
Antifungal Potential of Azotobacter salinestris Strain Azt 31 against Phytopathogenic Fusarium spp. Associated with Cereals

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Inoculum Preparation
2.2.1. Bacterial Cell Suspension
2.2.2. Fungal Spore Preparation
2.3. Seed Treatment
2.4. In Vitro Studies of A. salinestris against Fusarium spp.
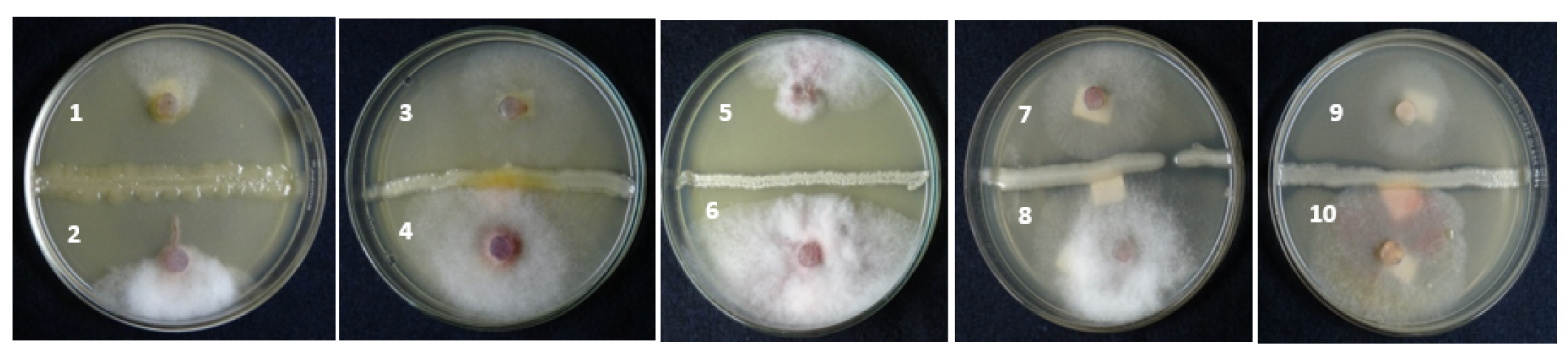
2.4.1. Antagonistic Activity of A. salinestris Investigated by Dual-Culture Method
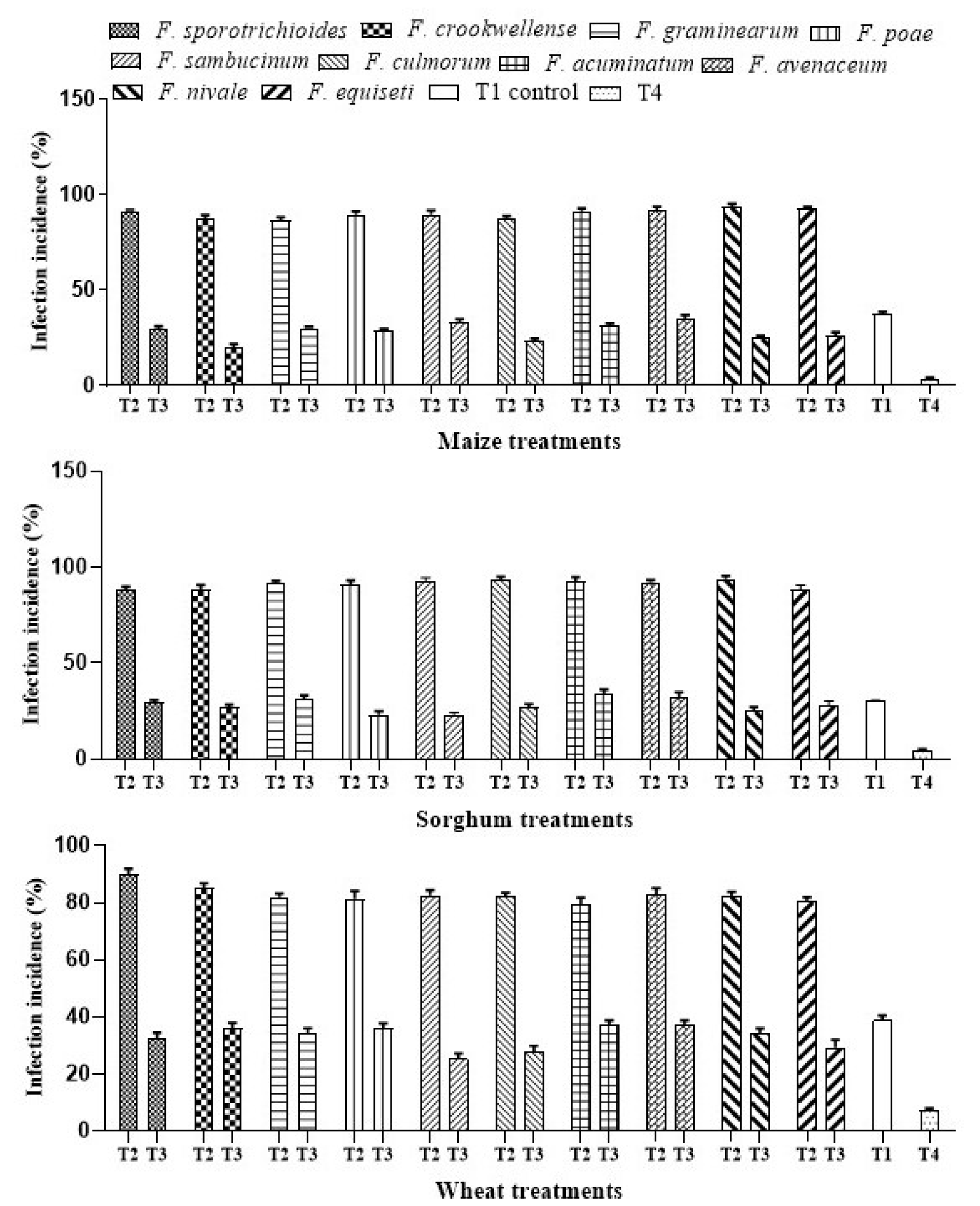
2.4.2. Effect of Azotobacter against Fusarium spp. and Infection Incidence
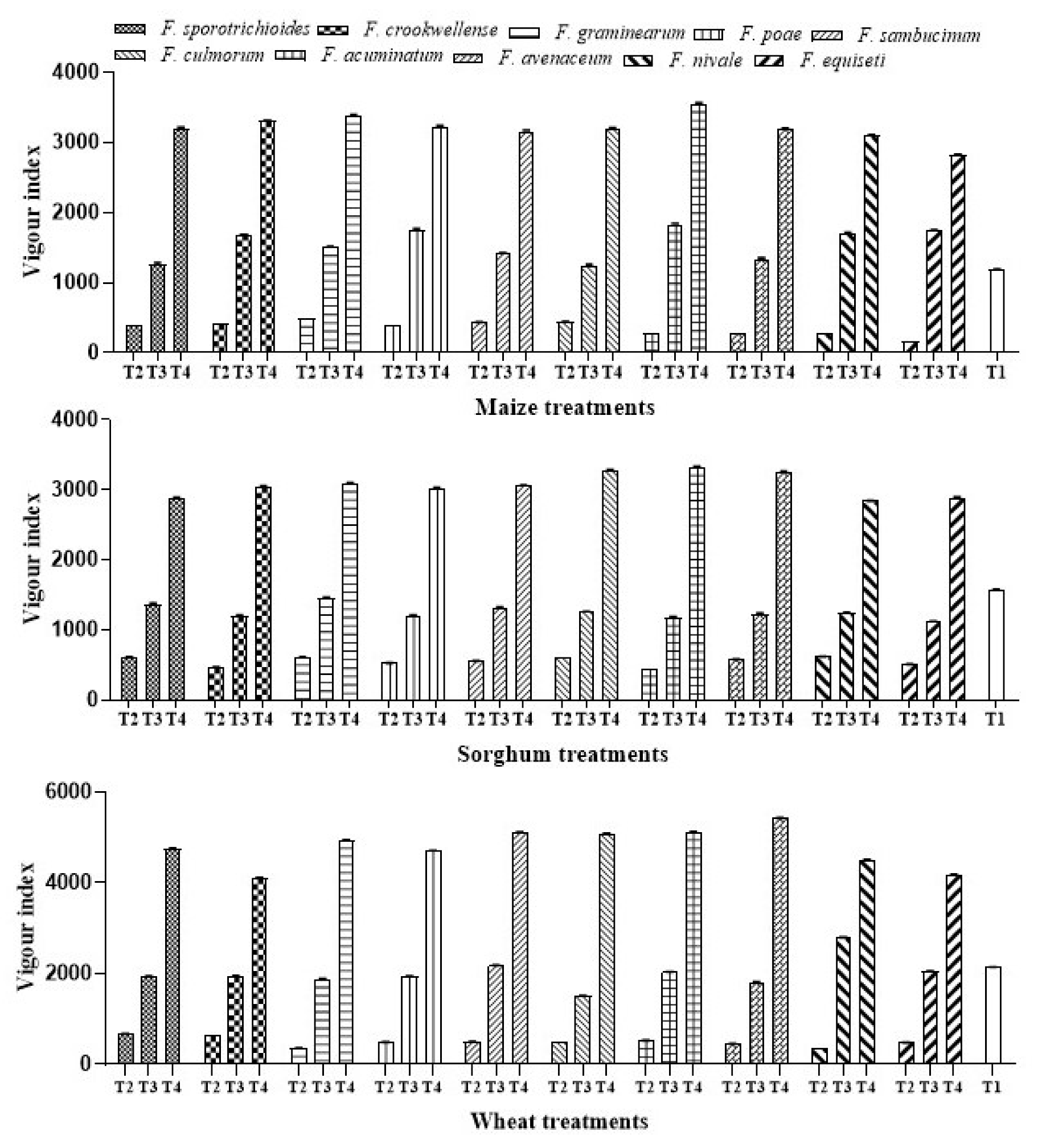
2.4.3. Effect of A. salinestris against Vigor Index of Maize, Sorghum, and Wheat
2.5. Effect of A. salinestris against Root Colonization of Cereal by Fusarium spp.
2.6. In Planta Antagonistic Activity of A. salinestris against Fusarium spp.
2.7. Statistical Analysis
3. Results
3.1. In Vitro Studies of A. salinestris against Fusarium spp.
3.1.1. Antagonistic Activity of A. salinestris Investigated by Dual-Culture Method
3.1.2. Effect of Azotobacter against Fusarium spp. and Infection Incidence in Cereals
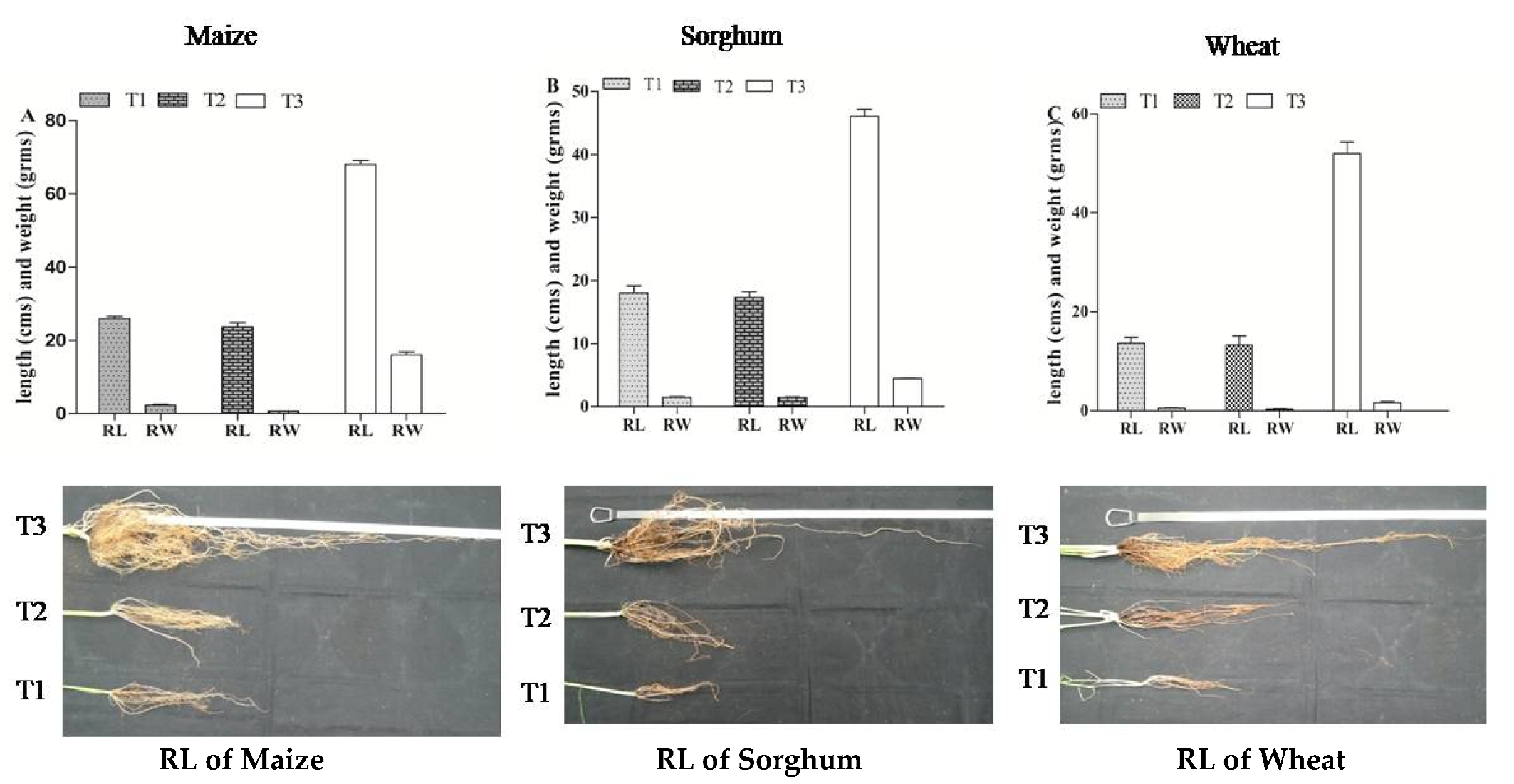
3.1.3. Effect of A. salinestris against Vigor Index of Maize, Sorghum, and Wheat
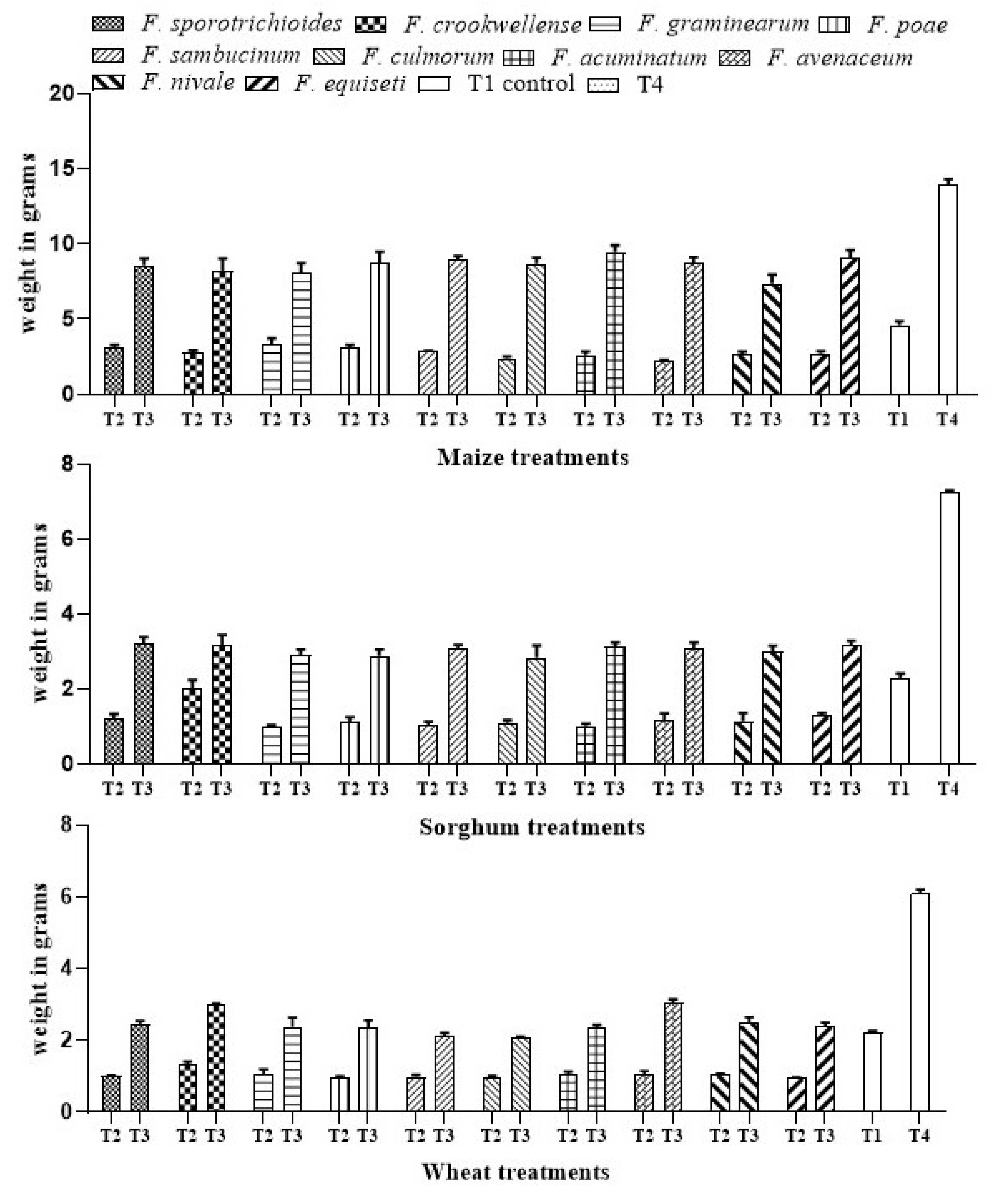
3.1.4. Effect of A. salinestris against Weight of Cereals under Different Treatments
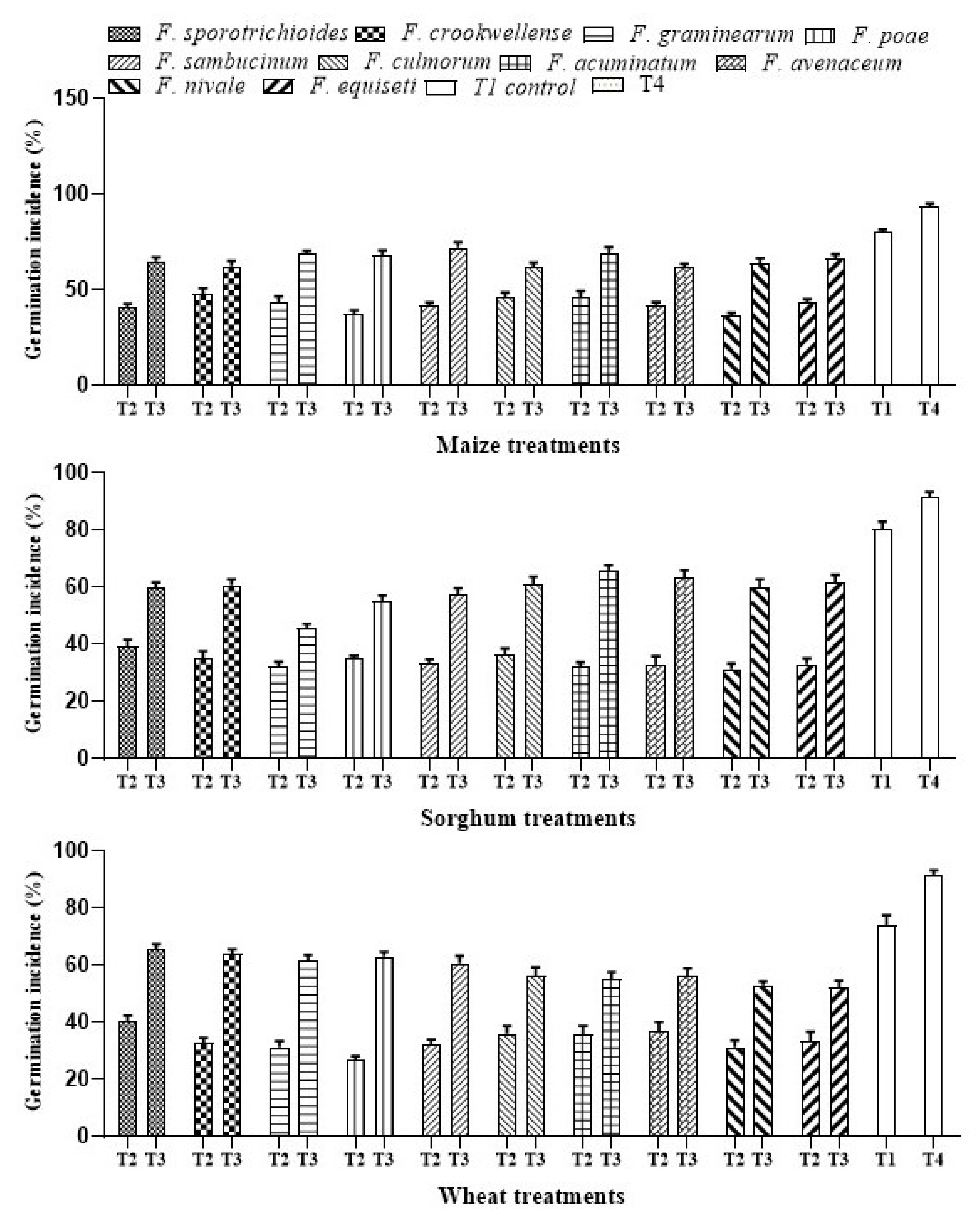
3.1.5. Effect of A. salinestris Treatment on Percentage of Germination in Cereal Grains
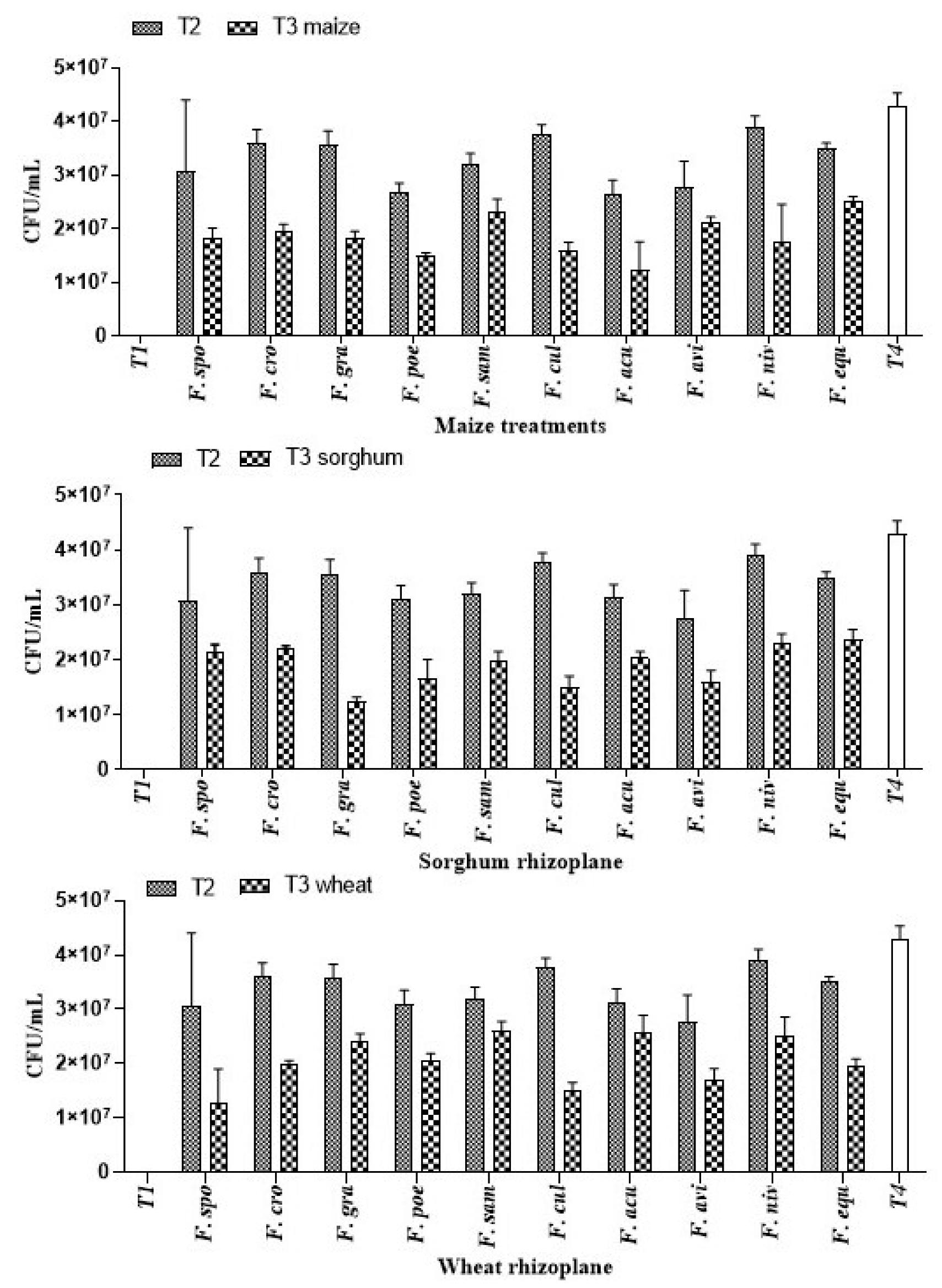
