
Enhanced Production, Cloning, and Expression of a Xylanase Gene from Endophytic Fungal Strain Trichoderma harzianum kj831197.1: Unveiling the In Vitro Anti-Fungal Activity against Phytopathogenic Fungi

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation of Xylanase Producing Strain from the Sugar Beet Plant
2.3. Identification of the Fungal Isolate
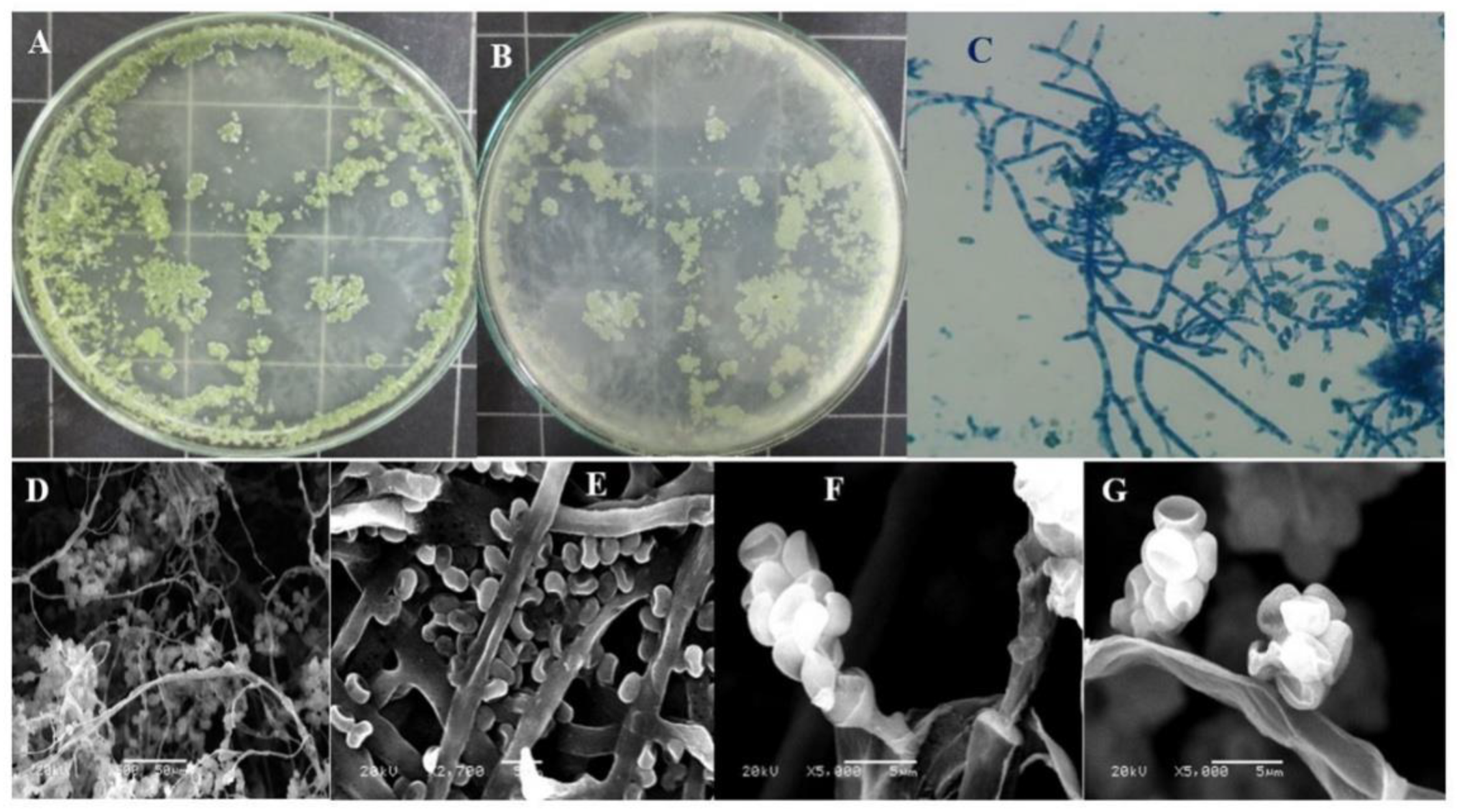
2.3.1. Morphological Identification
2.3.2. Molecular Identification
Genomic DNA Extraction and Primer Designing
PCR and DNA Sequencing
2.4. Bacterial Strain and Culture Circumstances
2.5. Plasmids
2.6. Fungal Growth and Optimization of Xylanase Productivity
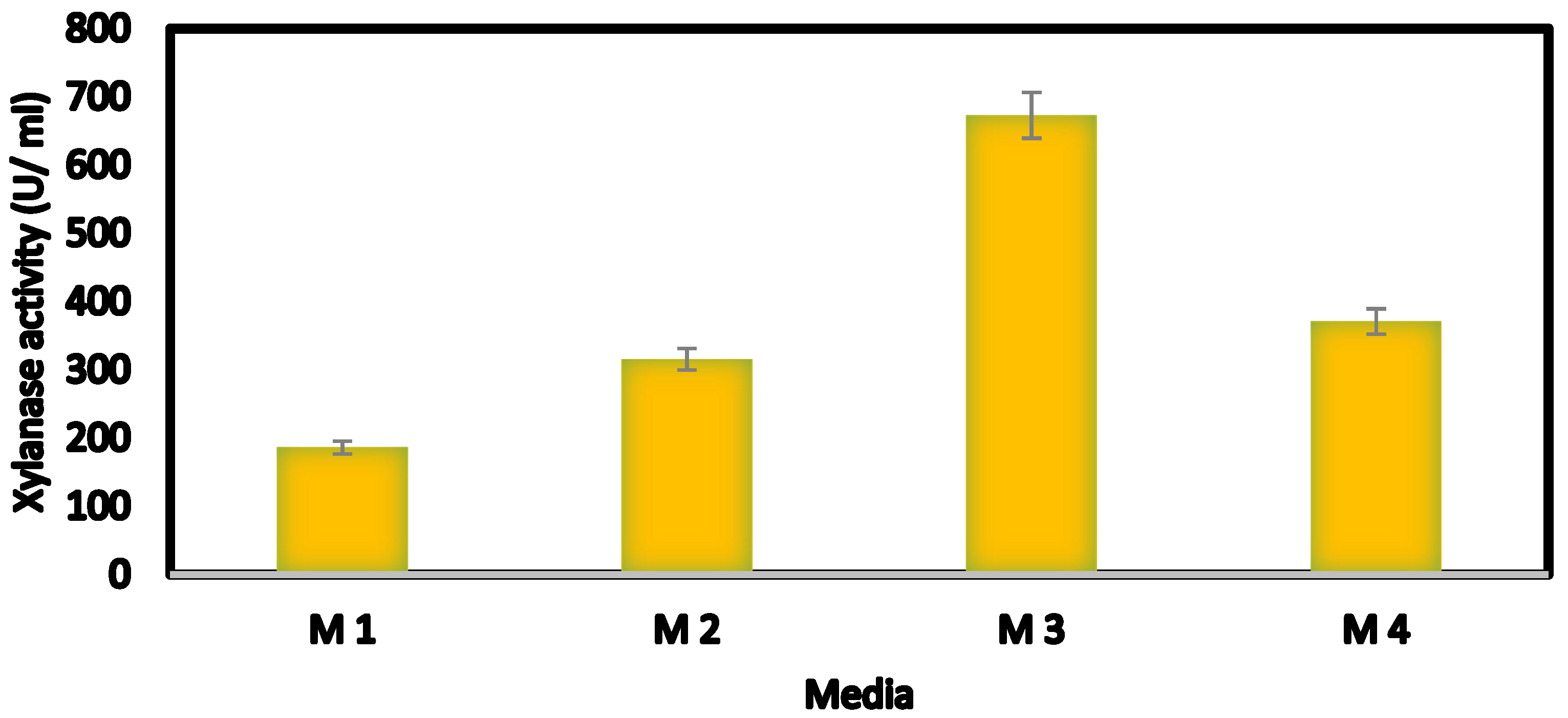
2.6.1. Selection of the Best Medium Supporting Highest Enzyme Activity
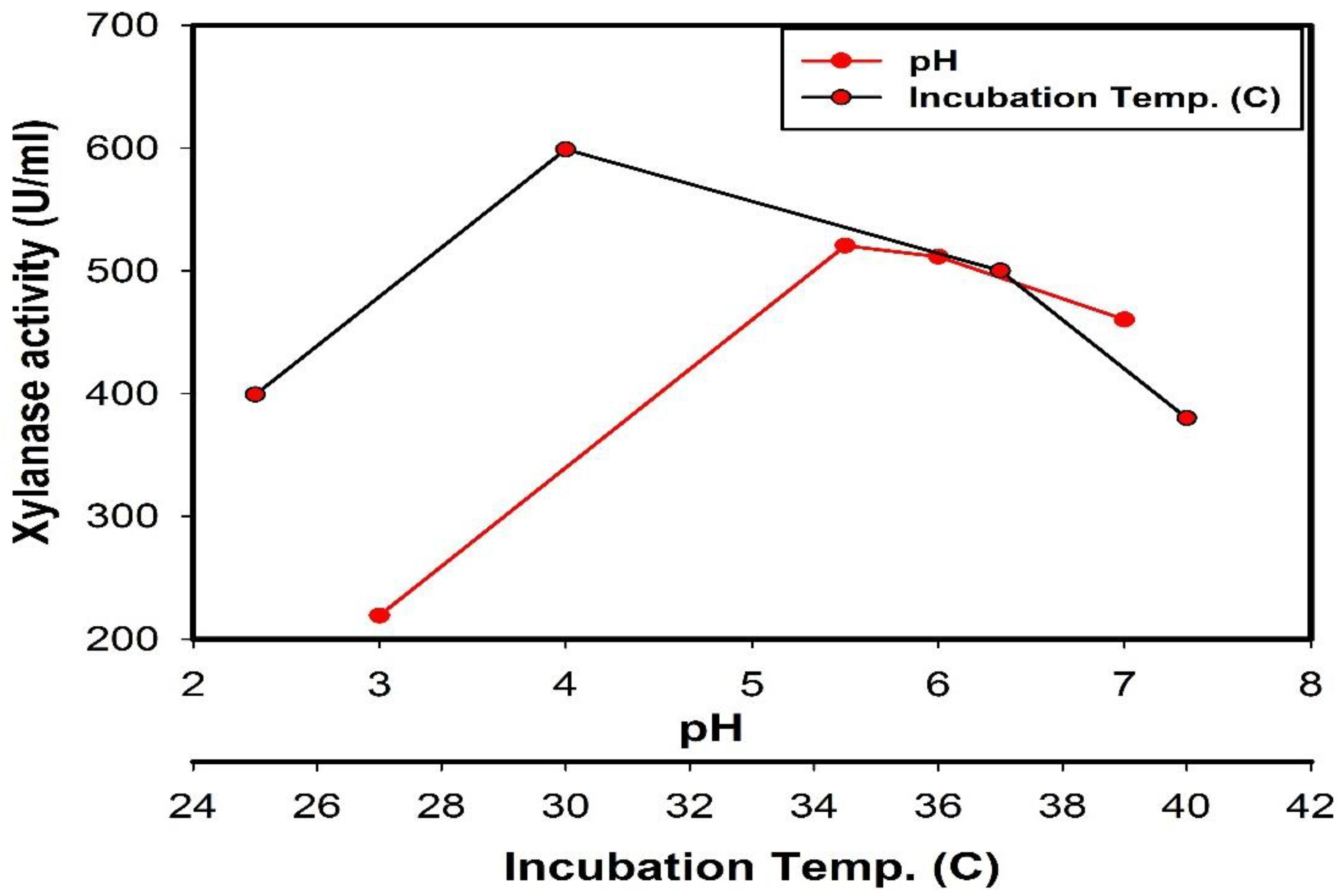
2.6.2. Effect of Different pH Values
2.6.3. Effect of Incubation Conditions (Shaking and Static Manner)
2.6.4. Effect of Different Incubation Temperature
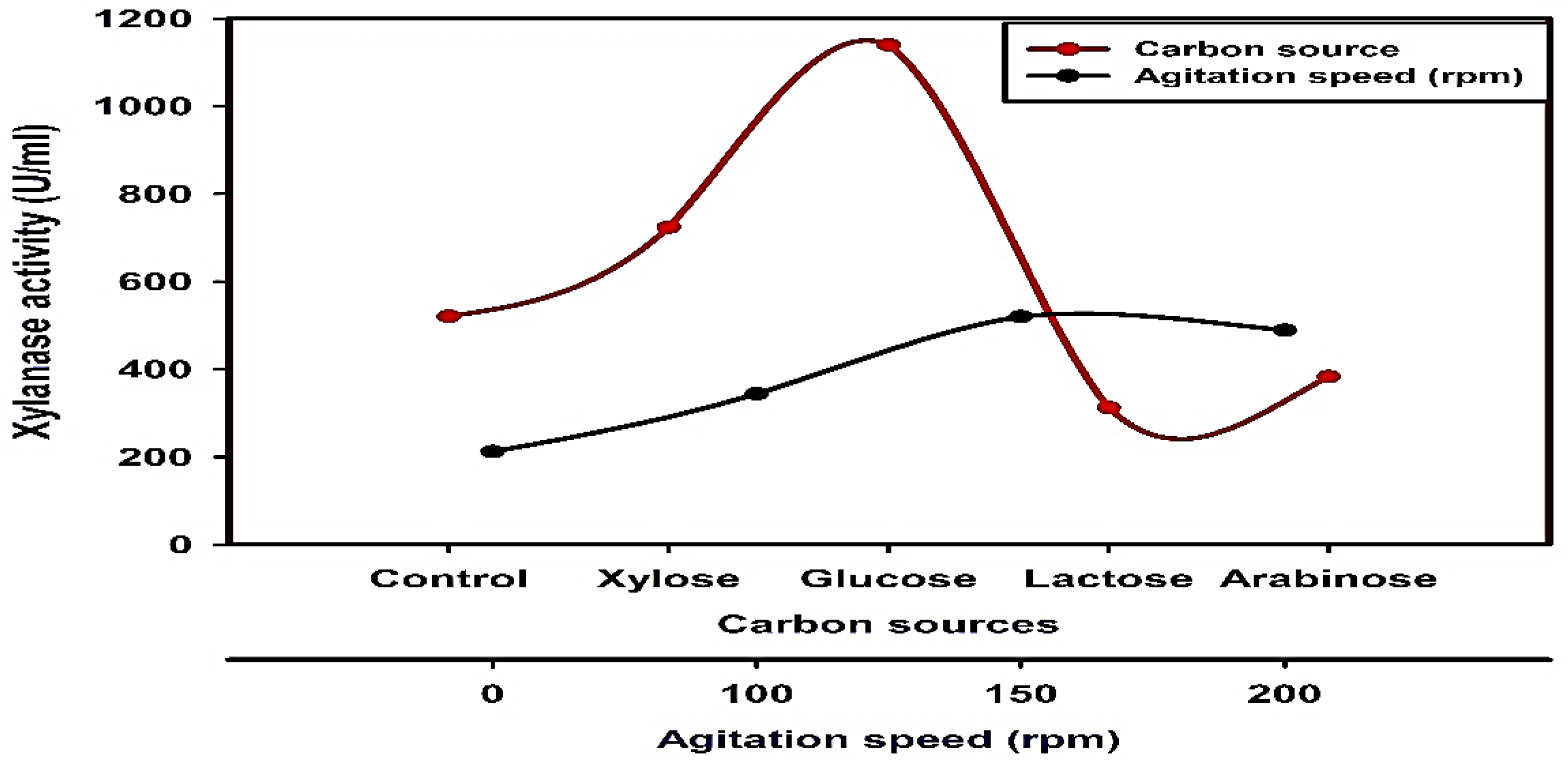
2.6.5. Effect of Carbon Sources
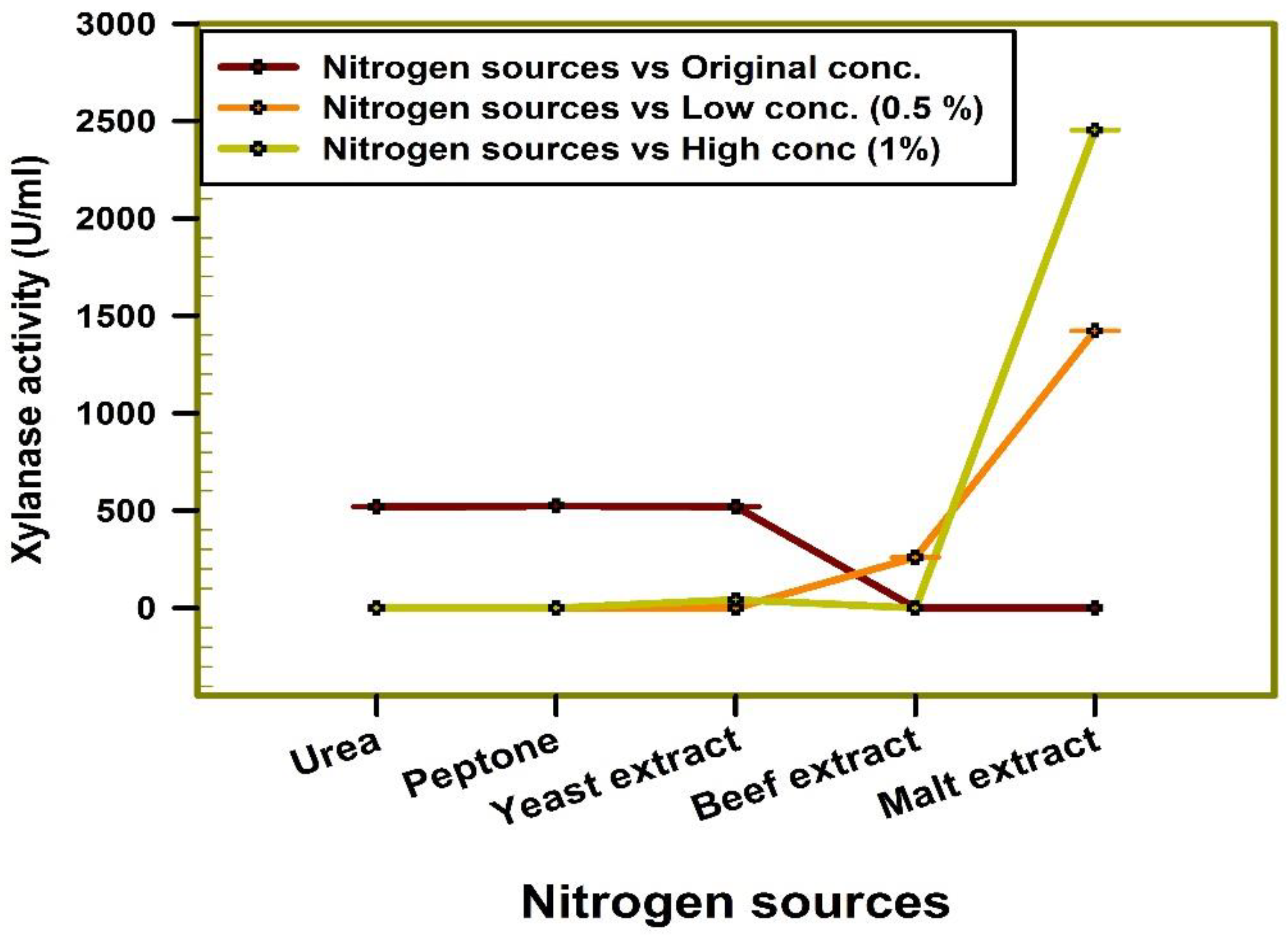
2.6.6. Effect of Nitrogen Sources
2.6.7. Effect of Different Inducers
2.6.8. Effect of Tween 80 as an Additive
2.6.9. Effect of Agricultural Wastes
2.7. Purification of Crude Xylanase
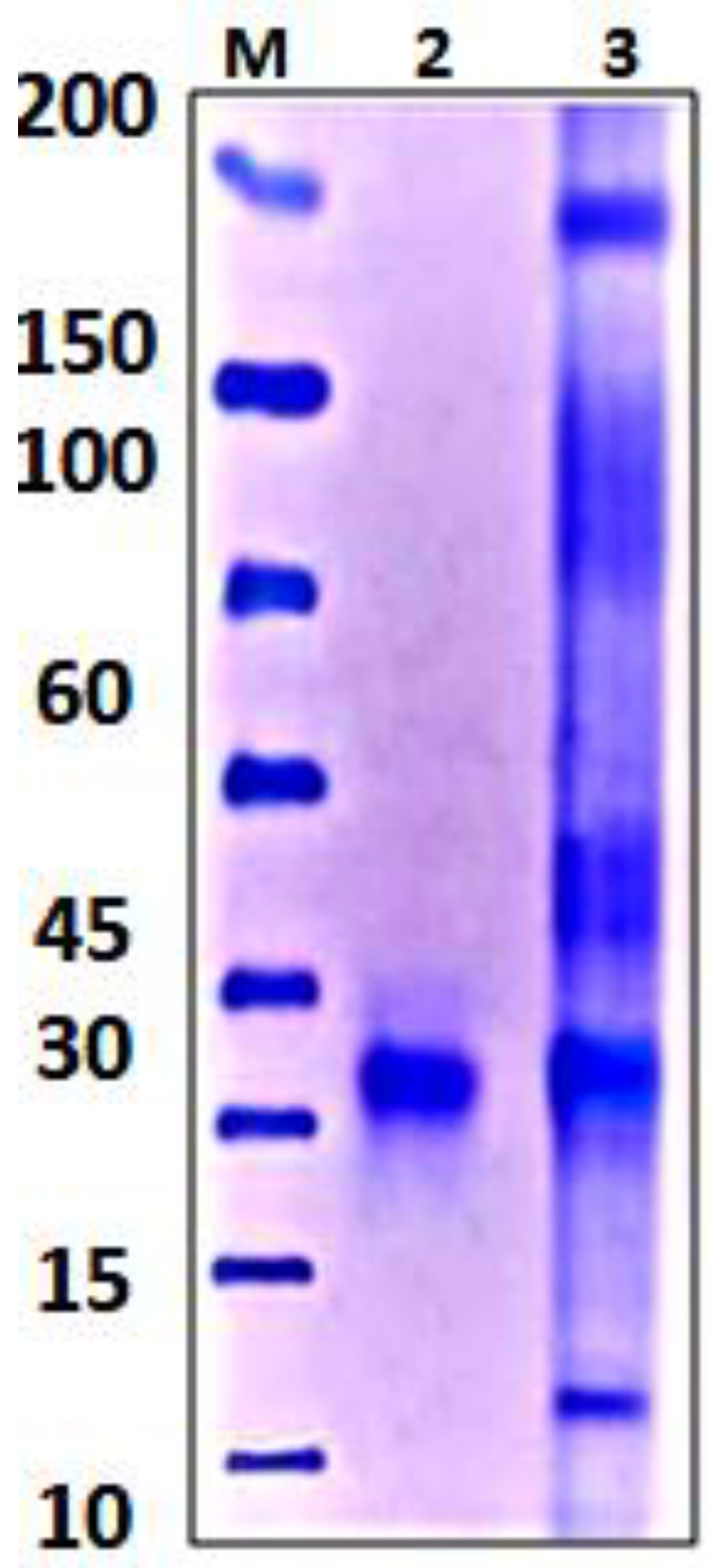
2.7.1. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.7.2. Characterization of Purified Xylanase
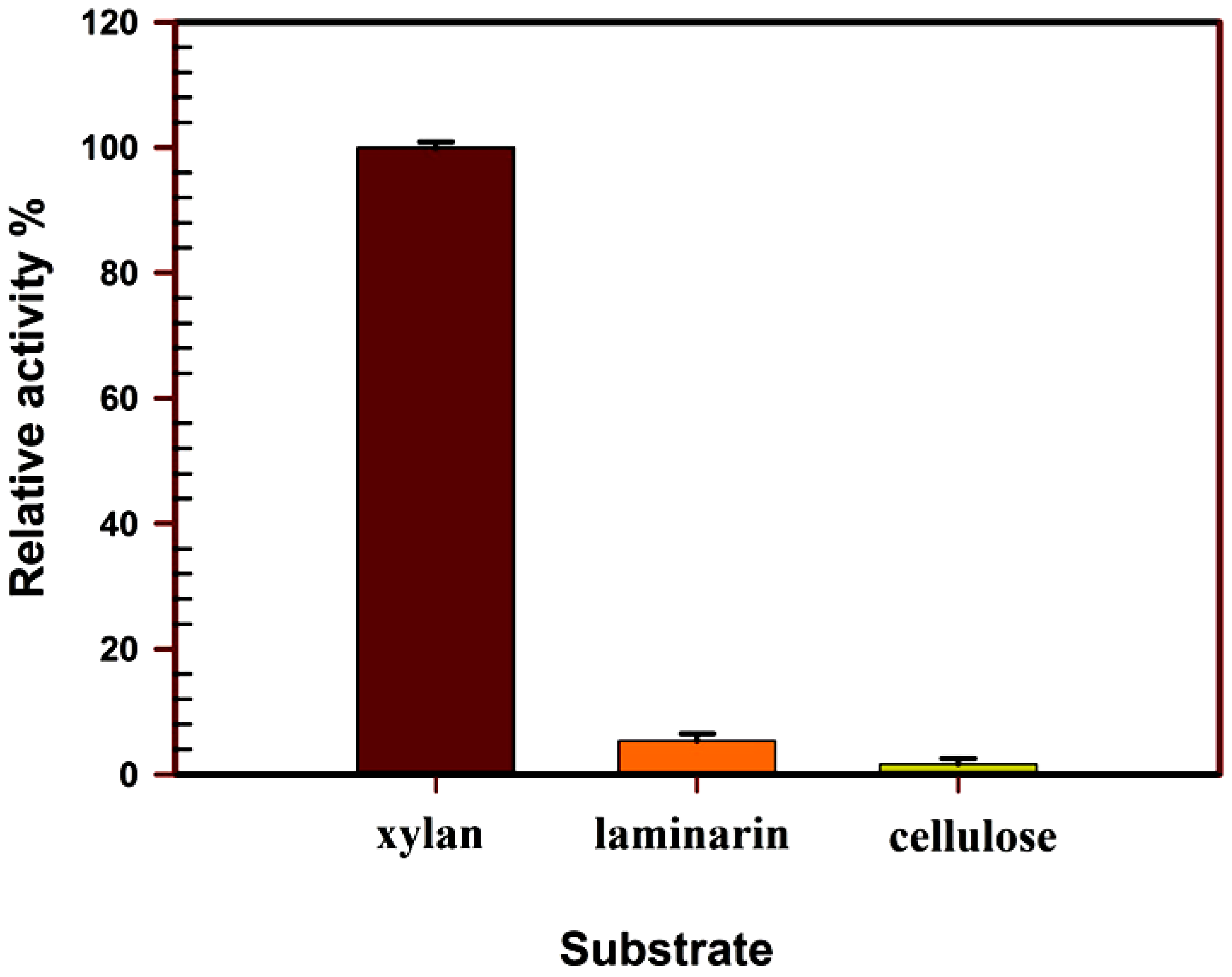
Substrate Specificity
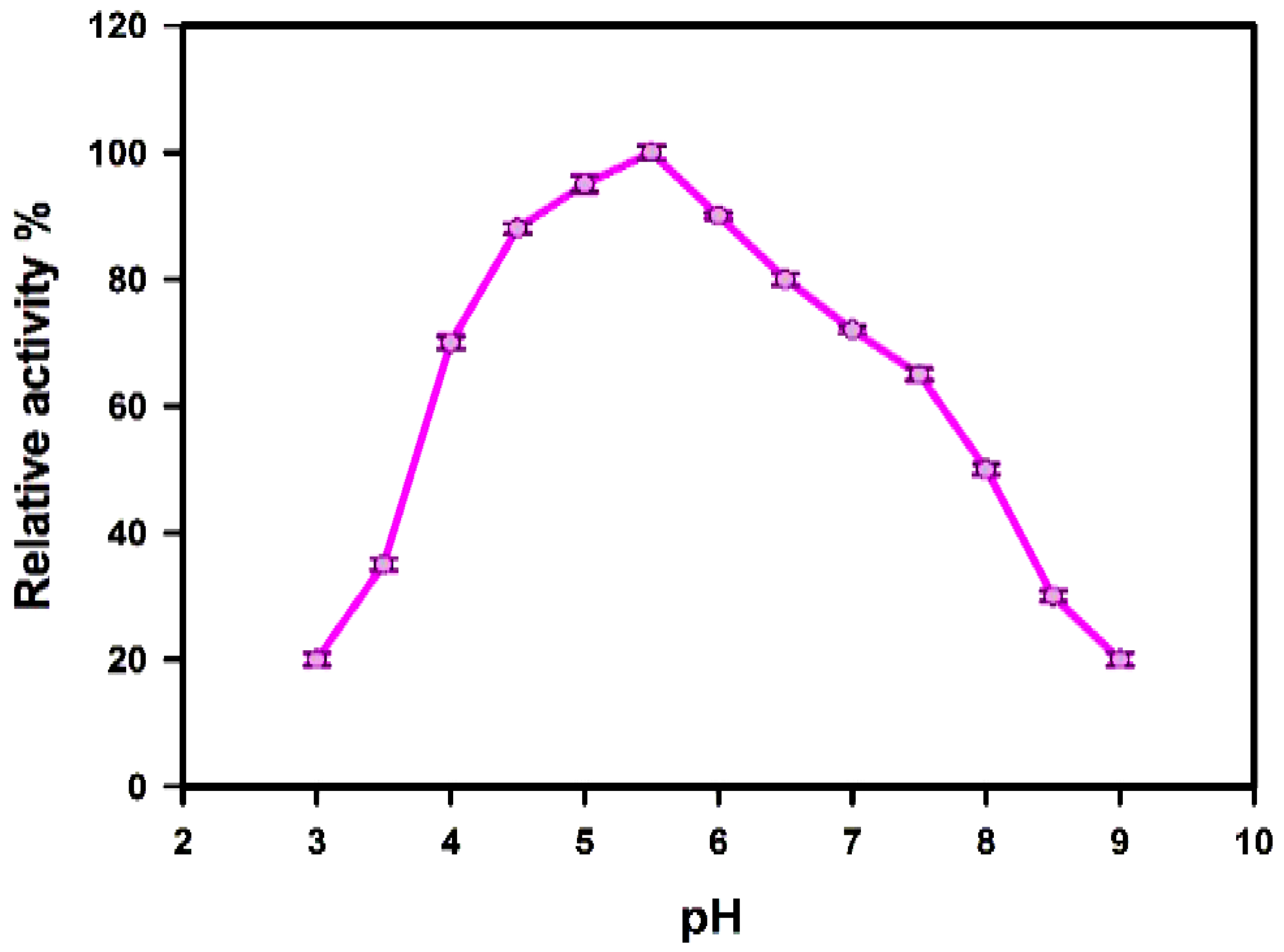
Effect of pH
Effect of Temperature
2.8. Bioreactor Cultivation Conditions
2.9. DNA Manipulation
2.9.1. Isolation of Xylanase Gene from T. harzianum
2.9.2. Screening of the Transformed Cells and Confirmation of Clones
2.9.3. PCR Confirmation of Recombinant PUC19 Plasmid
2.9.4. Screening of Xylanase Production
2.9.5. Extraction of Xylanase Gene and Assay of Enzyme
2.9.6. Antifungal Activity Assay
2.10. Statistical Analysis
3. Results
3.1. Phylogenetic Relationships
3.2. Optimization of Xylanase Productivity
3.3. Purification of Xylanase Enzyme
3.4. Characterization of Purified Xylanase
3.4.1. Substrate Specificity
3.4.2. Temperature and pH Stability
3.4.3. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
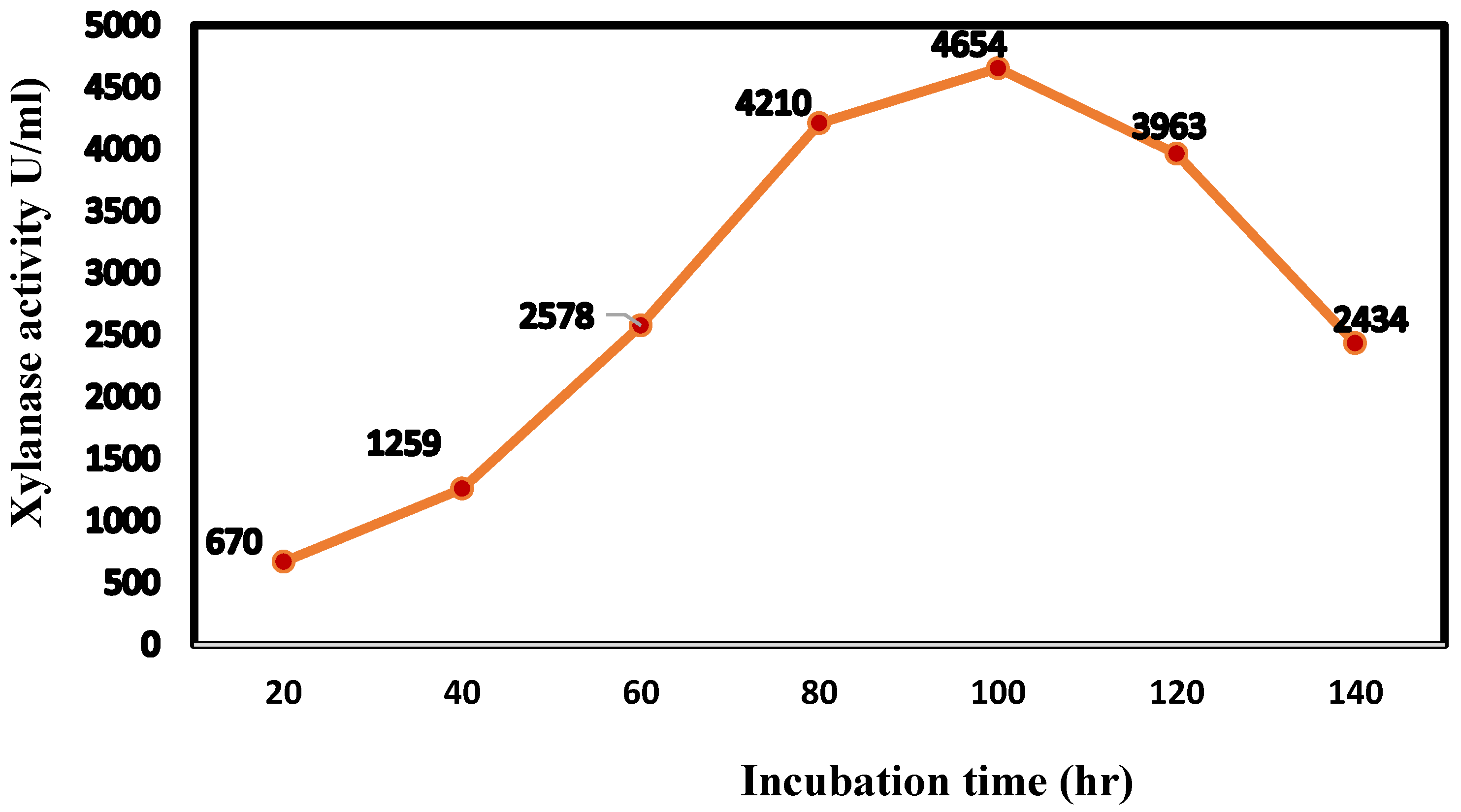
3.5. Cultivation Conditions for Large-Scale Production of Xylanase Enzyme in a Bioreactor
3.6. Isolation and Transformation of Xylanase Gene
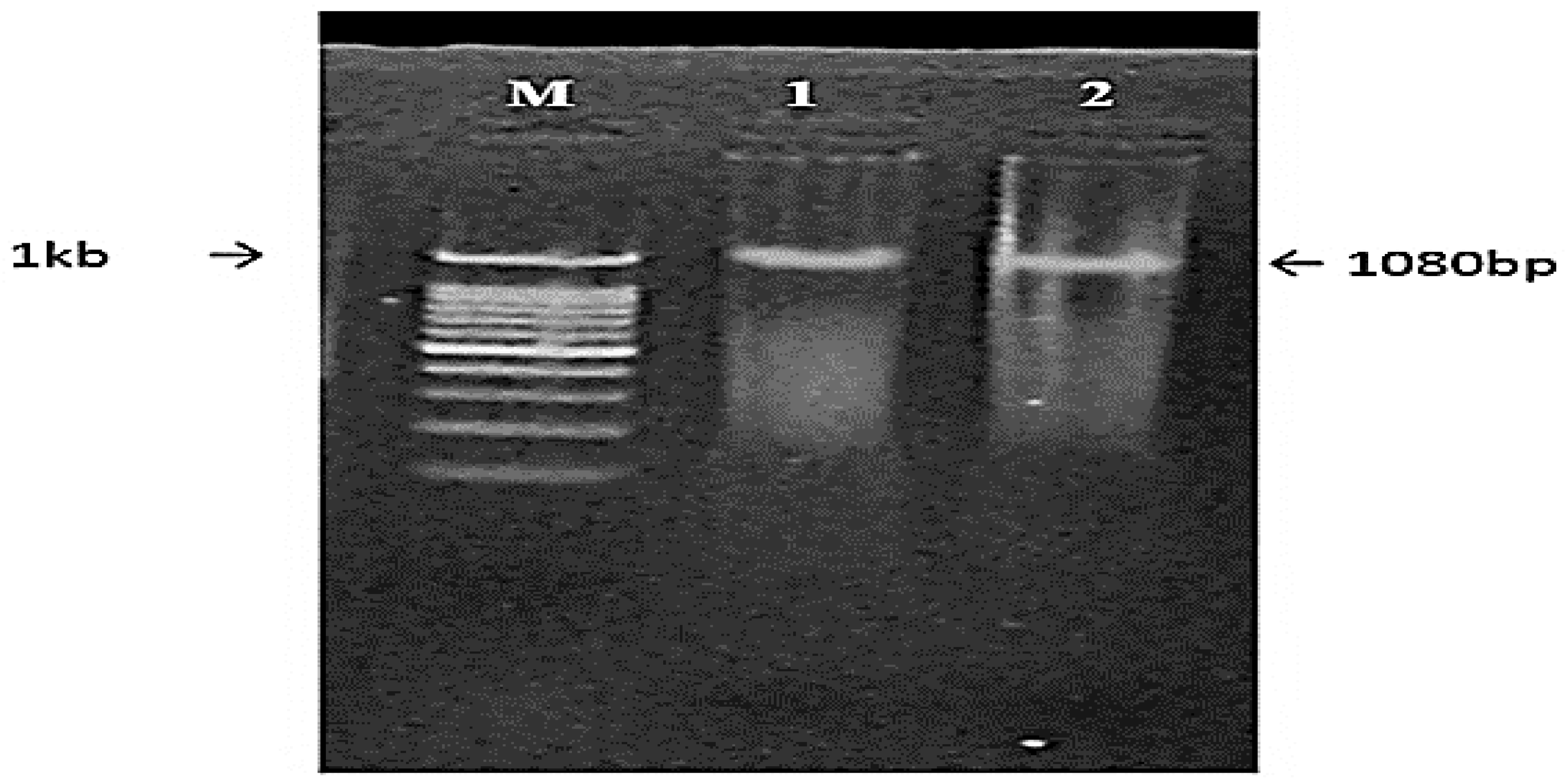
3.6.1. Screening of the Transformed Cells
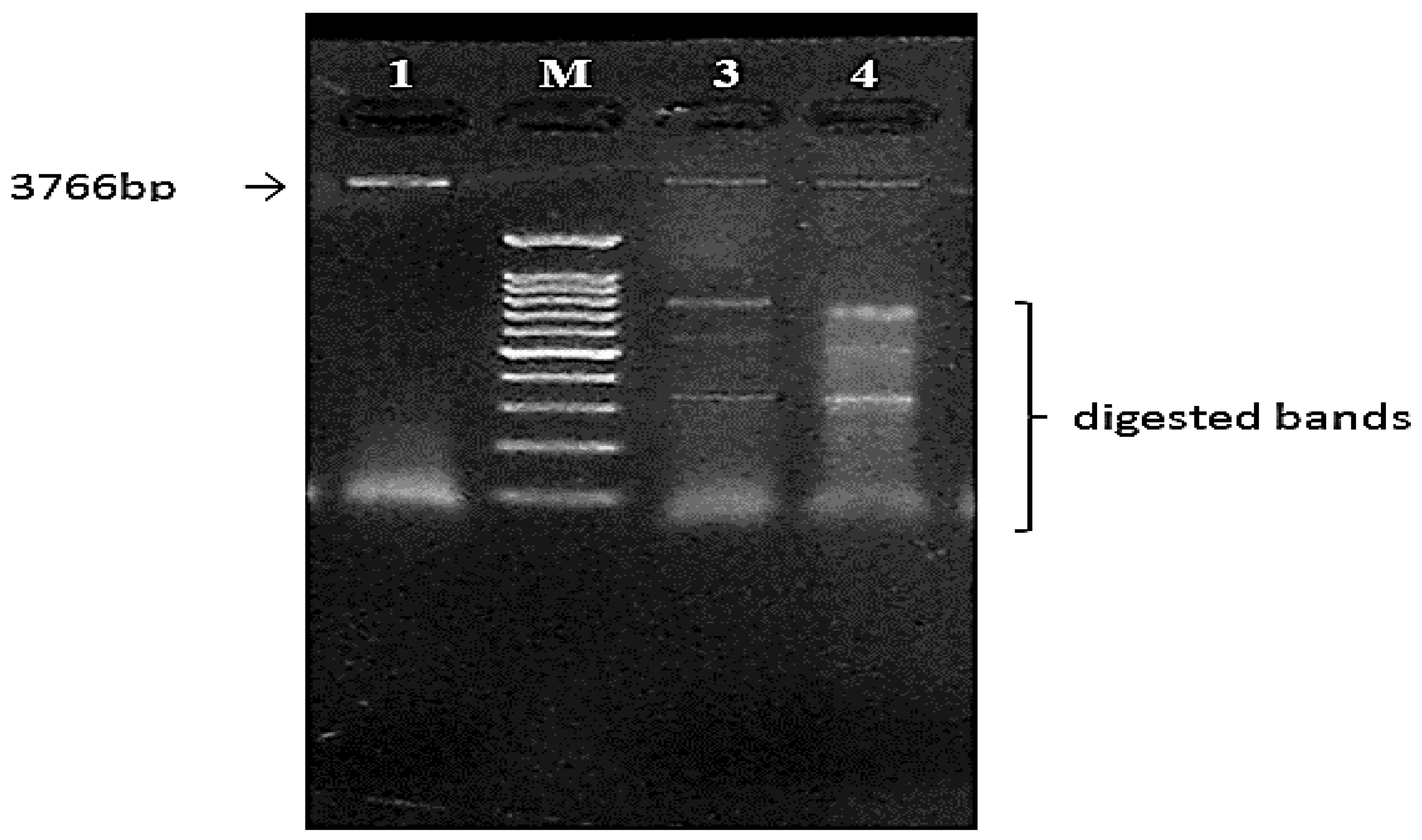
3.6.2. Confirmation Using Restriction Analysis
3.6.3. Estimation of Xylanase Activity
3.6.4. In Vitro Antifungal Activity of Crude Xylanase
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez-Pacheco, M.M.; Flores-García, A.; Zamudio-Jaramillo, M.A.; Chávez- Parga, M.C.; Alvarez-Navarrete, M. Optimization of production of xylanases with low cellulases in Fusarium solani by means of a solid state fermentation using statistical experimental design. Rev. Argent. De Microbiol. 2020, 52, 328–338. [Google Scholar] [CrossRef]
