic fungi as potential biocontrol agents of the ecto-parasitic mite, Varroa destructor, and their effect on the immune response of honey bees (Apis mellifera L.). J. Invertebr. Pathol. 2012, 111, 237–243. [Google Scholar] [CrossRef]
- Sánchezpeña, S.R.; Lara, J.S.; Medina, R.F. Occurrence of entomopathogenic fungi from agricultural and natural ecosystems in Saltillo, México, and their virulence towards thrips and whiteflies. J. Insect Sci. 2011, 11, 1. [Google Scholar] [CrossRef]
- Bordalo, M.D.; Gravato, C.; Beleza, S.; Campos, D.; Lopes, I.; Pestana, J.L.T. Lethal and sublethal toxicity assessment of Bacillus thuringiensis var. israelensis and Beauveria bassiana based bioinsecticides to the aquatic insect Chironomus riparius. Sci. Total Environ. 2020, 698, 134155. [Google Scholar] [CrossRef]
- Jiang, W.; Peng, Y.; Ye, J.; Wen, Y.; Liu, G.; Xie, J. Effects of the entomopathogenic fungus Metarhizium anisopliae on the mortality and immune response of Locusta migratoria. Insects 2019, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Fite, T.; Tefera, T.; Negeri, M.; Damte, T.; Sori, W. Evaluation of Beauveria bassiana, Metarhizium anisopliae, and Bacillus thuringiensis for the management of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) under laboratory and field conditions. Biocontrol Sci. Technol. 2019, 30, 278–295. [Google Scholar] [CrossRef]
- Rice, S.J.; Baker, D.K.; Leemon, D.M. Development of mycoinsecticide formulations with Beauveria bassiana and Metarhizium anisopliae for the control of lesser mealworm, Alphitobius diaperinus, in chicken broiler houses. Biocontrol 2019, 64, 489–500. [Google Scholar] [CrossRef]
- Peng, G.X.; Xie, J.Q.; Guo, R.; Keyhani, N.O.; Zeng, D.Y.; Yang, P.Y.; Xia, Y.X. Long-term field evaluation and large-scale application of a Metarhizium anisopliae strain for controlling major rice pests. J. Pest Sci. 2021, 94, 969–980. [Google Scholar] [CrossRef]
- Tang, J.; Liu, X.; Ding, Y.; Jiang, W.; Xie, J. Evaluation of Metarhizium anisopliae for rice planthopper control and its synergy with selected insecticides. Crop Prot. 2019, 121, 132–138. [Google Scholar] [CrossRef]
- Pan, C.Y.; Cai, Y.J.; Li, T.C.; Zhang, W.Q. Control effect of Metarhizium anisopliae and its mixture with dsRNA on the brown planthopper. J. Environ. Entomol. 2016, 38, 1071–1077. [Google Scholar]
- Fang, W.; Vega-Rodriguez, J.; Ghosh, A.K.; Jacobs-Lorena, M.; Kang, A.; St. Leger, R.J. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 2011, 331, 1074–1077. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, S. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73. [Google Scholar] [CrossRef]
- Duressa, T.F.; Vanlaer, R.; Huybrechts, R. Locust cellular defense against inoculations: Sites of pathogen clearance and hemocyte proliferation. Dev. Comp. Immunol. 2015, 48, 244–253. [Google Scholar] [CrossRef]
- Olombrada, M.; Lazaro-Gorines, R.; Lopez-Rodriguez, J.C.; Martinez-Del-Pozo, A.; Onaderra, M.; Maestro-Lopez, M.; Lacadena, J.; Gavilanes, J.G.; Garcia-Ortega, L. Fungal Ribotoxins: A review of potential biotechnological applications. Toxins 2017, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, M.; Xu, X.; Xu, J.; Li, S.; Yu, J.; Zhou, X.; Xu, X.; Hu, Q.; Yu, X.; Jin, F. Genome-Wide identification of destruxin A-responsive immunity-related microRNAs in diamondback moth, Plutella xylostella. Front. Immunol. 2018, 9, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Cao, Y.; Xia, Y.; Liu, F. Wright-Giemsa staining to observe phagocytes in Locusta migratoria infected with Metarhizium acridum. J. Invertebr. Pathol. 2016, 139, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.L.; St. Leger, R.J. Insect immunity to entomopathogenic fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar] [PubMed]