3.1.6. Effect of A. salinestris against Root Colonization of Cereals by Fusarium spp.
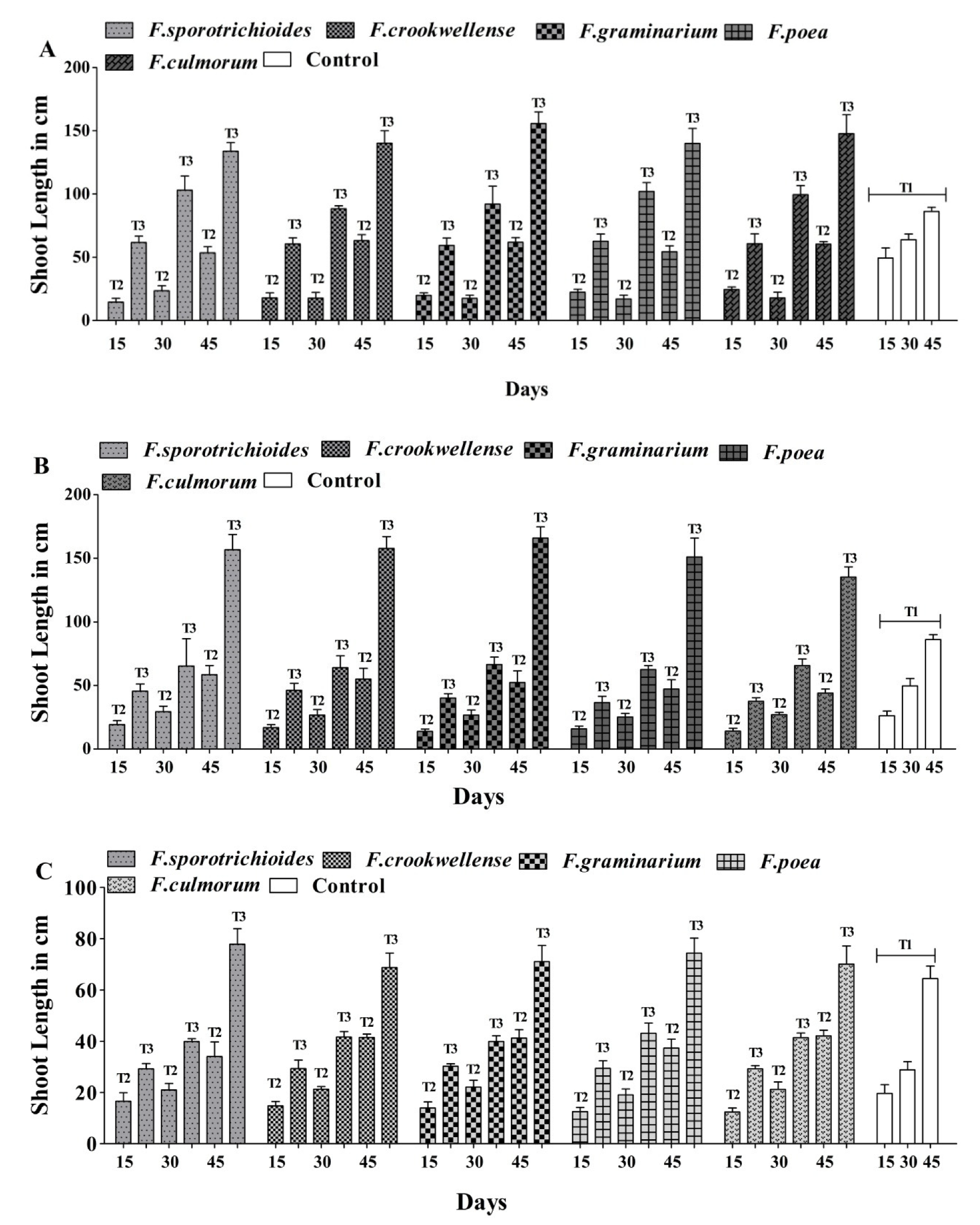
3.2. In Planta Studies of A. salinestris against Fusarium spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed consent statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bottalico, A. Fusarium diseases of cereals: Species complex and related mycotoxin profiles, in Europe. J. Plant Pathol. 1998, 80, 85–103. [Google Scholar]
- Murray, G.M.; Brennan, J.P. Estimating disease loss to the Australian wheat industry. Australas. Plant Pathol. 2009, 38, 558–570. [Google Scholar] [CrossRef]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Bensassi, F.; Zaied, C.; Abid, S.; Hajlaoui, M.R.; Bicha, H. Occurrence of deoxynivalenol in durum wheat in Tunisia. Food Control 2010, 21, 281–285. [Google Scholar] [CrossRef]
- Gelderblom, W.C.A.; Jaskiewicz, K.; Marasas, W.F.O.; Thiel, P.G. Fumonisins—Novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [CrossRef] [Green Version]
- Fandohan, P.; Hell, K.; Marasas, W.F.O.; Wingfield, M.J. Infection of maize by Fusarium species and contamination with fumonisin in Africa. Afr. J. Biotechnol. 2003, 2, 570–579. [Google Scholar]
- Chrestensen, C.M.; Aufmann, K.H.H. Grain Storage. The Role of Fungi in Quality Loss; University of Minnesota Press: Minneapolis, MN, USA, 1969; pp. 36–108. [Google Scholar]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef]
- Deepa, N.; Premila Achar, N.; Sreenivasa, M.Y. Current perspectives of biocontrol agents for management of Fusarium verticillioides and its fumonisin in cereals—A review. J. Fungi 2021, 7, 776. [Google Scholar]