- Jae, W.L.; Jun, Y.P.; Mi, K.; In, G. Purification and characterization of a thermostable xylanase from the brown rot fungus Laetiporus sulphureus. J. Biosci. Bioeng. 2009, 107, 33–37. [Google Scholar]
- Meineke, T.; Manisseri, C.; Voigt, C.A. Phylogeny in defining model plants for lignocellulosic ethanol production: A comparative study Phylogeny in defining model plants for lignocellulosic ethanol production: A comparative study of Brachypodium distachyon, wheat, maize, and Miscanthus x giganteus leaf and stem biomass. PLoS ONE 2014, 9, e103580. [Google Scholar] [CrossRef]
- Sella, L.; Gazzetti, K.; Faoro, F.; Odorizzi, S.; D’Ovidio, R.; Schäfer, W.; Favaron, F. A Fusarium graminearum xylanase expressed during wheat infection is a necrotizing factor but is not essential for virulence. Plant Physiol. Biochem. 2013, 64, 1–10. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V.; Partridge, G.G. Beneficial effects of xylanase and/or phytase inclusions on ileal amino acid digestibility, energy utilization, mineral retention and growth performance in wheat-based broiler diets. Anim. Feed Sci. Technol. 2009, 153, 303–313. [Google Scholar] [CrossRef]
- Dhiman, S.S.; Sharma, J.; Battan, B. Industrial applications and future prospects of microbial xylanases: A Review. BioResources 2008, 3, 1377–1402. [Google Scholar]
- Maalej-Achouri, I.; Guerfali, M.; Gargouri, A.; Belghith, H. Production of xylooligosaccharides from agro-industrial residues using immobilized Talaromyces thermophilus xylanase. J. Mol. Catal. B Enzym. 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Selinheimo, E.; Kruus, K.; Buchert, J.; Hopia, A.; Autio, K. Kinetics of transglutaminase-induced cross-linking of wheat proteins in the dough. J. Cereal. Sci. 2006, 43, 152–159. [Google Scholar] [CrossRef]
- Nitin, K.S.; Vivek, K.T.; Santosh, K.M. The production of xylanase enzyme (E.C. Number = 3.2.1.8) using solid substrate fermentation. Biotech. Indian J. Biol. -Technol. 2017, 13, 145–153. [Google Scholar]
- Shabeena, K.S.; Ravi, M.; Jayaraj, Y.M. Microbial production of xylanase using regional agro-wastes. Int. J. Pharm. Biol. Sci. 2017, 8, 796–804. [Google Scholar] [CrossRef]
- Chadha, B.S.; Kaur, B.; Basotra, N.; Tsang, A.; Pandey, A. Thermostable xylanases from thermophilic fungi and bacteria: Current perspective. Bioresour. Technol. 2019, 277, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Yaashikaa, P.R.; Saravanan, A. Isolation, characterization, and purification of xylanase producing bacteria from sea sediment. Biocatal. Agric. Biotechnol. 2018, 13, 299–303. [Google Scholar] [CrossRef]
- Ergun, B.G.; Çalık, P. Lignocellulose degrading extremozymes produced by Pichia pastoris: Current status and future prospects. Bioprocess Biosyst. Eng. 2016, 39, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Yardimci, G.D.; Cekmecelioglu, D. Assessment and optimization of xylanase production using co-cultures of Bacillus subtilis and Kluyveromyces marxianus. 3 Biotech 2018, 8, 290. [Google Scholar] [CrossRef]
