
Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of an Azotobacter chroococcum Strain for Interaction with Oryza sativa in Presence of Piriformospora indica
2.2. Inoculation of O. sativa Seedlings with P. indica and W5
2.3. Nutrient Acquisition in O. sativa under Dual Inoculation
2.4. Effect of W5 and P. indica on the O. sativa mRNA Expression Specific to Roots
2.5. Impact of W5 on the P. indica-O. sativa Association
2.6. Identification of Plant Root Proteins and Secretory Proteins during Fungal-Bacterial Co-Inoculation
2.7. Preparation of Sample for LC-MS/MS
2.8. Mass Spectrometric Analysis of Peptide Mixtures
2.9. Data Processing
2.10. Statistical Analysis
3. Results
3.1. Interaction of P. indica with A. chroococcum Strains W5 and A41
3.2. Co-Inoculation Stimulates Phenotypic Traits in O. sativa
3.3. Co-Inoculation Contributes to Nutrient Acquisition of O. sativa
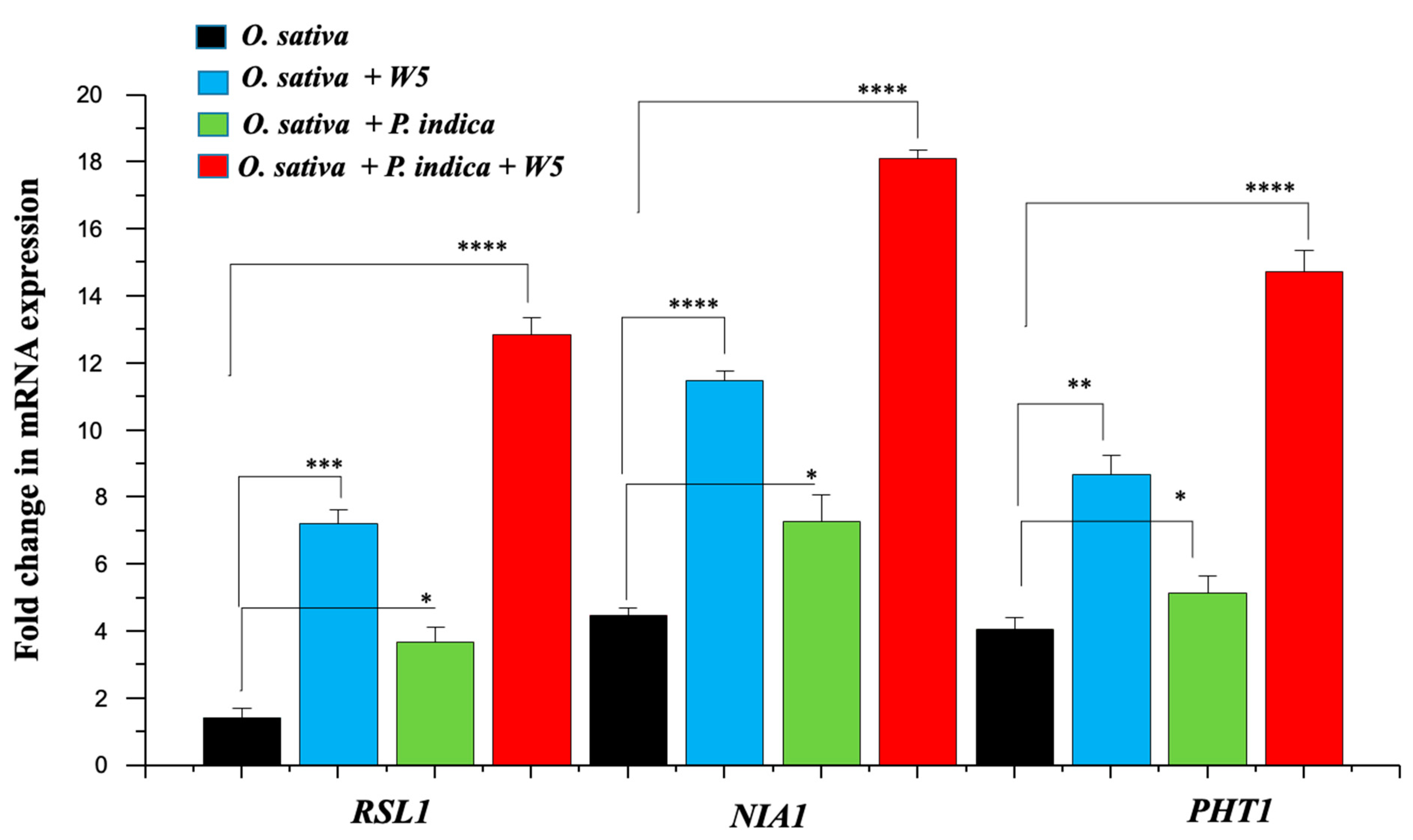
3.4. Impact of W5 and P. indica on the Selected O. sativa RNA Transcripts
3.5. W5 Stimulates Carbon (C), Nitrogen (N) and Phosphorous (P) Allocation by P. indica into O. sativa
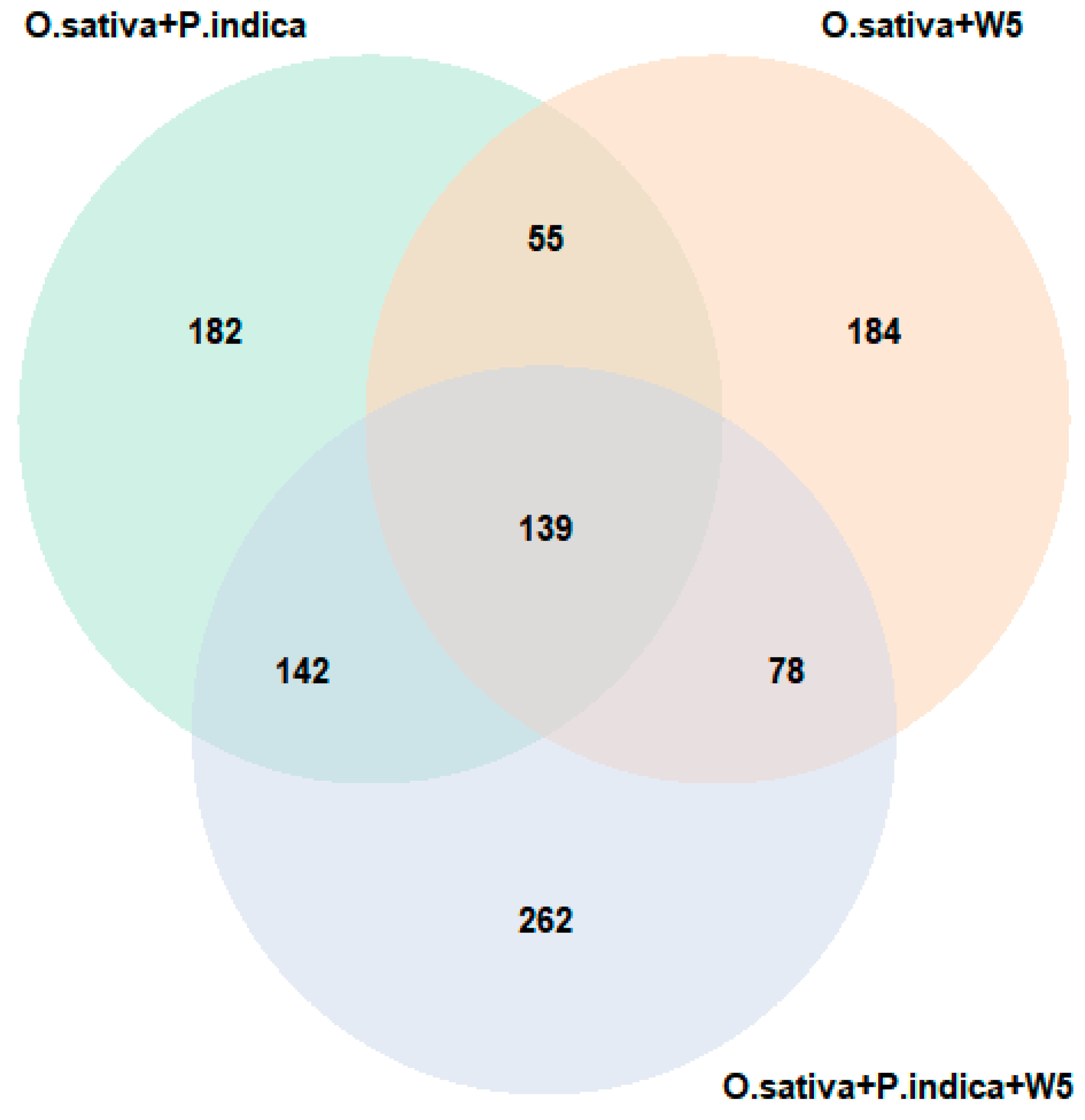
3.6. Identification of Differentially Abundant (DAP) Proteins in Individual and Co-Inoculation with P. indica and W5
3.7. Identification of DAP Proteins in P. indica in Presence of W5
4. Discussion
4.1. Effect of Microbes on Plant Growth
4.2. Enzyme Activity and Expression Analysis of Nutrition Acquisition Genes during Individual Inoculation and Co-Inoculation in O. sativa
4.3. Proteome Changes in P. indica during Inoculation with W5
4.4. Proteomic Changes in O. sativa after Inoculation with W5 and P. indica and in Combination
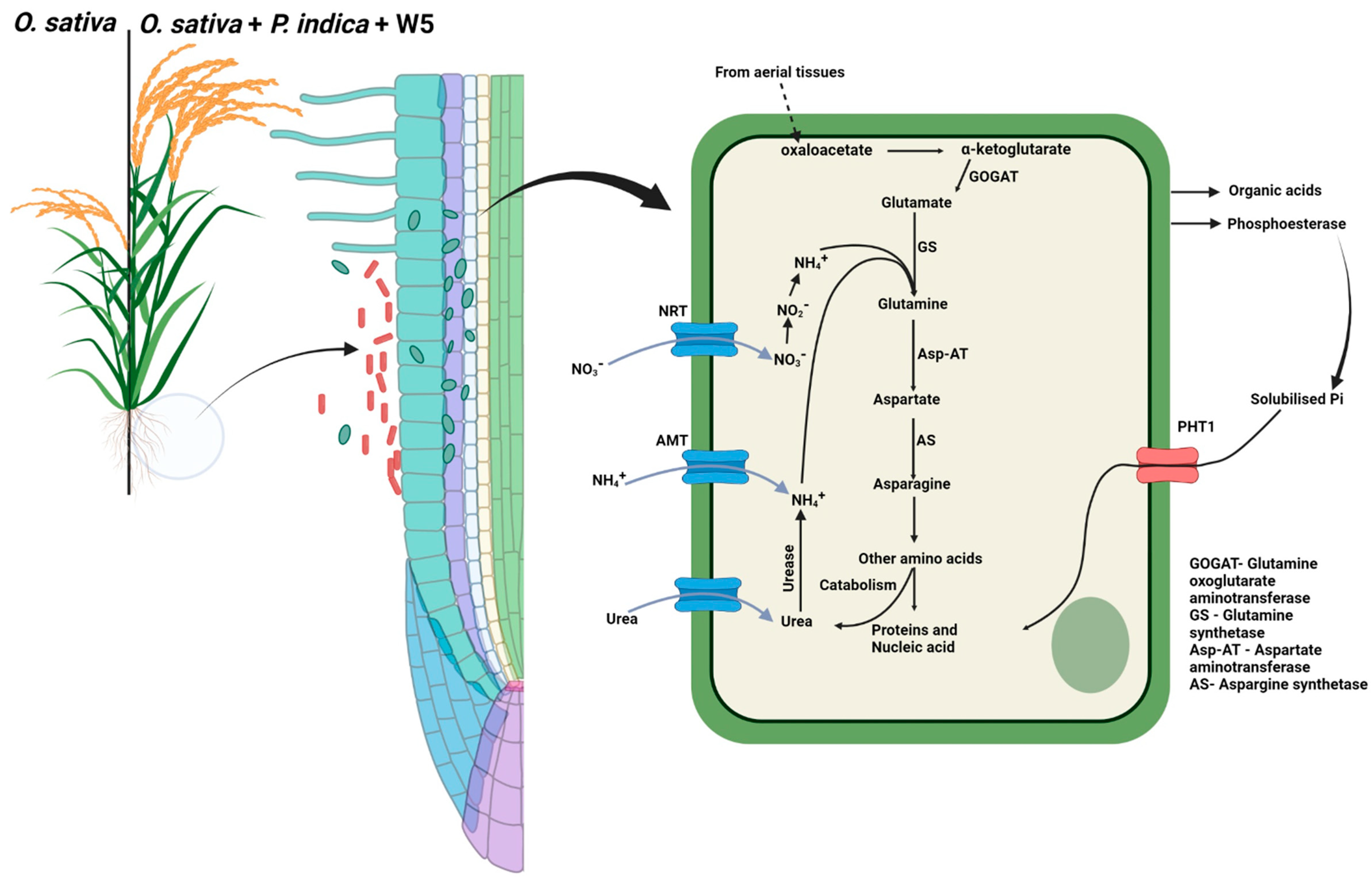
4.5. Mechanism of Nutrient Uptake and for Increased Growth in O. sativa
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brundrett, M.C. Coevolution of Roots and Mycorrhizas of Land Plants. New Phytol. 2002, 154, 275–304. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. The Enhancement of Plant Growth by Free-Living Bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Rao, A.; Tak, R. Influence of Mycorrhizal Fungi on the Growth of Different Tree Species and Their Nutrient Uptake in Gypsum Mine Spoil in India. Appl. Soil Ecol. 2001, 17, 279–284. [Google Scholar] [CrossRef]
- Bonfante, P. The Endless Tale of Endobacteria: A Conversation with Paola Bonfante. Trends Plant Sci. 2014, 19, 744–746. [Google Scholar] [CrossRef] [PubMed]