- Deepa, N.; Adkar-Purushothama, C.R.; Sreenivasa, M.Y. Chapter 6: “Molecular Technologies for the Early Detection of Fungal Phytopathogens Associated with Cereal Crops”. Microbial Genomic Technologies to Combat the Problems of Farming Lands. In Microbes in Land Use Change Management; Elsevier: Amsterdam, The Netherlands, 2021; pp. 137–154. [Google Scholar]
- Ayala, S.; Prakasa Rao, E.V.S. Perspectives of soil fertility management with a focus on fertilizer use for crop productivity. Curr. Sci. 2002, 82, 797–807. [Google Scholar]
- Mousa, M.A.; Abo-Elyousr, K.A.; Abdel Alal, A.M.; Alshareef, N.O. Management Fusarium wilt disease in tomato by combnation of Bacillus amyloliquifaciens and peppermint oil. Agronomy 2021, 11, 2536. [Google Scholar] [CrossRef]
- Madriz-Ordeñana, K.; Pazarlar, S.; Jørgensen, H.J.L.; Nielsen, T.K.; Zhang, Y.; Nielsen, K.L.; Hansen, L.H.; Thordal-Christensen, H. The Bacillus cereus strain EC9 primers the plant immune system for superior biocontrol of F. oxysporum. Plants 2022, 11, 687. [Google Scholar] [CrossRef]
- de Fátima Dias Diniz, G.; Cota, L.V.; Figueiredo, J.E.F.; Aguiar, F.M.; da Silva, D.D.; de Paula Lana, U.G.; Dos Santos, V.L.; Marriel, I.E.; de Oliveira-Paiva, C.A. Antifungal activity of bacterial strains from maize silks against Fusarium. Arch. Microbiol. 2022, 81, 204. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, H.; Chennappa, G.; Rakesh, S.; Naik, M.K.; Amaresh, Y.S.; Sreenivasa, M.Y. Antifungal activity of Azotobacter nigricans against trichothecene producing Fusarium species associated with cereals. Food Sci. Biotechnol. 2016, 25, 1197–1204. [Google Scholar] [CrossRef]