- van den Brink, J.; de Vries, R.P. Fungal enzyme sets for plant polysaccharide degradation. Appl. Microb. Biol. -Technol. 2011, 91, 1477–1492. [Google Scholar] [CrossRef] [Green Version]
- Long, C.; Liu, J.; Gan, L.; Zeng, B.; Long, M. Optimization of xylanase production by Trichoderma orientalis using corn cobs and wheat bran via statistical strategy. Waste Biomass Valorization 2019, 10, 1277–1284. [Google Scholar] [CrossRef]
- Atalla, M.M.S.; Ahmed, N.E.; Awad, H.M.; El Gamal, N.G.; El Shamy, A.R. Statistical optimization of xylanase production, using different agricultural wastes by Aspergillus oryzaeMN894021, as a biological control of faba bean root diseases. Egypt J. Biol. Pest Control. 2020, 30, 125. [Google Scholar] [CrossRef]
- Sanjay, K.; Mandal, A.P.; das Mohapatra, K.; Mondal, K.C.; Pati, B.R. Production of cellulase-free xylanase by Trichoderma reesei saf3. Braz. J. Microbiol. 2006, 37, 462–464. [Google Scholar]
- Silva, L.A.O.; Terrasan, C.R.F.; Carmona, E.C. Purification and characterization of xylanases from Trichoderma inhamatum. Electron. J. Biotechnol. 2015, 18, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Singh, J. Toxic molds and indoor air quality. Indoor Built Environ. 2005, 14, 229–234. [Google Scholar] [CrossRef]
- Wong, K.K.Y.; Tan, L.U.L.; Saddler, J.N. Multiplicity of b-1,4- xylanases in microorganisms: Functions and applications. Microbiol. Rev. 2005, 52, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.B.; Oh, D.K. Xylitol production by Candida tropicalis in a chemically defined medium. Biotechnol. Lett. 2003, 25, 2085–2088. [Google Scholar] [CrossRef]
- Mishra, R.K.; Monika, M.; Naimuddin, K.V. Trichoderma asperellum: A potential biocontrol agents against wilt of pigeonpea caused by Fusarium udum Butler. J. Food Legumes 2018, 31, 50–53. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Sikora RA, Schaefer K, Dababat AA Modes of action associated with microbially induced in planta suppression of plant-parasitic nematodes. Australas Plant Pathol. 2007, 36, 124–134. [CrossRef]
- Monte, E.; Bettiol, W.; Hermosa, R. Trichodermae Seus mecanismos de ação para o controle de doenças de plantas. In Trichoderma—Usona Agricultura; Meyer, M.C., Mazaro, S.M., Silva, J.C., Eds.; Embrapa: Brasília, Brazil, 2019; 538p. [Google Scholar]
- Ximenes, E.A.; Dien, B.S.; Ladisch, M.R.; Mosier, N.; Cotta, M.A.; Li, X.L. Enzyme production by industrially relevant fungi cultured on coproduct from corn dry-grind ethanol plants. Appl. Biochem. Biotechnol. 2007, 137, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A. Recent process developments in solid-state fermentation. Process Biochem. 1992, 27, 12–17. [Google Scholar] [CrossRef]
- Álvarez-Cervantes, J.; Domínguez-Hernández, E.M.; Mercado-Flores, Y.; O’Donovan, A.; Díaz-Godínez, G. Mycosphere Essay 10: Properties and characteristics of microbial xylanases. Mycosphere 2016, 7, 1600–1619. [Google Scholar] [CrossRef]
