
Arbuscular Mycorrhizal Fungi Induced Plant Resistance against Fusarium Wilt in Jasmonate Biosynthesis Defective Mutant and Wild Type of Tomato



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Fungi and Growth Substrate
2.2. Experimental Design and Procedure
2.3. Net Photosynthetic and Transpiration Rate
2.4. Disease Assessment
2.5. Plant Harvest
2.6. Mycorrhizal Colonization
2.7. Plant P Concentrations
2.8. RNA Extraction and Gene Expression Analysis
2.9. PPO and PAL Activities
2.10. Statistical Analysis
3. Results
3.1. Mycorrhizal Colonization
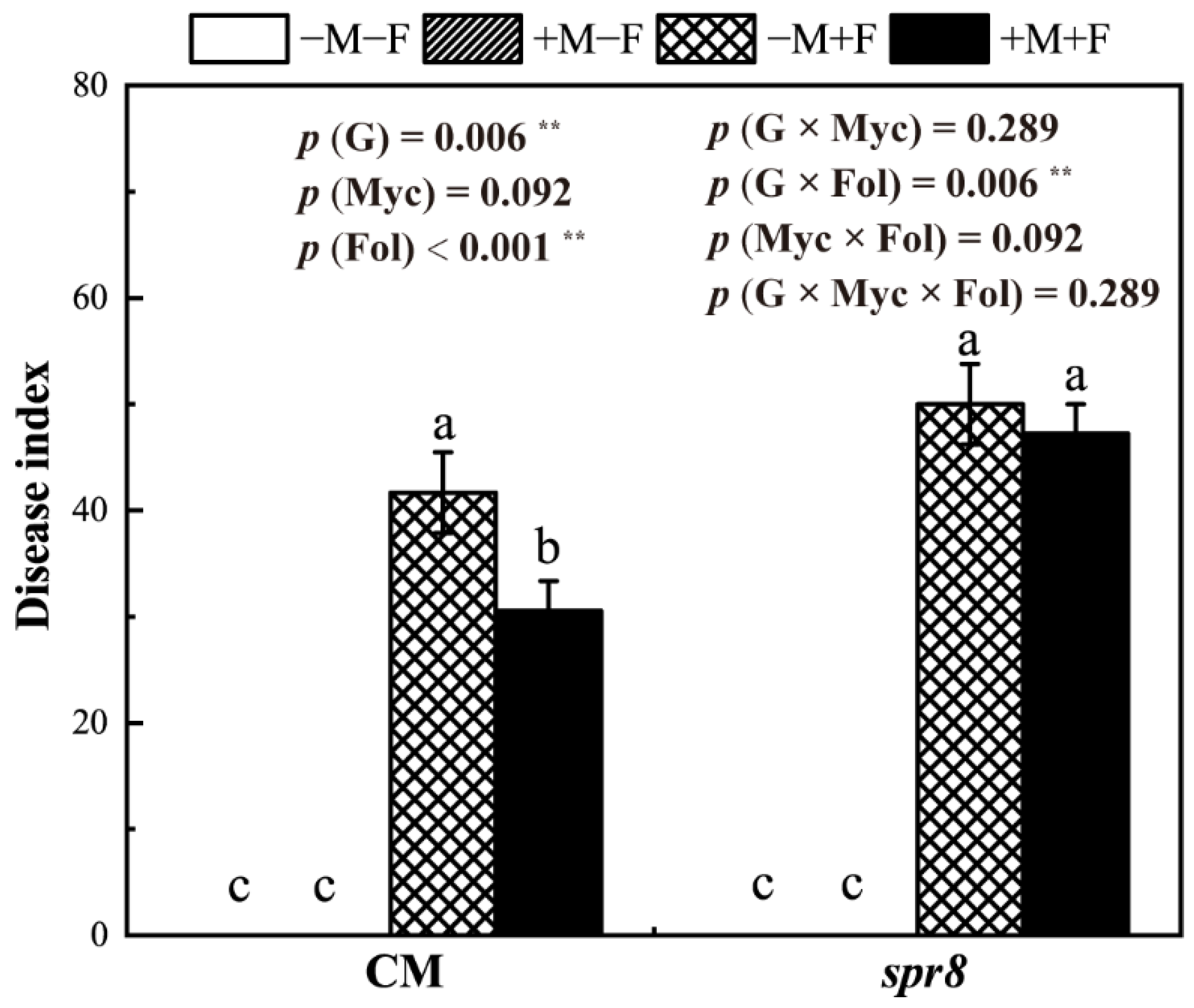
3.2. Disease Index
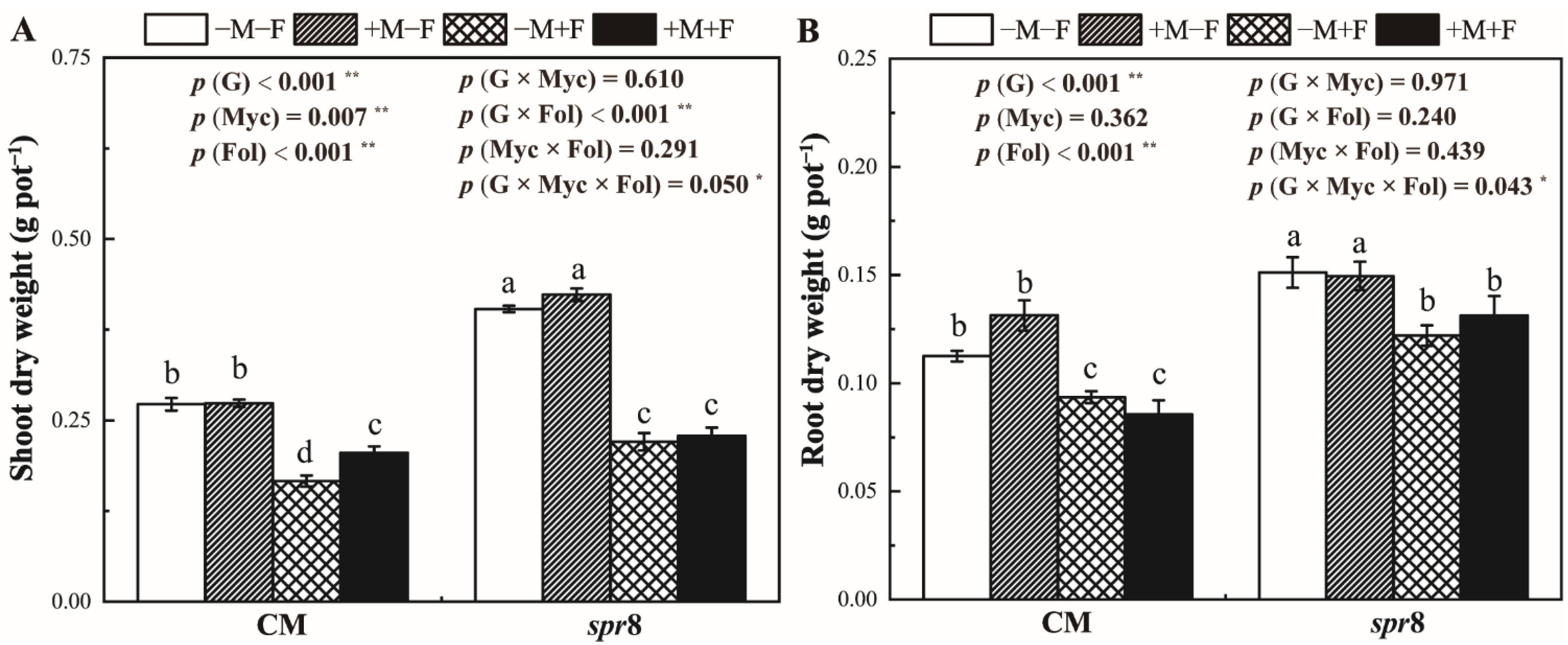
3.3. Plant Growth
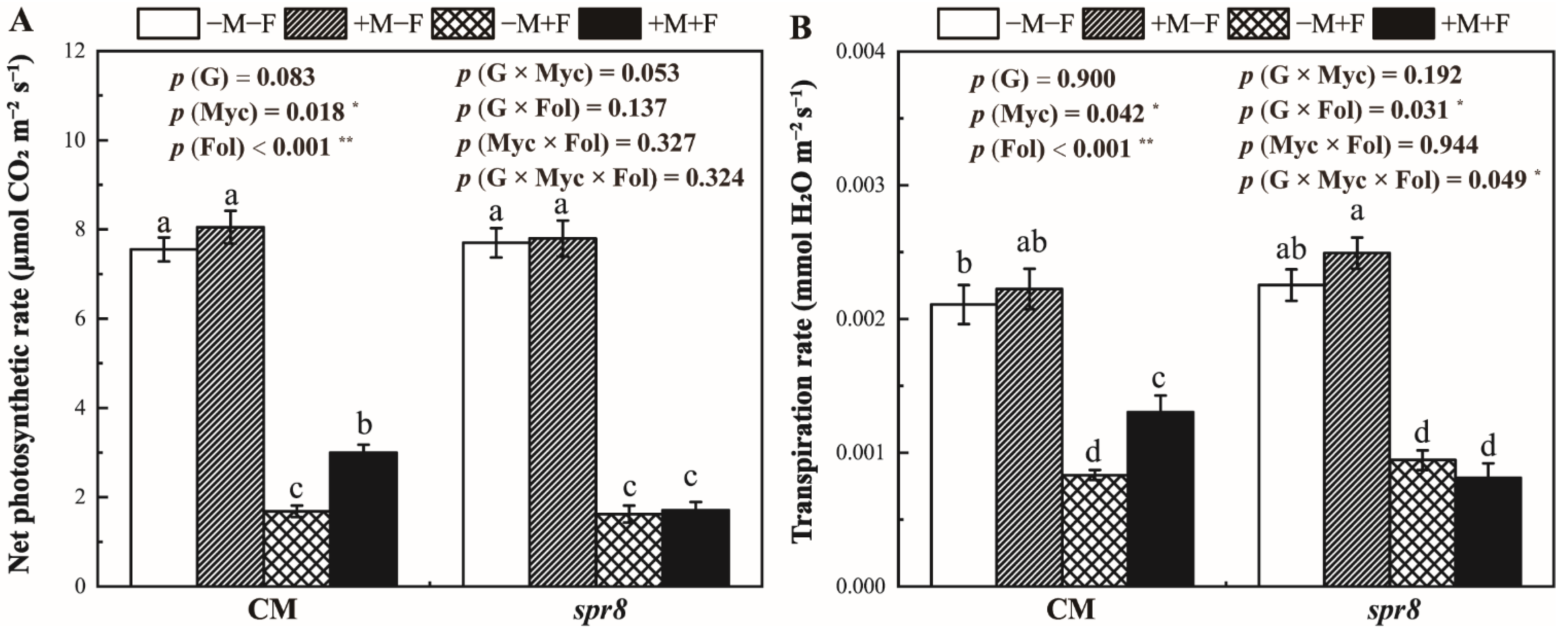
3.4. Net Photosynthetic and Transpiration Rate
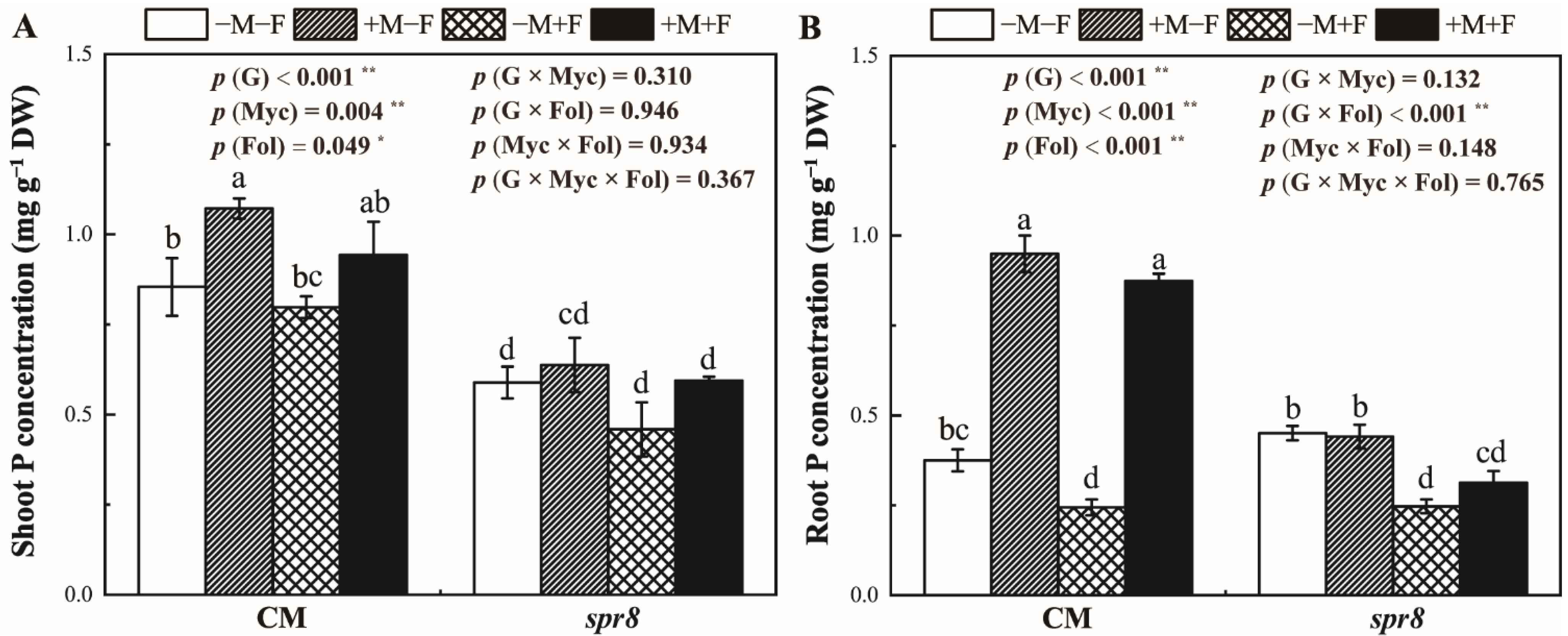
3.5. Shoot and Root P Concentrations
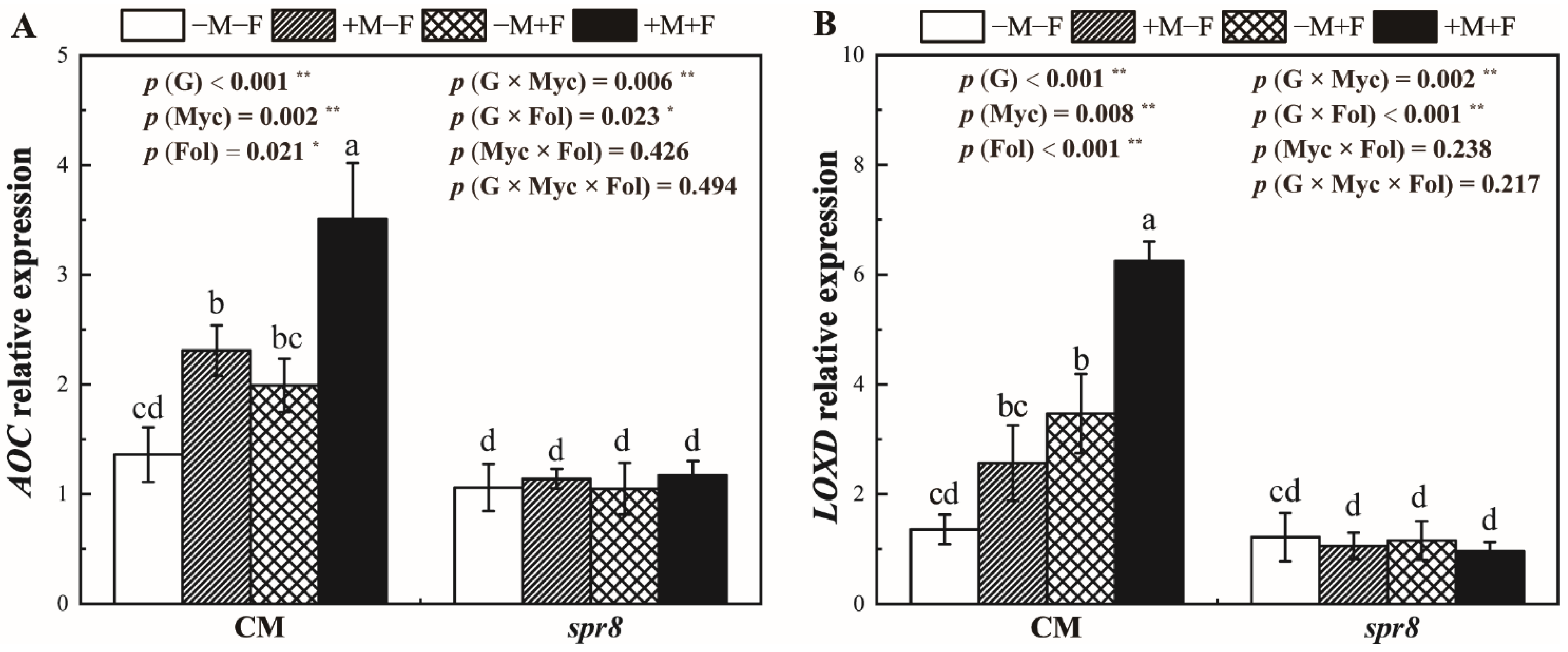
3.6. AOC and LOXD Relative Expression
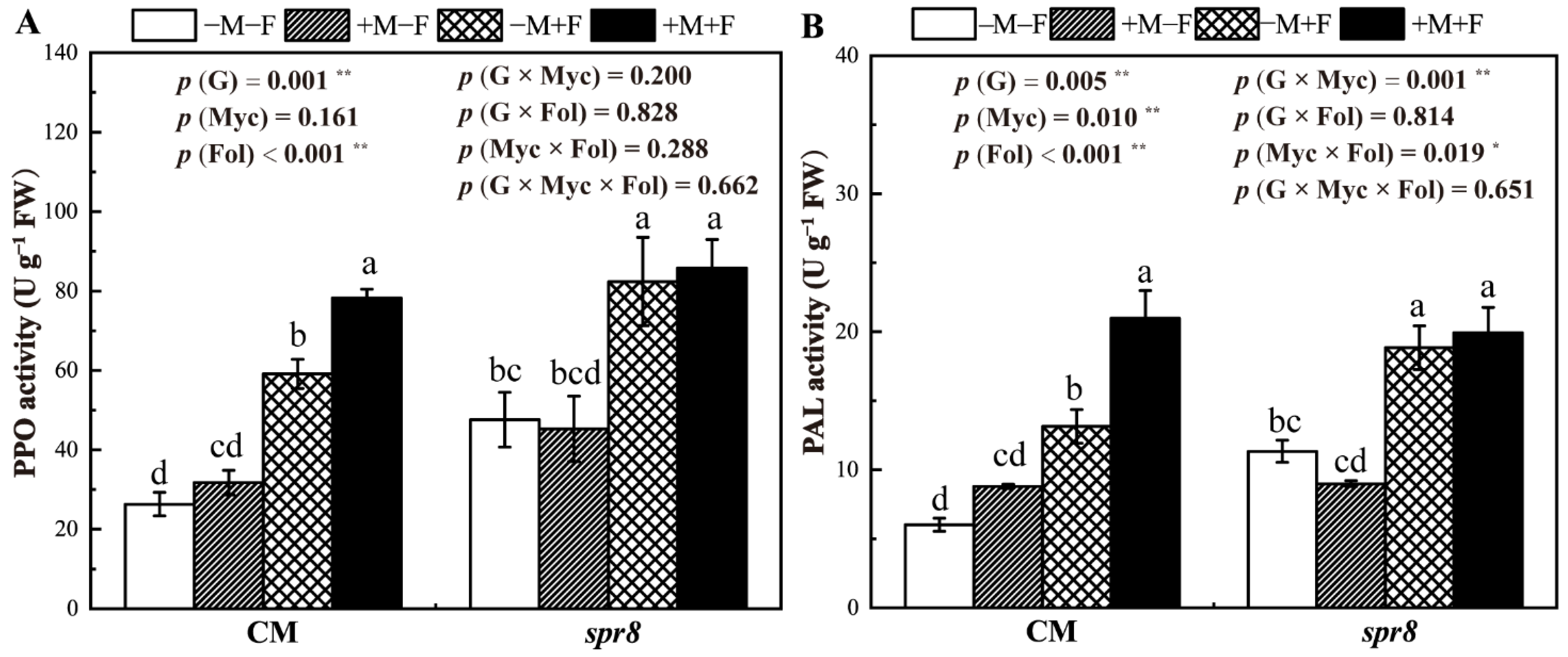
3.7. PPO and PAL Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vos, C.M.; Yang, Y.; De Coninck, B.; Cammue, B.P.A. Fungal (-like) biocontrol organisms in tomato disease control. Biol. Control 2014, 74, 65–81. [Google Scholar] [CrossRef]
- Panthee, D.R.; Chen, F. Genomics of fungal disease resistance in tomato. Curr. Genom. 2010, 11, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousa, M.A.A.; Abo-Elyousr, K.A.M.; Abdel Alal, A.M.K.; Alshareef, N.O. Management fusarium wilt disease in tomato by combinations of Bacillus amyloliquefaciens and peppermint Oil. Agronomy 2021, 11, 2536. [Google Scholar] [CrossRef]
- Chialva, M.; Salvioli di Fossalunga, A.; Daghino, S.; Ghignone, S.; Bagnaresi, P.; Chiapello, M.; Novero, M.; Spadaro, D.; Perotto, S.; Bonfante, P. Native soils with their microbiotas elicit a state of alert in tomato plants. New Phytol. 2018, 220, 1296–1308. [Google Scholar] [CrossRef] [Green Version]
- Redkar, A.; Ibanez, S.G.; Sabale, M.; Zechmann, B.; Solano, R.; Di Pietro, A. Marchantia polymorpha model reveals conserved infection mechanisms in the vascular wilt fungal pathogen Fusarium oxysporum. New Phytol. 2022, 234, 227–241. [Google Scholar] [CrossRef]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, A.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity-A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar]