- Aiyaz, M.; Divakar, S.T.; Nayaka, S.C.; Hariprasad, P.; Niranjana, S.R. Application of beneficial rhizospheric microbes for the mitigation of seed- borne mycotoxigenic fungal infection and mycotoxins in maize. Biocontrol Sci. Technol. 2015, 25, 1105–1119. [Google Scholar] [CrossRef]
- Bharadwaj, D.; Ansari, M.W.; Sahoo, R.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Punja, Z.K.; Rodriguez, G.; Tirajoh, A. Effects of Bacillus subtilis strain QST 713 and storage temperatures on post-harvest disease development on greenhouse tomatoes. Crop Prot. 2016, 84, 98–104. [Google Scholar] [CrossRef]
- Nagaraja, H.; Chennappa, G.; Poorna Chandra Rao, K.; Mahadev Prasad, G.; Sreenivasa, M.Y. Diversity of toxic and phytopathogenic Fusarium species occurring on cereals grown in Karnataka state, India. 3 Biotech 2016, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Chennappa, G.; Adkar-Purushotama, C.R.; Suraj, U.; Tamilvendan, K.; Sreenivasa, M.Y. Pesticide tolerant Azotobacter isolates from paddy growing areas of northern Karnataka, India. World J. Microbiol. Biotechnol. 2013, 30, 1–7. [Google Scholar] [CrossRef]
- Peraira, P.; Nesci, A.; Etcheverry, M.G. Efficacy of bacterial seed treatment for the control of Fusarium verticillioides in maize. BioControl 2009, 54, 103–111. [Google Scholar] [CrossRef]
- Daniel, F.; Castillo, H.; Maria, A. In Vitro Antagonist Action of Trichoderma Strains against Sclerotinia sclerotiorum and Sclerotium cepivorum. Am. J. Agric. Biol. Sci. 2011, 6, 410–417. [Google Scholar]