- Rosmine, E.; Sainjan, N.C.; Silvester, R. Statistical optimization of xylanase production by estuarine Streptomyces sp. and its application in clarification of fruit juice. J. Genet. Eng. Biotechnol. 2017, 15, 393–401. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Mathew, A.K. Development of a novel ultrasound-assisted alkali pretreatment strategy for the production of bioethanol and xylanases from chili post-harvest residue. Biol. -Resour. Technol. 2017, 242, 146–151. [Google Scholar] [CrossRef]
- Yong Syuan, K.; Ong Gaik Ai, L.; Kim Suan, T. Evaluation of cellulase and xylanase production from Trichoderma harzianum using acid-treated rice straw as solid substrate. Mater. Today Proc. 2018, 5, 22109–22117. [Google Scholar] [CrossRef]
- Karthick Raja Namasivayam, S.; Babu, M.; Arvind Bharani, R.S. Evaluation of lignocellulosic agro wastes for the enhanced production of extracellular cellulase and xylanase by Trichoderma harzianum. Nat. Environ. Pollut. Technol. 2015, 14, 47–52. [Google Scholar]
- Zafar, A.; Aftab, M.N.; Din, Z.U. Cloning, Expression, and Purification of Xylanase Gene from Bacillus licheniformis for Use in Saccharification of Plant Biomass. Appl. Biol. -Chembiol. -Technol. 2016, 178, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Riaz, S.; Jamil, A. Molecular cloning of fungal xylanases: An overview. Appl. Microbiol. Biol. -Technol. 2009, 84, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.H.; Lin, Z.P.; Tu, J. Xylanase production by Paenibacillus campinasensis BL11 and its pretreatment of hardwood kraft pulp bleaching. Int. Biodeterior. Biodegrad. 2010, 64, 13–19. [Google Scholar] [CrossRef]
- Lee, C.K.; Jang, M.Y.; Park, H.R.; Choo, G.C.; Cho, H.S.; Park, S.B.; Oh, K.C.; An, J.B.; Ki, B.G. Cloning and characterization of xylanase in cellulolytic Bacillus sp. strain JMY1 isolated from forest soil. Appl. Biol. Chem. 2016, 59, 415–423. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification, and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phyto-Chem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Nayaka, C.S.; Niranjan Raj, S.; Venkataramana, M.; Gupta, V.K.; Gurunathan, S.; Shobith, R.; Shetty, H.S.; Srivastava, R.K. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, 43991. [Google Scholar]
- Paul, S.P.; Shin, Y.C.; Park, C.S.; Chung, Y.R. Biological control of fusarium wilt of cucumber by chitinolytic bacteria. Phytopathology 1999, 89, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Mandels, M.; Andreotti, R.; Roche, R. Measurement of saccharifying cellulase. Biotechnol. Bioeng. Symp. 1976, 6, 17–37. [Google Scholar]
- Haltrich, D.; Nidetzky, B.; Kulbe, K.D.; Steiner, W.; Zupaneie, S. Production of fungal xylanases. Bioresour. Technol. 1996, 58, 137–161. [Google Scholar] [CrossRef]
- Tangnu, S.K.; Blanch, H.W.; Wilke, C.R. Enhanced production of cellulase, hemicellulase, and β-glucosidase by Trichoderma reesei Rut C-30. Biotechnol. Bioeng. 1981, 23, 1837–1849. [Google Scholar] [CrossRef]
- Panda, T.; Gruber, H.; Kubicek, C.P. Stimulation of protein secretion in Trichoderma reesei by Tween surfactants is not correlated with changes in enzyme localization or membrane fatty acid composition. FEMS Microbiol. Lett. 1987, 41, 85–90. [Google Scholar] [CrossRef]