- Alabid, I.; Glaeser, S.P.; Kogel, K.-H. Endofungal Bacteria Increase Fitness of Their Host Fungi and Impact Their Association with Crop Plants. Curr. Issues Mol. Biol. 2019, 30, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Deveau, A.; Bonito, G.; Uehling, J.; Paoletti, M.; Becker, M.; Bindschedler, S.; Hacquard, S.; Hervé, V.; Labbé, J.; Lastovetsky, O.A.; et al. Bacterial–Fungal Interactions: Ecology, Mechanisms and Challenges. FEMS Microbiol. Rev. 2018, 42, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, M.G.A.; de Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.P.; Schlaeppi, K. A Widespread Plant-Fungal-Bacterial Symbiosis Promotes Plant Biodiversity, Plant Nutrition and Seedling Recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial Endophytic Bacteria-Serendipita Indica Interaction for Crop Enhancement and Resistance to Phytopathogens. Front. Microbiol. 2019, 10, 2888. [Google Scholar] [CrossRef]
- Van Overbeek, L.S.; Saikkonen, K. Impact of Bacterial-Fungal Interactions on the Colonization of the Endosphere. Trends Plant Sci. 2016, 21, 230–242. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial Interactions and Biocontrol in the Rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Yang, N.; Wen, Z.; Sun, X.; Chai, Y.; Ma, Z. Wheat Microbiome Bacteria Can Reduce Virulence of a Plant Pathogenic Fungus by Altering Histone Acetylation. Nat. Commun. 2018, 9, 3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.R. Bacteria with ACC Deaminase Can Promote Plant Growth and Help to Feed the World. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Ghignone, S.; Salvioli, A.; Anca, I.; Lumini, E.; Ortu, G.; Petiti, L.; Cruveiller, S.; Bianciotto, V.; Piffanelli, P.; Lanfranco, L.; et al. The Genome of the Obligate Endobacterium of an AM Fungus Reveals an Interphylum Network of Nutritional Interactions. ISME J. 2012, 6, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, U.; Ouziad, F.; Marner, F.-J.; Bothe, H. The Bacterium Paenibacillus Validus Stimulates Growth of the Arbuscular Mycorrhizal Fungus Glomus Intraradices up to the Formation of Fertile Spores. FEMS Microbiol. Lett. 2006, 254, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miransari, M. Arbuscular Mycorrhizal Fungi and Nitrogen Uptake. Arch. Microbiol. 2011, 193, 77–81. [Google Scholar] [CrossRef]
- Zhou, J.; Chai, X.; Zhang, L.; George, T.S.; Wang, F.; Feng, G. Different Arbuscular Mycorrhizal Fungi Cocolonizing on a Single Plant Root System Recruit Distinct Microbiomes. mSystems 2020, 5, e00929-20. [Google Scholar] [CrossRef]