- Bragulat, M.R.; Martınez, E.; Castella, G.; Cabanes, F.J. Selective efficacy of culture media recommended for isolation and enumeration of Fusarium spp. J. Food Prot. 2004, 67, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing Professional: Hoboken, NJ, USA, 2006; pp. 8–240. [Google Scholar]
- Tang, A.; Harana, A.O.; Ab Majid, M.N.; Jalloh, M.B. Effects of selected functional bacteria on maize growth and nutrient use efficiency. Microorganisms 2020, 8, 854. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: A greenhouse trial. Geoderma 2005, 125, 155–166. [Google Scholar] [CrossRef]
- Aasfar, A.; Bargez, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Kadmiu, I.M. Nitrogen fixing Azotobacter species as potential soil biological enhancers for crops nutrition and yield stability. Front. Microbiol. 2021, 12, 354. [Google Scholar] [CrossRef]
- Cavaglieri, L.R.; Passone, A.; Etcheverry, M.G. Correlation between screening procedures to select root endophytes for biological control of Fusarium verticillioides in Zea mays L. Biol. Control 2004, 31, 259–267. [Google Scholar] [CrossRef]
- Chauhan, S.; Wadhwa, K.; Vasudeva, M.; Narula, N. Potential of Azotobacter spp. as biocontrol agents against Rhizoctonia solani and Fusarium oxysporum in cotton (Gossypium hirsutum), guar (Cyamopsis tetragonoloba) and tomato (Lycopersicum esculentum). Arch. Agron. Soil Sci. 2012, 58, 1365–1385. [Google Scholar] [CrossRef]