- Xie, J.; Peng, Y.; Xia, Y. Genome-Wide identification and analysis of Nilaparvata lugens microRNAs during challenge with the entomopathogenic fungus Metarhizium anisopliae. J. Fungi 2021, 7, 295. [Google Scholar] [CrossRef] [PubMed]
- Junges, A.; Boldo, J.T.; Souza, B.K.; Guedes, R.L.; Sbaraini, N.; Kmetzsch, L.; Thompson, C.E.; Staats, C.C.; de Almeida, L.G.; de Vasconcelos, A.T.; et al. Genomic analyses and transcriptional profiles of the glycoside hydrolase family 18 genes of the entomopathogenic fungus Metarhizium anisopliae. PLoS ONE 2014, 9, e107864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Xu, X.X.; Shakeel, M.; Li, S.Z.; Wang, S.; Zhou, X.Q.; Yu, J.L.; Xu, X.J.; Yu, X.Q.; Jin, F.L. The Entomopathogenic fungi Isaria fumosorosea plays a vital role in suppressing the immune system of Plutella xylostella: RNA-Seq and DGE analysis of immunity-related genes. Front. Microbiol. 2017, 8, 1421. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Del Marmol, J.; Yedlin, M.A.; Ruta, V. The structural basis of odorant recognition in insect olfactory receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef]
- Zhang, J.; Komail Raza, S.A.; Wei, Z.; Keesey, I.W.; Parker, A.L.; Feistel, F.; Chen, J.; Cassau, S.; Fandino, R.A.; Grosse-Wilde, E.; et al. Competing beetles attract egg laying in a hawkmoth. Curr. Biol. 2022, 32, 861–869. [Google Scholar] [CrossRef]
- Guo, X.J.; Yu, Q.Q.; Chen, D.F.; Wei, J.N.; Yang, P.C.; Yu, J.; Wang, X.H.; Kang, L. 4-Vinylanisole is an aggregation pheromone in locusts. Nature 2020, 584, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; He, X.; Zheng, X.; Yang, Y.; Tian, J.; Lu, Z. Southern rice black-streaked dwarf virus (SRBSDV) directly affects the feeding and reproduction behavior of its vector, Sogatella furcifera (Horvath) (Hemiptera: Delphacidae). Virol. J. 2014, 11, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.S.; Qin, J.H.; Cao, Y.Z.; Li, K.B.; Yin, J. Two classic OBPs modulate the responses of female Holotrichia oblita to three major ester host plant volatiles. Insect Mol. Biol. 2021, 30, 390–399. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Chen, G.L.; Li, S.; Wang, J.; Ma, Y.F.; Pan, Y.F.; He, M. Evolution and functional analysis of odorant-binding proteins in three rice planthoppers: Nilaparvata lugens, Sogatella furcifera, and Laodelphax striatellus. Pest Manag. Sci. 2019, 75, 1606–1620. [Google Scholar] [CrossRef]
- Zhan, H.; Dewer, Y.; Zhang, J.; Tian, J.; Li, D.; Qu, C.; Yang, Z.; Li, F.; Luo, C. Odorant-binding protein 1 plays a crucial role in the olfactory response of Bemisia tabaci to R-curcumene. J. Agric. Food Chem. 2021, 69, 12785–12793. [Google Scholar] [CrossRef]
- Zhou, G.H.; Wen, J.J.; Cai, D.J.; Li, P.; Xu, D.L.; Zhang, S.G. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Peng, Y.; Tang, J.; Hong, M.; Xie, J. Suppression of rice planthopper populations without affecting the rice microbiota by the entomopathogenic fungus Metarhizium anisopliae. Appl. Environ. Microbiol. 2020, 86, e01337-20. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, W.; Zhou, Y.; Wu, J.; Zhou, X. Transcriptome and comparative gene expression analysis of Sogatella furcifera (Horvath) in response to southern rice black-streaked dwarf virus. PLoS ONE 2012, 7, e36238. [Google Scholar] [CrossRef]
- Lin, J.; He, J.; Liang, A.; Wang, F. Transcriptome profiling and dimorphic expression of sex-related genes in fifth-instar nymphs of Sogatella furcifera, an important rice pest. Genomics 2020, 112, 1105–1111. [Google Scholar] [CrossRef]
- Yang, S.; Xu, R.; Yang, S.-Y.; Kuang, R.-P. Olfactory responses of Aphidius gifuensis to odors of host plants and aphid-plant complexes. Insect Sci. 2009, 16, 503–510. [Google Scholar] [CrossRef]
- Wang, L.; Tang, N.; Gao, X.; Chang, Z.; Zhang, L.; Zhou, G.; Guo, D.; Zeng, Z.; Li, W.; Akinyemi, I.A.; et al. Genome sequence of a rice pest, the white-backed planthopper (Sogatella furcifera). Gigascience 2017, 6, giw004. [Google Scholar]