- Wang, J.; Xu, C.; Sun, Q.; Xu, J.; Chai, Y.; Berg, G.; Cernava, T.; Ma, Z.; Chen, Y. Post-Translational Regulation of Autophagy Is Involved in Intra-Microbiome Suppression of Fungal Pathogens. Microbiome 2021, 9, 131. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A Review on the Plant Microbiome: Ecology, Functions, and Emerging Trends in Microbial Application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Varma, A.; Verma, S.; Sudha; Sahay, N.; Bütehorn, B.; Franken, P. Piriformospora Indica, a Cultivable Plant-Growth-Promoting Root Endophyte. Appl. Environ. Microbiol. 1999, 65, 2741–2744. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora Indica: Potential and Significance in Plant Stress Tolerance. Front. Microbiol. 2016, 7, 332. [Google Scholar] [CrossRef] [Green Version]
- Stein, E.; Molitor, A.; Kogel, K.-H.; Waller, F. Systemic Resistance in Arabidopsis Conferred by the Mycorrhizal Fungus Piriformospora Indica Requires Jasmonic Acid Signaling and the Cytoplasmic Function of NPR1. Plant Cell Physiol. 2008, 49, 1747–1751. [Google Scholar] [CrossRef] [PubMed]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.-H.; Schäfer, P.; Schwarczinger, I.; et al. Salt Tolerance of Barley Induced by the Root Endophyte Piriformospora Indica Is Associated with a Strong Increase in Antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, P.; Pfiffi, S.; Voll, L.M.; Zajic, D.; Chandler, P.M.; Waller, F.; Scholz, U.; Pons-Kühnemann, J.; Sonnewald, S.; Sonnewald, U.; et al. Manipulation of Plant Innate Immunity and Gibberellin as Factor of Compatibility in the Mutualistic Association of Barley Roots with Piriformospora Indica. Plant J. Cell Mol. Biol. 2009, 59, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Franken, P. The Plant Strengthening Root Endophyte Piriformospora Indica: Potential Application and the Biology Behind. Appl. Microbiol. Biotechnol. 2012, 96, 1455–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wu, C.; Oelmüller, R.; Zhang, W. Role of Phytohormones in Piriformospora Indica-Induced Growth Promotion and Stress Tolerance in Plants: More Questions Than Answers. Front. Microbiol. 2018, 9, 1646. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Schmid, M.; Rothballer, M.; Hause, G.; Zuccaro, A.; Imani, J.; Kämpfer, P.; Domann, E.; Schäfer, P.; Hartmann, A.; et al. Detection and Identification of Bacteria Intimately Associated with Fungi of the Order Sebacinales. Cell. Microbiol. 