- Pereira, P.; Nesci, A.; Etcheverry, M. Effects of biocontrol agents on Fusarium verticillioides count and fumonisin content in the maize agroecosystem, Impact on rhizospheric bacterial and fungal groups. Biol. Control 2007, 42, 281–287. [Google Scholar] [CrossRef]
- Cavaglieri, L.; Orlando, J.; Rodríguez, M.I. Biocontrol of Bacillus subtilis against Fusarium verticillioides in vitro and at the maize root level. Res. Microbiol. 2005, 156, 748–754. [Google Scholar] [CrossRef]
- Kokalis-Burelle, N.; Kloepper, J.W.; Reddy, M.S. Plant growth-promoting rhizobacteria as transplant amendments and their effects on indigenous rhizosphere microorganisms. Appl. Soil Ecol. 2006, 31, 91–100. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S. Induced resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vessey, J. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Biari, A.; Gholami, A.; Rahmani, H.A. Growth promotion and enhanced nutrient uptake of maize (Zea maya L.) by application of plant growth promoting rhizobacteria in arid region of Iran. J. Biol. Sci. 2008, 8, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.; Nesci, A.; Castillo, C.; Etcheverry, M. Impact of bacterial biological control agents on fumonisin B1 content and Fusarium verticillioides infection of field-grown maize. Biol. Control 2010, 53, 258–266. [Google Scholar] [CrossRef]
- Egamberndiyeva, D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Gao, C.; El-Sauah, A.M.; Ali, D.F.I.; Hamoud, Y.A.; Shaghalah, H.; Sheteiuy, M.J. The integration of bio and organic fertilizers improve plant growth grain yield quality and metabolism of hybrid maize (Zea mays). Agronomy 2020, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Johri, B.N. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res. 2003, 158, 243–248. [Google Scholar] [CrossRef]