- Jayamohan, N.S.; Patil, S.V.; Kumudini, B.S. Seed priming with Pseudomonas putida isolated from rhizosphere triggers innate resistance against Fusarium wilt in tomato through pathogenesis-related protein activation and phenylpropanoid pathway. Pedosphere 2020, 30, 651–660. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef]
- Hanson, P.; Lu, S.F.; Wang, J.F.; Chen, W.; Kenyon, L.; Tan, C.W.; Tee, K.L.; Wang, Y.Y.; Hsu, Y.C.; Schafleitner, R.; et al. Conventional and molecular marker-assisted selection and pyramiding of genes for multiple disease resistance in tomato. Sci. Hortic. 2016, 201, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Tanwar, A.; Aggarwal, A.; Panwar, V. Arbuscular mycorrhizal fungi and Trichoderma viride mediated Fusarium wilt control in tomato. Biocontrol Sci. Technol. 2013, 23, 485–498. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Abdel-Motaal, F.; El-Sayed, M.; Jogaiah, S.; Shigyo, M.; Ito, S.; Tran, L.S.P. Dissection of Trichoderma longibrachiatum-induced defense in onion (Allium cepa L.) against Fusarium oxysporum f. sp. cepa by target metabolite profiling. Plant Sci. 2016, 246, 128–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Abo-Elyousr, K.A.M.; Mousa, M.A.A.; Saad, M.M. A study on the synergetic effect of Bacillus amyloliquefaciens and dipotassium phosphate on Alternaria solani causing early blight disease of tomato. Eur. J. Plant Pathol. 2022, 162, 63–77. [Google Scholar] [CrossRef]
- Wang, X.; Ding, T.; Li, Y.; Guo, Y.; Duan, T. Dual inoculation of alfalfa (Medicago sativa L.) with Funnelliformis mosseae and Sinorhizobium medicae can reduce Fusarium wilt. J. Appl. Microbiol. 2020, 129, 665–679. [Google Scholar] [CrossRef]
- Walder, F.; van der Heijden, M.G.A. Regulation of resource exchange in the arbuscular mycorrhizal symbiosis. Nat Plants 2015, 1, 15159. [Google Scholar] [CrossRef] [PubMed]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2010. [Google Scholar]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal Rewards Stabilize Cooperation in the Mycorrhizal Symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef] [Green Version]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Lin, J.X.; Wang, Y.N.; Sun, S.N.; Mu, C.S.; Yan, X.F. Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition. Sci. Total. Environ. 2017, 576, 234–241. [Google Scholar] [CrossRef]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.W.; Yue, F.X.; Yan, X.W.; Wang, F.Y.; Bloszies, S.; Wang, Y.F. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef]