2008, 10, 2235–2246. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Imani, J.; Alabid, I.; Guo, H.; Kumar, N.; Kämpfer, P.; Hardt, M.; Blom, J.; Goesmann, A.; Rothballer, M.; et al. Non-Pathogenic Rhizobium Radiobacter F4 Deploys Plant Beneficial Activity Independent of Its Host Piriformospora Indica. ISME J. 2016, 10, 871–884. [Google Scholar] [CrossRef]
- Kumar Bhuyan, S.; Bandyopadhyay, P.; Kumar, P.; Kumar Mishra, D.; Prasad, R.; Kumari, A.; Chandra Upadhyaya, K.; Varma, A.; Kumar Yadava, P. Interaction of Piriformospora Indica with Azotobacter Chroococcum. Sci. Rep. 2015, 5, 13911. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, P.; Bhuyan, S.K.; Yadava, P.K.; Varma, A.; Tuteja, N. Emergence of Plant and Rhizospheric Microbiota as Stable Interactomes. Protoplasma 2017, 254, 617–626. [Google Scholar] [CrossRef]
- Toro, M.; Azcon, R.; Barea, J. Improvement of Arbuscular Mycorrhiza Development by Inoculation of Soil with Phosphate-Solubilizing Rhizobacteria To Improve Rock Phosphate Bioavailability ((Sup32)P) and Nutrient Cycling. Appl. Environ. Microbiol. 1997, 63, 4408–4412. [Google Scholar] [CrossRef] [Green Version]
- Jahandideh Mahjen Abadi, V.A.; Sepehri, M.; Khatabi, B.; Rezaei, M. Alleviation of Zinc Deficiency in Wheat Inoculated with Root Endophytic Fungus Piriformospora Indica and Rhizobacterium Pseudomonas Putida. Rhizosphere 2021, 17, 100311. [Google Scholar] [CrossRef]
- Arora, M.; Saxena, P.; Abdin, M.Z.; Varma, A. Interaction between Piriformospora Indica and Azotobacter Chroococcum Governs Better Plant Physiological and Biochemical Parameters in Artemisia annua L. Plants Grown under in Vitro Conditions. Symbiosis 2018, 75, 103–112. [Google Scholar] [CrossRef]
- Silletti, S.; Di Stasio, E.; Van Oosten, M.J.; Ventorino, V.; Pepe, O.; Napolitano, M.; Marra, R.; Woo, S.L.; Cirillo, V.; Maggio, A. Biostimulant Activity of Azotobacter Chroococcum and Trichoderma Harzianum in Durum Wheat under Water and Nitrogen Deficiency. Agronomy 2021, 11, 380. [Google Scholar] [CrossRef]
- Beltayef, H.; Melki, M.; Saidi, W.; Hajri, R.; Cruz, C.; Muscolo, A.; ben Youness, M. Potential Piriformospora Indica Effect on Growth and Mineral Nutrition of Phaseolus Vulgaris Crop under Low Phosphorus Intake. J. Plant Nutr. 2021, 44, 498–507. [Google Scholar] [CrossRef]