- Bagewadi, Z.K.; Mulla, S.I.; Ninnekar, H.Z. Purification and characterization of endo b-1,4-D-glucanase from Trichoderma harzianum strain HZN11 and its application in the production of bioethanol from sweet sorghum bagasse. 3 Biotech 2016, 6, 101. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Würleitner, E.; Pera, L.; Wacenovsky, C.; Mach, R. Transcriptional Regulation of xyn2 in Hypocreajecorina. Eukaryot. Cell 2003, 2, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, C.; Annie, N.; Chang, C.Y.; Hsu, L. Non-chromosomal Antibiotic Resistance in Bacteria: Genetic Transformation of Escherichia coli by R-factor DNA. Proc. Natl. Acad. Sci. USA 1972, 69, 2110–2114. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Sanger, F., Nicklen, S., Coulson, A.R., Eds.; Cold Spring Harbor Laboratory; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor (NY), Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; pp. 132–150. [Google Scholar]
- Birnboim, H.C.; Doly, J.A. Rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Roberts, W.K.; Selitrennikoff, C.P. Isolation and partial characterization of two antifungal proteins from barley. Biochim. Biol. -Phys. Acta 1986, 880, 161–170. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetic analysis across computing platform. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Seeta Laxman, R.; Sonawane, A.P.; More, S.V.; Seetarama Rao, B.; Rele, M.V.; Jogdand, V.V.; Deshpande, V.V.; Rao, M.B. Optimization and scale-up of production of alkaline protease from Conidiobolus coronatus. Process Biochem. 2005, 40, 3152–3158. [Google Scholar] [CrossRef]
- de Queiroz-Fernandes, M.; Martins, B.L.; Geisiany, C.B.R. Reuse of wastewater from pulp industry for the optimization of fungal xylanase production. Acta Sci. Biol. Sci. 2017, 39, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Bhushan, B.; Hoondal, G.S. Isolation, purification, and characterization of xylanase from Staphylococcus sp. SG-13 and its application in bio-bleaching of kraft pulp. J Appl. Microbiol. 2000, 88, 325–334. [Google Scholar] [CrossRef]
- Al-Sa’ady Ali, J.R. Determination of optimum conditions for xylanase production by Aspergillus niger using solid-state fermentation. Iraqi J. Sci. 2016, 57, 2220–2227. [Google Scholar]
- Parichart, K.; laothanachareon, T.; Rattanaphan n Tinnasulanon, P.; Apawasin, S.; Paemanee, A.; Ruanglek, V.; Tanapongpipat, S.; Champreda, V.; Eurwilaichitr, L. Optimization of Xylanase Production from aspergillus niger for Bio-bleaching of EucalyptusPulp. Biosci. Biotechnol. Biochem. 2011, 75, 1129–1134. [Google Scholar] [CrossRef]
- Sonika, P.; Srivastava, M.; Sharma, A.; Singh, A.; Kumar, V.; Srivastava, Y. Isolation and Optimized Production of Xylanase under Solid-State Fermentation Condition from Trichoderma sp. Int. J. Adv. Res. 2014, 2, 263–273. [Google Scholar]
- Shah, A.R.; Madamwar, D. Xylanase production by newly isolated Aspergillus foresides strain and its characterization. Process Biochem. 2005, 40, 1763–1771. [Google Scholar] [CrossRef]