- Cheng, J. Rice Planthoppers in the Past Half Century in China. In Rice Planthoppers; Springer: Dordrecht, The Netherlands, 2015; pp. 1–32. [Google Scholar]
- Liao, X.; Xu, P.F.; Gong, P.P.; Wan, H.; Li, J.H. Current susceptibilities of brown planthopper Nilaparvata lugens to triflumezopyrim and other frequently used insecticides in China. Insect Sci. 2021, 28, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 1996, 34, 249–274. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chai, R.; Liu, X.; Dong, Y.; Su, D.; Desneux, N.; Tan, X.; Luo, C. Barley yellow dwarf virus-infected wheat plant modulated selection behavior of vector aphids. J. Pest Sci. 2021, 95, 1273–1285. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Guo, H.; Zhu-Salzman, K.; Ge, F.; Sun, Y. PEBP balances apoptosis and autophagy in whitefly upon arbovirus infection. Nat. Commun. 2022, 13, 846. [Google Scholar] [CrossRef]
- Wang, S.; Guo, H.; Ge, F.; Sun, Y. Apoptotic neurodegeneration in whitefly promotes the spread of TYLCV. Elife 2020, 9, e56168. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Perez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 578. [Google Scholar] [CrossRef] [Green Version]
- Dan, H. Drosophila immunity: Paths and patterns. Curr. Opin. Immunol. 2003, 15, 12–19. [Google Scholar]
- Li, R.T.; Huang, L.Q.; Dong, J.F.; Wang, C.Z. A moth odorant receptor highly expressed in the ovipositor is involved in detecting host-plant volatiles. Elife 2020, 9, e53706. [Google Scholar] [CrossRef]
- Wei, Z.; Ortiz-Urquiza, A.; Keyhani, N.O. Altered expression of chemosensory and odorant binding proteins in response to fungal infection in the red imported rire ant, Solenopsis invicta. Front. Physiol. 2021, 12, 596571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, X.; Chen, Y.; Hu, J.; Dong, J.; Su, X.; Zhang, Z. Southern rice black-streaked dwarf virus infection improves host suitability for its insect vector, Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2014, 107, 92–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Q30 |
|---|---|---|---|---|---|
| T_24 h_1 | 38,699,940 | 5.80G | 35,830,030 | 5.37G | 92.91% |
| T_24 h_2 | 42,519,476 | 6.37G | 40,606,948 | 6.09G | 93.49% |
| T_24 h_3 | 38,627,148 | 5.79G | 36,601,008 | 5.49G | 93.25% |
| CK_24 h_1 | 41,683,106 | 6.25G | 39,847,928 | 5.98G | 93.70% |
| CK_24 h_2 | 44,749,802 | 6.71G | 42,760,666 | 6.41G | 93.92% |
| CK_24 h_3 | 46,649,430 | 6.99G | 44,910,818 | 6.74G | 93.08% |
| T_48 h_1 | 40,096,262 | 6.01G | 38,405,376 | 5.76G | 93.02% |
| T_48 h_2 | 42,917,082 | 6.43G | 41,055,774 | 6.16G | 93.73% |
| T_48 h_3 | 40,289,444 | 6.04G | 38,627,478 | 5.79G | 93.26% |
| CK_48 h_1 | 43,887,028 | 6.58G | 42,403,914 | 6.36G | 92.82% |
| CK_48 h_2 | 38,657,406 | 5.79G | 36,866,622 | 5.53G | 93.27% |
| CK_48 h_3 | 40,229,530 | 6.03G | 38,382,242 | 5.76G | 93.54% |
| Blank_1 | 41,866,614 | 6.27G | 40,000,444 | 6.00G | 93.20% |
| Blank_2 | 38,396,532 | 5.75G | 36,692,418 | 5.50G | 94.56% |
| Blank_3 | 43,095,904 | 6.46G | 41,381,568 | 6.21G | 94.07% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Han, L.; Xia, Y.; Xie, J. The Entomopathogenic Fungus Metarhizium anisopliae Affects Feeding Preference of Sogatella furcifera and Its Potential Targets’ Identification. J. Fungi 2022, 8, 506. https://doi.org/10.3390/jof8050506
Wang Y, Han L, Xia Y, Xie J. The Entomopathogenic Fungus Metarhizium anisopliae Affects Feeding Preference of Sogatella furcifera and Its Potential Targets’ Identification. Journal of Fungi. 2022; 8(5):506. https://doi.org/10.3390/jof8050506
Chicago/Turabian StyleWang, Yirong, Lijuan Han, Yuxian Xia, and Jiaqin Xie. 2022. "The Entomopathogenic Fungus Metarhizium anisopliae Affects Feeding Preference of Sogatella furcifera and Its Potential Targets’ Identification" Journal of Fungi 8, no. 5: 506. https://doi.org/10.3390/jof8050506