- Hill, T.; Kafer, E. Improved Protocols for Aspergillus Minimal Medium: Trace Element and Minimal Medium Salt Stock Solutions. Fungal Genet. Rep. 2001, 48, 20–21. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Boscari, A.; Horta Araújo, N.; Maucourt, M.; Hanchi, M.; Bernillon, S.; Rolin, D.; Puppo, A.; Brouquisse, R. Plant Nitrate Reductases Regulate Nitric Oxide Production and Nitrogen-Fixing Metabolism During the Medicago Truncatula–Sinorhizobium Meliloti Symbiosis. Front. Plant Sci. 2020, 11, 1313. [Google Scholar] [CrossRef]
- Fellbaum, C.R.; Gachomo, E.W.; Beesetty, Y.; Choudhari, S.; Strahan, G.D.; Pfeffer, P.E.; Kiers, E.T.; Bücking, H. Carbon Availability Triggers Fungal Nitrogen Uptake and Transport in Arbuscular Mycorrhizal Symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 2666–2671. [Google Scholar] [CrossRef] [Green Version]
- Khade, S.W.; Rodrigues, B.F.; Sharma, P.K. Arbuscular Mycorrhizal Status and Root Phosphatase Activities in Vegetative Carica Papaya L. Varieties. Acta Physiol. Plant. 2010, 32, 565–574. [Google Scholar] [CrossRef]
- Raorane, M.L.; Pabuayon, I.M.; Varadarajan, A.R.; Mutte, S.K.; Kumar, A.; Treumann, A.; Kohli, A. Proteomic Insights into the Role of the Large-Effect QTL QDTY12.1for Rice Yield under Drought. Mol. Breed. 2015, 35, 139. [Google Scholar] [CrossRef]
- Li, M.; Tu, S.; Li, Z.; Tan, F.; Liu, J.; Wang, Q.; Zhang, Y.; Xu, J.; Zhang, Y.; Zhou, F.; et al. MAP: Model-Based Analysis of Proteomic Data to Detect Proteins with Significant Abundance Changes. Cell Discov. 2019, 5, 40. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Priebe, S.; Linde, J.; Albrecht, D.; Guthke, R.; Brakhage, A.A. FungiFun: A Web-Based Application for Functional Categorization of Fungal Genes and Proteins. Fungal Genet. Biol. 2011, 48, 353–358. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Love, J.; Selker, R.; Marsman, M.; Jamil, T.; Dropmann, D.; Verhagen, J.; Ly, A.; Gronau, Q.F.; Šmíra, M.; Epskamp, S.; et al. JASP: Graphical Statistical Software for Common Statistical Designs. J. Stat. Softw. 2019, 88, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rani, M.; Raj, S.; Dayaman, V.; Kumar, M.; Dua, M.; Johri, A.K. Functional Characterization of a Hexose Transporter from Root Endophyte Piriformospora Indica. Front. Microbiol. 2016, 7, 1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blilou, I.; Bueno, P.; Ocampo, J.A.; García-Garrido, J.M. Induction of Catalase and Ascorbate Peroxidase Activities in Tobacco Roots Inoculated with the Arbuscular Mycorrhizal Glomus Mosseae. Mycol. Res. 2000, 104, 722–725. [Google Scholar] [CrossRef]