- Sridevi, B.; Charya, M.A.S. Isolation, identification, and screening of potential cellulase-free xylanase-producing fungi. Afr. J. Biotechnol. 2011, 10, 4624–4630. [Google Scholar]
- Pang, P.K.; Darah, I.; Poppe, L.; Szakacs, G.; Ibrahim, C.O. Xylanase production by a local isolate, Trichoderma spp. FETL c3-2 via solid-state fermentation using agricultural wastes as substrates. Malays. J. Microbiol. 2006, 2, 7–14. [Google Scholar]
- Ghoshal, G.; Kamble, A.; Shivhare, U.S.; Banerjee, U.C. Optimization of culture conditions for the production of xylanase in submerges fermentation by Penicillium citrinum using response surface methodology. IRAS 2011, 6, 132–137. [Google Scholar]
- Richhariya, J.; Sharma, T.K.; Dassani, S. Production and optimization of enzyme xylanase by Aspergillus flavus using agricultural waste residues. J. Appl. Biol. Biotech. 2020, 8, 82–89. [Google Scholar]
- Azzouz, Z.; Bettache, A.; Boucherba, N.; Prieto, A.; Martinez, M.J.; Benallaoua, S.; de Eugenio, L.I. Optimization of β-1,4-Endoxylanase Production by an Aspergillus niger Strain Growing on Wheat Straw and Application in Xylooligosaccharides Production. Molecules 2021, 26, 2527. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Ljungdahl, L.G. Cloning, sequencing and regulation of a xylanase gene from the fungus Aureobasidium pullulans Y-2311-1. Appl. Environ. Microbiol. 1994, 60, 3160–3166. [Google Scholar] [CrossRef] [Green Version]
- Nagar, S.; Gupta, V.K.; Kumar, D.; Kumar, L.; Kuhad, R.C. Production and optimization of cellulase-free, alkali-stable xylanase by Bacillus pumilus SV-85S in submerged fermentation. J. Ind. Microbiol. Biotechnol. 2010, 37, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Arnesen, S.; Eriksen, S.H.; Olsen, J.O.; Jensen, B. Increased production of α- amylase from Thermomyces lanuginosus by addition of Tween 80. Enzyme Microbe. Technol. 1998, 23, 249–252. [Google Scholar] [CrossRef]
- Eriksson, T.; Börjesson, J.; Tjerneld, F. Mechanism of surfactant effect in enzymatic hydrolysis of lignocelluloses. Enzyme Microb. Technol. 2002, 31, 12. [Google Scholar] [CrossRef]
- Kruszewska, J.; Palamarczyk, G.; Kubicek, C.P. Stimulation of exoprotein secretion by choline and Tween 80 in Trichoderma reesei QM 9414 correlates with increased activities of dolichol phosphate mannose synthase. J. Gen. Microbiol. 1990, 136, 1293–1298. [Google Scholar] [CrossRef] [Green Version]
- Kheng, P.P.; Omar, I.C. Xylanase production by a local fungal isolate, Aspergillus niger USM AI 1 via solid-state fermentation using palm kernel cake (PKC) as substrate Songklanakarin. J. Sci. Technol. 2005, 27, 325–336. [Google Scholar]
- Okafor, U.A.; Okochi, V.I.; Onyegeme-okerenta, B.M.; Nwodo Chinedu, S. Xylanase production by Aspergillus niger ANL 301 using agro-wastes. Afr. J. Biotechnol. 2007, 6, 1710–1714. [Google Scholar]
- Bakri, Y.; Jawhar, M.; Arabi, M.I.E. Improvement of xylanase production by Cochliobolus sativus in submerged culture. Food Technol. Biotechnol. 2008, 46, 116–118. [Google Scholar]