- Ona, O.; Smith, I.; Gysegom, P.; Bernaerts, K.; Impe, J.V.; Prinen, E.; Vanderleyden, J. Effect of pH on Indole-3-aceticacid (IAA) biosynthesis of Azospirillum brailense sp7. Symbiosis 2003, 35, 199–208. [Google Scholar]
- Kumar, B.; Dubey, R.C.; Maheshwari, D.K. Biocontrol of Macrophomina Phaseolina: Prospects and Constraints. In Microbial Diversity: Current Perspectives and Potential Applications; Satyanarayana, T., Jorhi, B.N., Eds.; International Research Publication House: New Delhi, India, 2005; pp. 471–492. [Google Scholar]
- Deepa, N.; Sreenivasa, M.Y. Chapter 13: “Biocontrol Strategies for Effective Management of Phytopathogenic Fungi Associated with Cereals”. In New and Future Developments in Microbial Biotechnology and Bioengineering-Microbial Biotechnology in Agro-Environmental Sustainability; Elsevier: Amsterdam, The Netherlands, 2019; pp. 177–189. [Google Scholar]
- Deepa, N.; Sreenivasa, M.Y. Chapter 9: “Sustainable Approaches for Biological Control of Mycotoxigenic Fungi and Mycotoxins in Cereals”. In Use of Microbial Bio-Fertilizers Technology in Agro-Environmental Sustainability- Recent Trends in Biological Control Mycotoxigenic Fungi in Cereals; Elsevier: Amsterdam, The Netherlands, 2019; pp. 149–161. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number |
|---|---|
| Fusarium sporotrichioides | KJ371098 |
| F. crookwellense | KJ371105 |