- Sepehri, M.; Ghaffari, M.R.; Khayam Nekoui, M.; Sarhadi, E.; Moghadam, A.; Khatabi, B.; Hosseini Salekdeh, G. Root Endophytic Fungus Serendipita Indica Modulates Barley Leaf Blade Proteome by Increasing the Abundance of Photosynthetic Proteins in Response to Salinity. J. Appl. Microbiol. 2021, 131, 1870–1889. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pallavi; Chugh, C.; Seem, K.; Kumar, S.; Vinod, K.K.; Mohapatra, T. Characterization of Contrasting Rice (Oryza Sativa L.) Genotypes Reveals the Pi-Efficient Schema for Phosphate Starvation Tolerance. BMC Plant Biol. 2021, 21, 282. [Google Scholar] [CrossRef] [PubMed]
- Svistoonoff, S.; Creff, A.; Reymond, M.; Sigoillot-Claude, C.; Ricaud, L.; Blanchet, A.; Nussaume, L.; Desnos, T. Root Tip Contact with Low-Phosphate Media Reprograms Plant Root Architecture. Nat. Genet. 2007, 39, 792–796. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Wakayama, M.; Aoki, N.; Komatsu, S.; Ichikawa, H.; Hirochika, H.; Ohsugi, R. Overexpression of a Calcium-Dependent Protein Kinase Gene Enhances Growth of Rice under Low-Nitrogen Conditions. Plant Biotechnol. 2010, 27, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Kim, S.G.; Kang, K.Y.; Kim, J.-G.; Park, S.-R.; Gupta, R.; Kim, Y.H.; Wang, Y.; Kim, S.T. Overexpression of a Pathogenesis-Related Protein 10 Enhances Biotic and Abiotic Stress Tolerance in Rice. Plant Pathol. J. 2016, 32, 552–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, I.; Eid, K.E.; Abbas, M.H.H.; Salem, A.A.; Ahmed, N.; Ali, M.; Shah, G.M.; Fang, C. Use of Plant Growth Promoting Rhizobacteria (PGPR) and Mycorrhizae to Improve the Growth and Nutrient Utilization of Common Bean in a Soil Infected with White Rot Fungi. Ecotoxicol. Environ. Saf. 2019, 171, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, K.; Chaudhary, R.; Sarwar, A.; Ahmad, B.; Gul, A.; Hano, C.; Abbasi, B.H.; Anjum, S. Melatonin as Master Regulator in Plant Growth, Development and Stress Alleviator for Sustainable Agricultural Production: Current Status and Future Perspectives. Sustainability 2021, 13, 294. [Google Scholar] [CrossRef]
- Igarashi, D.; Tsuchida, H.; Miyao, M.; Ohsumi, C. Glutamate: Glyoxylate Aminotransferase Modulates Amino Acid Content during Photorespiration1. Plant Physiol. 2006, 142, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Zecchin, V.J.S.; Ikeda, A.C.; Mógor, Á.F. Contribution of Plant Growth-Promoting Bacteria to the Maize Yield. In Advances in PGPR Research; Singh, H.B., Sarma, B.K., Keswani, C., Eds.; CABI: Wallingford, UK, 2017; pp. 234–245. ISBN 978-1-78639-032-5. [Google Scholar]