- Pal, A.; Ramana, K.V. Purification and characterization of bacteriocin from Weissella paramesenteroides DFR-8, an isolate from cucumber (Cucumis sativus). J. Food Biochem. 2010, 34, 932–948. [Google Scholar] [CrossRef]
- da Silva, R.; Lago, E.S.; Merheb, C.W.; Merheb, M.M.; Macchione, Y.; Park, K.; Gomes, E. Production of xylanase and CMCase on solid-state fermentation in different residues by Thermoascus aurantiacus Miehe. Braz. J. Microbiol. 2005, 36, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Muthezhilan, R.; Ashok, R.; Jayalakshmi, S. Production and optimization of thermostable alkaline xylanase by Penicillium oxalicum in solid-state fermentation. Afr. J. Microbiol. Res. 2007, 20–28. [Google Scholar]
- Terrasan, C.R.; Temer, B.; Duarte, M.C.; Carmona, E.C. Production of xylanolytic enzymes by Penicillium janczewskii. Bioresour Technol. 2010, 101, 4139–4143. [Google Scholar] [CrossRef]
- Howard, R.L.; Abotsi, E.; Jansen van Rensburg, E.L.; Howard, S. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2003, 2, 602–619. [Google Scholar] [CrossRef]
- Jhamb, K.; Sahoo, D.K. Production of soluble recombinant proteins in Escherichia coli: Effects of process conditions and chaperone co-expression on cell growth and production of xylanase. Bioresour. Technol. 2012, 123, 135–143. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production, and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Motta, F.L.; Andrade, C.C.P.; Santana, M.H.A. A review of xylanase production by the fermentation of xylan: Classification, characterization and applications. In Sustainable Degradation of Lignocellulosic Biomass Techniques, Applications, and Commercialization; InTechOpen: New York, NY, USA, 2013; pp. 251–275. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Thakur, P.; Rathore, U.S.; Kumar, S.; Mishra, R.K.; Amaresan, N.; Pandey, S.; Mishra, M. Plant beneficial effects of Trichoderma spp. Suppressing Fusarium wilt and enhancing growth in Tomato. Vegetos 2022, 35, 188–195. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Ros, M.; Pascual, J. Added-value of Trichoderma amended compost as biopesticide organic substrates: Alternative to traditional organic substrates. Acta Horticulturae 2011, 898, 189–196. [Google Scholar] [CrossRef]
- Mishra, R.K.; Bohra, A.; Kamaal, N.; Kumar, K.; Sujayanand, G.K.; Saabale, P.R.; Naik, S.J.; Sarma, B.K.; Kumar, D.; Mishra, M.; et al. Utilization of biopesticides as sustainable solutions for management of pests in legume crops: Achievements and prospects. Egypt. J. Biol. Pest Control 2018, 28, 3. [Google Scholar] [CrossRef]
- Gomes, E.V.; Ulhoa, C.J.; Cardoza, R.E.; Silva, R.N.; Gutiérrez, S. Involvement of Trichoderma harzianum Epl-1 protein in the regulation of botrytis virulenceand tomato defense-related genes. Front. Plant Sci. 2017, 8, 880. [Google Scholar] [CrossRef] [PubMed]
- Jayalakshmi, S.K.; Raju, S.; Usha Rani, S.; Benagi, V.I.; Sreeramulu, K. Trichoderma harzianum L1 as a potential source for lytic enzymes and elicitor of defense responses in chickpea (Cicer arietinum L.) against wilt disease caused by Fusarium oxysporum f. sp. ciceri. Aust. J. Crop Sci. 2009, 3, 44–52. [Google Scholar]