- Hao, Z.P.; Fayolle, L.; van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef]
- Chialva, M.; Zhou, Y.; Spadaro, D.; Bonfante, P. Not only priming: Soil microbiota may protect tomato from root pathogens. Plant Signal. Behav. 2018, 13, e1464855. [Google Scholar] [CrossRef]
- Hao, Z.P.; van Tuinen, D.; Fayolle, L.; Chatagnier, O.; Li, X.L.; Chen, B.D.; Gianinazzi, S.; Gianinazzi-Pearson, V. Arbuscular mycorrhiza affects grapevine fanleaf virus transmission by the nematode vector Xiphinema index. Appl. Soil Ecol. 2018, 129, 107–111. [Google Scholar] [CrossRef]
- Eke, P.; Wakam, L.N.; Fokou, P.V.T.; Ekounda, T.V.; Sahu, K.P.; Kamdem Wankeu, T.H.; Boyom, F.F. Improved nutrient status and Fusarium root rot mitigation with an inoculant of two biocontrol fungi in the common bean (Phaseolus vulgaris L.). Rhizosphere 2019, 12, 100172. [Google Scholar] [CrossRef]
- Hao, Z.P.; Christie, P.; Qin, L.; Wang, C.X.; Li, X.L. Control of Fusarium wilt of cucumber seedlings by inoculation with an arbuscular mycorrhical fungus. J. Plant Nutr. 2005, 28, 1961–1974. [Google Scholar] [CrossRef] [Green Version]
- Mora-Romero, G.A.; Cervantes-Gamez, R.G.; Galindo-Flores, H.; Gonzalez-Ortiz, M.A.; Felix-Gastelum, R.; Maldonado-Mendoza, I.E.; Salinas Perez, R.; Leon-Felix, J.; Martinez-Valenzuela, M.C.; Lopez-Meyer, M. Mycorrhiza-induced protection against pathogens is both genotype-specific and graft-transmissible. Symbiosis 2015, 66, 55–64. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, W.; Hao, Z.P.; Yu, M.; Wu, Z.X.; Zhao, A.H.; Li, J.L.; Zhang, X.; Chen, B.D. Improved phosphorus nutrition by arbuscular mycorrhizal symbiosis as a key factor facilitating glycyrrhizin and liquiritin accumulation in Glycyrrhiza uralensis. Plant Soil 2019, 439, 243–257. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; Van Wees, S.C.M.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [Green Version]
- McDowell, J.M. Focus on activation, regulation, and evolution of MTI and ETI. Mol. Plant Microbe. Interact. 2019, 32, 5. [Google Scholar] [CrossRef] [Green Version]