- Pandey, A.; Sharma, E.; Palni, L.M.S. Influence of Bacterial Inoculation on Maize in Upland Farming Systems of the Sikkim Himalaya. Soil Biol. Biochem. 1998, 30, 379–384. [Google Scholar] [CrossRef]
- Velmourougane, K.; Prasanna, R.; Singh, S.; Chawla, G.; Kumar, A.; Saxena, A.K. Modulating Rhizosphere Colonisation, Plant Growth, Soil Nutrient Availability and Plant Defense Enzyme Activity through Trichoderma Viride-Azotobacter Chroococcum Biofilm Inoculation in Chickpea. Plant Soil 2017, 421, 157–174. [Google Scholar] [CrossRef]
- Velmourougane, K.; Prasanna, R.; Chawla, G.; Nain, L.; Kumar, A.; Saxena, A.K. Trichoderma-Azotobacter Biofilm Inoculation Improves Soil Nutrient Availability and Plant Growth in Wheat and Cotton. J. Basic Microbiol. 2019, 59, 632–644. [Google Scholar] [CrossRef]
- Kilam, D.; Saifi, M.; Abdin, M.Z.; Agnihotri, A.; Varma, A. Combined Effects of Piriformospora Indica and Azotobacter Chroococcum Enhance Plant Growth, Antioxidant Potential and Steviol Glycoside Content in Stevia Rebaudiana. Symbiosis 2015, 66, 149–156. [Google Scholar] [CrossRef]
- Xu, F.; Liao, H.; Zhang, Y.; Yao, M.; Liu, J.; Sun, L.; Zhang, X.; Yang, J.; Wang, K.; Wang, X.; et al. Coordination of Root Auxin with the Fungus Piriformospora Indica and Bacterium Bacillus Cereus Enhances Rice Rhizosheath Formation under Soil Drying. ISME J. 2022, 16, 801–811. [Google Scholar] [CrossRef]
- Tahiri, A.; Raklami, A.; Bechtaoui, N.; Anli, M.; Boutasknit, A.; Oufdou, K.; Meddich, A. Beneficial Effects of Plant Growth Promoting Rhizobacteria, Arbuscular Mycorrhizal Fungi and Compost on Lettuce (Lactuca Sativa) Growth Under Field Conditions. Gesunde Pflanz. 2022, 74, 219–235. [Google Scholar] [CrossRef]
- Dabral, S.; Saxena, S.C.; Choudhary, D.K.; Bandyopadhyay, P.; Sahoo, R.K.; Tuteja, N.; Nath, M. Synergistic Inoculation of Azotobacter Vinelandii and Serendipita Indica Augmented Rice Growth. Symbiosis 2020, 81, 139–148. [Google Scholar] [CrossRef]
- Sędzielewska, K.A.; Böer, E.; Bellebna, C.; Wartmann, T.; Bode, R.; Melzer, M.; Baronian, K.; Kunze, G. Role of the AFRD1-Encoded Fumarate Reductase in Hypoxia and Osmotolerance in Arxula Adeninivorans. FEMS Yeast Res. 2012, 12, 924–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Bueno, G.; Hernández, A.; López-Lluch, G.; Pérez-Castiñeira, J.R.; Navas, P.; Serrano, A. Inorganic Pyrophosphatase