- Ujiie, M.; Roy, C.; Yaguchi, M. Low-Molecular-Weight Xylanase from Trichoderma viride. Appl. Environ. Microbiol. 1991, 57, 1860–1862. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.U.L.; Wong, E.K.C.; Saddler, J.N. Purification and characterization of two D-xylanases from Trichoderma harzianum. Enzyme Microb. Technol. 1985, 7, 425–430. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tween 80 Conc. (mM) | Xylanase Activity (U/mL) |
|---|---|
| 0.1 mM + xylose | 1139.9 ± 0.02 |
| 0.1 mM + lactose | 2192.7 ± 0.9 |
| 0.2 mM + xylose | 347.68 ± 0.5 |
| 0.2 mM + lactose | 102.58 ± 0.44 |
| Inducer | Xylanase Activity (U/mL) |
|---|---|
| Galactose | 850 ± 0.3 |
| Sophorose | 712 ± 0.9 |
| Mannose | 590 ± 0.9 |
| Galactose + malt extract | 2636 ± 0.7 |
| Sophorose + malt extract | 2150 ± 1.1 |
| Mannose + malt extract | 1430 ± 1.4 |
| Agricultural Waste Concentration (%) | Xylanase Activity (U/mL) | ||||
|---|---|---|---|---|---|
| Different Agricultural Wastes | |||||
| Rice Bran | Rice Straw | Sugarcane Bagasse | Wheat Bran | Wood | |
| 0.25 | 1220.5 ± 0.72 | 843.9 ± 0.35 | 1250 ± 0.56 | 200 ± 0.1 | 2910 ± 0.40 |
| 0.5 | 3100.1 ± 0.48 | 2550 ± 0.12 | 2800.2 ± 0.82 | 319 ± 0.46 | 2800 ± 0.85 |
| 1 | 850.2 ± 0.30 | 2170.3 ± 0.40 | 3620.2 ± 1.25 | 510 ± 0.32 | 1420.1 ± 0.66 |
| Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purity (Fold) |
|---|---|---|---|---|
| Crude xylanase | 105 | 580 | 18.10 | 1 |
| Ammonium sulfate precipitation (80%) | 58 | 107 | 54.2 | 2.99 |
| Sephadex G-100 | 39 | 9 | 433 | 23.9 |
| Fungal Strain | Inhibition % |
|---|---|
| Alternariaalternate STE-U4349 | 63 |
| Botrytis fabae MU BF1 | 58 |
| Fusarium oxysporum KJ831189 | 60 |
| Fusarium solani KJ831188 | 54 |
| Fusarium avenaceum P21 70 | 20 |
| Corynespora cassiicola YC46 | 81 |
| Fusarium decemcellulare C82NL | 56 |
| Bipolaris oryzae 232 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellatif, S.A.; Abdel Razik, E.S.; AL-surhanee, A.A.; Al-Sarraj, F.; Daigham, G.E.; Mahfouz, A.Y. Enhanced Production, Cloning, and Expression of a Xylanase Gene from Endophytic Fungal Strain Trichoderma harzianum kj831197.1: Unveiling the In Vitro Anti-Fungal Activity against Phytopathogenic Fungi. J. Fungi 2022, 8, 447. https://doi.org/10.3390/jof8050447
Ellatif SA, Abdel Razik ES, AL-surhanee AA, Al-Sarraj F, Daigham GE, Mahfouz AY. Enhanced Production, Cloning, and Expression of a Xylanase Gene from Endophytic Fungal Strain Trichoderma harzianum kj831197.1: Unveiling the In Vitro Anti-Fungal Activity against Phytopathogenic Fungi. Journal of Fungi. 2022; 8(5):447. https://doi.org/10.3390/jof8050447
Chicago/Turabian StyleEllatif, Sawsan Abd, Elsayed S. Abdel Razik, Ameena A. AL-surhanee, Faisal Al-Sarraj, Ghadir E. Daigham, and Amira Y. Mahfouz. 2022. "Enhanced Production, Cloning, and Expression of a Xylanase Gene from Endophytic Fungal Strain Trichoderma harzianum kj831197.1: Unveiling the In Vitro Anti-Fungal Activity against Phytopathogenic Fungi" Journal of Fungi 8, no. 5: 447. https://doi.org/10.3390/jof8050447