- Kloppholz, S.; Kuhn, H.; Requena, N. A secreted fungal effector of Glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Khaosaad, T.; Garcia-Garrido, J.M.; Steinkellner, S.; Vierheilig, H. Take-all disease is systemically reduced in roots of mycorrhizal barley plants. Soil Biol. Biochem. 2007, 39, 727–734. [Google Scholar] [CrossRef]
- French, K.E. Engineering mycorrhizal symbioses to alter plant metabolism and improve crop health. Front. Microbiol. 2017, 8, 1403. [Google Scholar] [CrossRef]
- Dowarah, B.; Gill, S.S.; Agarwala, N. Arbuscular mycorrhizal fungi in conferring tolerance to biotic stresses in plants. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Schoenherr, A.P.; Rizzo, E.; Jackson, N.; Manosalva, P.; Gomez, S.K. Mycorrhiza-induced resistance in potato involves priming of defense responses against cabbage looper (Noctuidae: Lepidoptera). Environ. Entomol. 2019, 48, 370–381. [Google Scholar] [CrossRef]
- Sanchez-Bel, P.; Troncho, P.; Gamir, J.; Pozo, M.J.; Camanes, G.; Cerezo, M.; Flors, V. The nitrogen availability interferes with mycorrhiza-induced resistance against Botrytis cinerea in tomato. Front. Microbiol. 2016, 7, 1598. [Google Scholar] [CrossRef] [Green Version]
- Formenti, L.; Rasmann, S. Mycorrhizal fungi enhance resistance to herbivores in tomato plants with reduced jasmonic acid production. Agronomy 2019, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Minton, M.M.; Barber, N.A.; Gordon, L.L. Effects of arbuscular mycorrhizal fungi on herbivory defense in two Solanum (Solanaceae) species. Plant Ecol. Evol. 2016, 149, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6, 786. [Google Scholar] [CrossRef] [Green Version]
- Meza, S.L.R.; Tobaruela, E.D.; Pascoal, G.B.; Massaretto, I.L.; Purgatto, E. Post-harvest treatment with methyl jasmonate impacts lipid metabolism in tomato pericarp (Solanum lycopersicum L. cv. Grape) at different ripening stages. Foods 2021, 10, 877. [Google Scholar] [CrossRef]
- Gruden, K.; Lidoy, J.; Petek, M.; Podpe, V.; Flors, V.; Papadopoulou, K.K.; Pappas, M.L.; Martinez-Medina, A.; Bejarano, E.; Biere, A.; et al. Ménage à Trois: Unraveling the mechanisms regulating plant-microbe-arthropod interactions. Trends Plant Sci. 2020, 25, 1215–1226. [Google Scholar] [CrossRef]