Defects Lead to Cell Cycle Arrest and Autophagic Cell Death through NAD+ Depletion in Fermenting Yeast. J. Biol. Chem. 2013, 288, 13082–13092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The Interaction between Arbuscular Mycorrhizal Fungi and Endophytic Bacteria Enhances Plant Growth of Acacia Gerrardii under Salt Stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäder, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A.; et al. Inoculation of Root Microorganisms for Sustainable Wheat–Rice and Wheat–Black Gram Rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Salvioli, A.; Ghignone, S.; Novero, M.; Navazio, L.; Venice, F.; Bagnaresi, P.; Bonfante, P. Symbiosis with an Endobacterium Increases the Fitness of a Mycorrhizal Fungus, Raising Its Bioenergetic Potential. ISME J. 2016, 10, 130–144. [Google Scholar] [CrossRef]
- Nizam, S.; Qiang, X.; Wawra, S.; Nostadt, R.; Getzke, F.; Schwanke, F.; Dreyer, I.; Langen, G.; Zuccaro, A. Serendipita Indica E5’NT Modulates Extracellular Nucleotide Levels in the Plant Apoplast and Affects Fungal Colonization. EMBO Rep. 2019, 20, e47430. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional Roles of Melatonin in Plants, and Perspectives in Nutritional and Agricultural Science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015; ISBN 1-60535-353-1. [Google Scholar]
- Chen, X.; Sun, M.; Chong, S.; Si, J.; Wu, L. Transcriptomic and Metabolomic Approaches Deepen Our Knowledge of Plant–Endophyte Interactions. Front. Plant Sci. 2022, 12, 700200. [Google Scholar] [CrossRef]
- Schlesinger, W.H. On the Fate of Anthropogenic Nitrogen. Proc. Natl. Acad. Sci. USA 2009, 106, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and Eutrophication: Where Do We Go from Here? Sustainability 2014, 6, 5853–5875. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandyopadhyay, P.; Yadav, B.G.; Kumar, S.G.; Kumar, R.; Kogel, K.-H.; Kumar, S. Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa. J. Fungi 2022, 8, 453. https://doi.org/10.3390/jof8050453
Bandyopadhyay P, Yadav BG, Kumar SG, Kumar R, Kogel K-H, Kumar S. Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa. Journal of Fungi. 2022; 8(5):453. https://doi.org/10.3390/jof8050453
Chicago/Turabian StyleBandyopadhyay, Prasun, Bal Govind Yadav, Srinivasan Ganesh Kumar, Rahul Kumar, Karl-Heinz Kogel, and Shashi Kumar. 2022. "Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa" Journal of Fungi 8, no. 5: 453. https://doi.org/10.3390/jof8050453