| F. graminearum | KJ 371099 |
| F. poae | KJ 371096 |
| F. sambucinum | KJ371095 |
| F. culmorum | KJ 371104 |
| F. acuminatum | KJ371100 |
| F. avenaceum | KJ371102 |
| F. nivale | KJ371097 |
| F. equiseti | KJ371094 |
| Treatment | Treatment Type |
|---|---|
| T1 | Sterile water |
| T2 | Suspension of each Fusarium spp. strain at approx. 107 spores mL−1. |
| T3 | Suspension of each Fusarium spp. strain at approx. 107 spores mL1 (T2) + 109 CFU/mL of A. salinestris. |
| T4 | A. salinestris cell suspension at approx. 109 CFU/mL. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagaraja, H.; Chennappa, G.; Deepa, N.; Naik, M.K.; Ajithkumar, K.; Amaresh, Y.S.; Achar, P.N.; Sreenivasa, M.Y. Antifungal Potential of Azotobacter salinestris Strain Azt 31 against Phytopathogenic Fusarium spp. Associated with Cereals. J. Fungi 2022, 8, 473. https://doi.org/10.3390/jof8050473
Nagaraja H, Chennappa G, Deepa N, Naik MK, Ajithkumar K, Amaresh YS, Achar PN, Sreenivasa MY. Antifungal Potential of Azotobacter salinestris Strain Azt 31 against Phytopathogenic Fusarium spp. Associated with Cereals. Journal of Fungi. 2022; 8(5):473. https://doi.org/10.3390/jof8050473
Chicago/Turabian StyleNagaraja, Hanumanthu, Gurikar Chennappa, Nagaraj Deepa, Manjunath Krishnappa Naik, Kadaiah Ajithkumar, Yatgal Sharanappa Amaresh, Premila N. Achar, and M. Y. Sreenivasa. 2022. "Antifungal Potential of Azotobacter salinestris Strain Azt 31 against Phytopathogenic Fusarium spp. Associated with Cereals" Journal of Fungi 8, no. 5: 473. https://doi.org/10.3390/jof8050473