- Nair, A.; Kolet, S.P.; Thulasiram, H.V.; Bhargava, S. Systemic jasmonic acid modulation in mycorrhizal tomato plants and its role in induced resistance against Alternaria alternata. Plant Biol. 2015, 17, 625–631. [Google Scholar] [CrossRef]
- Ding, P.; Ding, Y. Stories of salicylic acid: A plant defense hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.H.; Zhai, Q.Z.; Wei, J.N.; Li, S.Y.; Wang, B.; Huang, T.T.; Du, M.M.; Sun, J.Q.; Kang, L.; Li, C.B.; et al. Role of tomato lipoxygenase d in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genet. 2013, 9, e1003964. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, I.; Takehara, Y.; Ota, M.; Imada, K.; Sasaki, K.; Kajihara, H.; Sakai, S.; Jogaiah, S.; Ito, S.I. Magnesium oxide induces immunity against Fusarium wilt by triggering the jasmonic acid signaling pathway in tomato. J. Biotechnol. 2021, 325, 100–108. [Google Scholar] [CrossRef]
- Ouyang, S.Q.; Park, G.; Atamian, H.S.; Han, C.S.; Stajich, J.E.; Kaloshian, I.; Borkovich, K.A. MicroRNAs suppress nb domain genes in tomato that confer resistance to Fusarium oxysporum. PLoS Pathog. 2014, 10, e1004464. [Google Scholar] [CrossRef]
- Chen, S.C.; Ren, J.J.; Zhao, H.J.; Wang, X.L.; Wang, T.H.; Jin, S.D.; Wang, Z.H.; Li, C.Y.; Liu, A.R.; Lin, X.M.; et al. Trichoderma harzianum improves defense against Fusarium oxysporum by regulating ROS and RNS metabolism, redox balance, and energy flow in cucumber roots. Phytopathology 2019, 109, 972–982. [Google Scholar] [CrossRef]
- Biermann, B.; Linderman, R.G. Quantifying vesicular-arbuscular mycorrhizae—A proposed method towards standardization. New Phytol. 1981, 81, 63–67. [Google Scholar] [CrossRef]
- Trouvelot, A.; Fardeau, J.C.; Plenchette, C.; Gianinazzi, S.; Gianinazza-pearson, V. Nutritional balance and symbiotic expression in mycorrhizal wheat. Physiol. Veg. 1986, 24, 300. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Song, Y.Y.; Ye, M.; Li, C.Y.; Wang, R.L.; Wei, X.C.; Luo, S.M.; Zeng, R.S. Priming of anti-herbivore defense in tomato by arbuscular mycorrhizal fungus and involvement of the jasmonate pathway. J. Chem. Ecol. 2013, 39, 1036–1044. [Google Scholar] [CrossRef]
- Zhou, D.X.; Wang, E.Z.; Liu, D.; Jin, C.M.; Li, X.; Jiang, S.; Bai, H.T. Screening of biocontrol bacteria against tomato wilt and the effect on defense enzyme activity of plants. Chin. J. Ecol. 2020, 39, 1753–1760. [Google Scholar]
- Slezack, S.; Dumas-Gaudot, E.; Paynot, M.; Gianinazzi, S. Is a fully established arbuscular mycorrhizal symbiosis required for bioprotection of Pisum sativum roots against Aphanomyces euteiches? Mol. Plant-Microbe Interact. 2000, 13, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Cordier, C.; Pozo, M.J.; Barea, J.M.; Gianinazzi, S.; Gianinazzi-Pearson, V. Cell defense responses associated with localized and systemic resistance to Phytophthora parasitica induced in tomato by an arbuscular mycorrhizal fungus. Mol. Plant-Microbe Interact. 1998, 11, 1017–1028. [Google Scholar] [CrossRef] [Green Version]
- Hause, B.; Schaarschmidt, S. The role of jasmonates in mutualistic symbioses between plants and soil-born microorganisms. Phytochemistry 2009, 70, 1589–1599. [Google Scholar] [CrossRef]
- Tejeda-Sartorius, M.; de la Vega, O.M.; Delano-Frier, J.P. Jasmonic acid influences mycorrhizal colonization in tomato plants by modifying the expression of genes involved in carbohydrate partitioning. Physiol. Plant. 2008, 133, 339–353. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Biere, A.; van der Putten, W.H.; Wagenaar, R.; Klironomos, J.N. Chemical defense, mycorrhizal colonization and growth responses in Plantago lanceolata L. Oecologia 2009, 160, 433–442. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Xie, W.; Hao, Z.P.; Zhou, X.F.; Jiang, X.L.; Xu, L.J.; Wu, S.L.; Zhao, A.H.; Zhang, X.; Chen, B.D. Arbuscular mycorrhiza facilitates the accumulation of glycyrrhizin and liquiritin in Glycyrrhiza uralensis under drought stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef]
- Yu, M.; Xie, W.; Zhang, X.; Zhang, S.B.; Wang, Y.S.; Hao, Z.P.; Chen, B.D. Arbuscular mycorrhizal fungi can compensate for the loss of indigenous microbial communities to support the growth of liquorice (Glycyrrhiza uralensis Fisch.). Plants 2020, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Bernaola, L.; Cosme, M.; Schneider, R.W.; Stout, M. Belowground inoculation with arbuscular mycorrhizal fungi increases local and systemic susceptibility of rice plants to different pest organisms. Front. Plant Sci. 2018, 9, 747. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.M.; Seleim, M.A.A.; Abd-El-Moneem, K.M.H.; Saead, F.A. Integrated effect of Glomus mosseae and selected plant oils on the control of bacterial wilt disease of tomato. Crop Prot. 2014, 66, 67–71. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcon-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Elyousr, K.A.M.; Bagy, H.M.M.K.; Hashem, M.; Alamri, S.A.M.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt. J. Biol. Pest Control. 2019, 29, 54. [Google Scholar] [CrossRef]
- Bereika, F.F.M.; Sallam, N.M.A.; Alamri, S.A.M.; Abo-Elyousr, K.A.M.; Mostafa, Y.S.; Hashem, M. Approving the biocontrol strategy of potato wilt caused by Ralstonia solanacearum on field scale using Enterobacter cloacae PS14 and Trichoderma asperellum T34. Egypt. J. Biol. Pest Control 2020, 30, 1–13. [Google Scholar]
- Tian, L.; Zou, Y.N.; Wu, Q.S.; Kuca, K. Mycorrhiza-induced plant defence responses in trifoliate orange infected by Phytophthora parasitica. Acta. Physiol. Plant. 2021, 43, 45. [Google Scholar] [CrossRef]
- Pozo, M.J.; Verhage, A.; Garcia-Andrade, J.; Garcia, J.M.; Azcon-Aguilar, C. Priming plant defence against pathogens by arbuscular mycorrhizal fungi. In Mycorrhizas-Functional Processes and Ecological Impact; Azcon Aguilar, C., Barea, J.M., Gianinazzi Pearson, V., Gianinazzi, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 9, pp. 123–135. [Google Scholar]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Constabel, C.P. Systemin activates synthesis of wound-inducible tomato leaf polyphenol oxidase via the octadecanoid defense signaling pathway. Proc. Natl. Acad. Sci. USA 1995, 92, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, T.; Bhaskaran, R.; Muthusamy, M. Pseudomonas fluorescens induced enzymological changes in banana roots (Cv. Rasthali) against Fusarium wilt disease. Plant Pathol. J. 2004, 3, 72–80. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Chen, X.L.; Chai, X.F.; Xue, D.Q.; Zheng, W.; Shi, Y.Y.; Wang, A.X. The involvement of jasmonic acid, ethylene, and salicylic acid in the signaling pathway of Clonostachys rosea-induced resistance to gray mold disease in tomato. Phytopathology. 2019, 109, 1102–1114. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Ye, M.; Li, X.W.; Lin, S.B.; Sun, X.L. The jasmonic acid pathway positively regulates the polyphenol oxidase-based defense against tea geometrid caterpillars in the tea plant (Camellia sinensis). J. Chem. Ecol. 2020, 46, 308–316. [Google Scholar] [CrossRef]
- Jaiti, F.; Verdeil, J.L.; El Hadrami, I. Effect of jasmonic acid on the induction of polyphenoloxidase and peroxidase activities in relation to date palm resistance against Fusarium oxysporum f. sp. albedinis. Physiol. Mol. Plant Pathol. 2009, 74, 84–90. [Google Scholar] [CrossRef]
- Campos-Vargas, R.; Saltveit, M.E. Involvement of putative chemical wound signals in the induction of phenolic metabolism in wounded lettuce. Physiol. Plant. 2002, 114, 73–84. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Inoculation | Mycorrhizal Colonization Rate (%) | Arbuscule Abundance (%) |
|---|---|---|---|
| CM | −M−F | 0.0 ± 0.0 d | 0.0 ± 0.0 c |
| +M−F | 58.8 ± 4.0 a | 33.9 ± 4.4 a | |
| −M+F | 0.0 ± 0.0 d | 0.0 ± 0.0 c | |
| +M+F | 50.5 ± 3.1 b | 38.1 ± 1.6 a | |
| Spr8 | −M−F | 0.0 ± 0.0 d | 0.0 ± 0.0 c |
| +M−F | 31.6 ± 3.5 c | 13.7 ± 2.1 b | |
| −M+F | 0.0 ± 0.0 d | 0.0 ± 0.0 c | |
| +M+F | 32.7±1.4 c | 11.4 ± 1.6 b | |
| Significance of | |||
| G | p < 0.001 ** | p < 0.001 ** | |
| Myc | p < 0.001 ** | p < 0.001 ** | |
| Fol | p = 0.219 | p = 0.861 | |
| G × Myc | p < 0.001 ** | p < 0.001 ** | |
| G × Fol | p = 0.163 | p = 0.229 | |
| Myc × Fol | p = 0.219 | p = 0.861 | |
| G × Myc × Fol | p = 0.163 | p = 0.229 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Hao, Z.; Zhang, X.; Xie, W.; Chen, B. Arbuscular Mycorrhizal Fungi Induced Plant Resistance against Fusarium Wilt in Jasmonate Biosynthesis Defective Mutant and Wild Type of Tomato. J. Fungi 2022, 8, 422. https://doi.org/10.3390/jof8050422
Wang H, Hao Z, Zhang X, Xie W, Chen B. Arbuscular Mycorrhizal Fungi Induced Plant Resistance against Fusarium Wilt in Jasmonate Biosynthesis Defective Mutant and Wild Type of Tomato. Journal of Fungi. 2022; 8(5):422. https://doi.org/10.3390/jof8050422
Chicago/Turabian StyleWang, Haixi, Zhipeng Hao, Xin Zhang, Wei Xie, and Baodong Chen. 2022. "Arbuscular Mycorrhizal Fungi Induced Plant Resistance against Fusarium Wilt in Jasmonate Biosynthesis Defective Mutant and Wild Type of Tomato" Journal of Fungi 8, no. 5: 422. https://doi.org/10.3390/jof8050422