
Heterologous Expression of Secondary Metabolite Genes in Trichoderma reesei for Waste Valorization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbiological Methods
2.2. Molecular Biology Methods
2.3. Fermentation
2.4. Extraction of Transformants Grown on Waste Materials
2.5. Chemical Analysis
3. Results
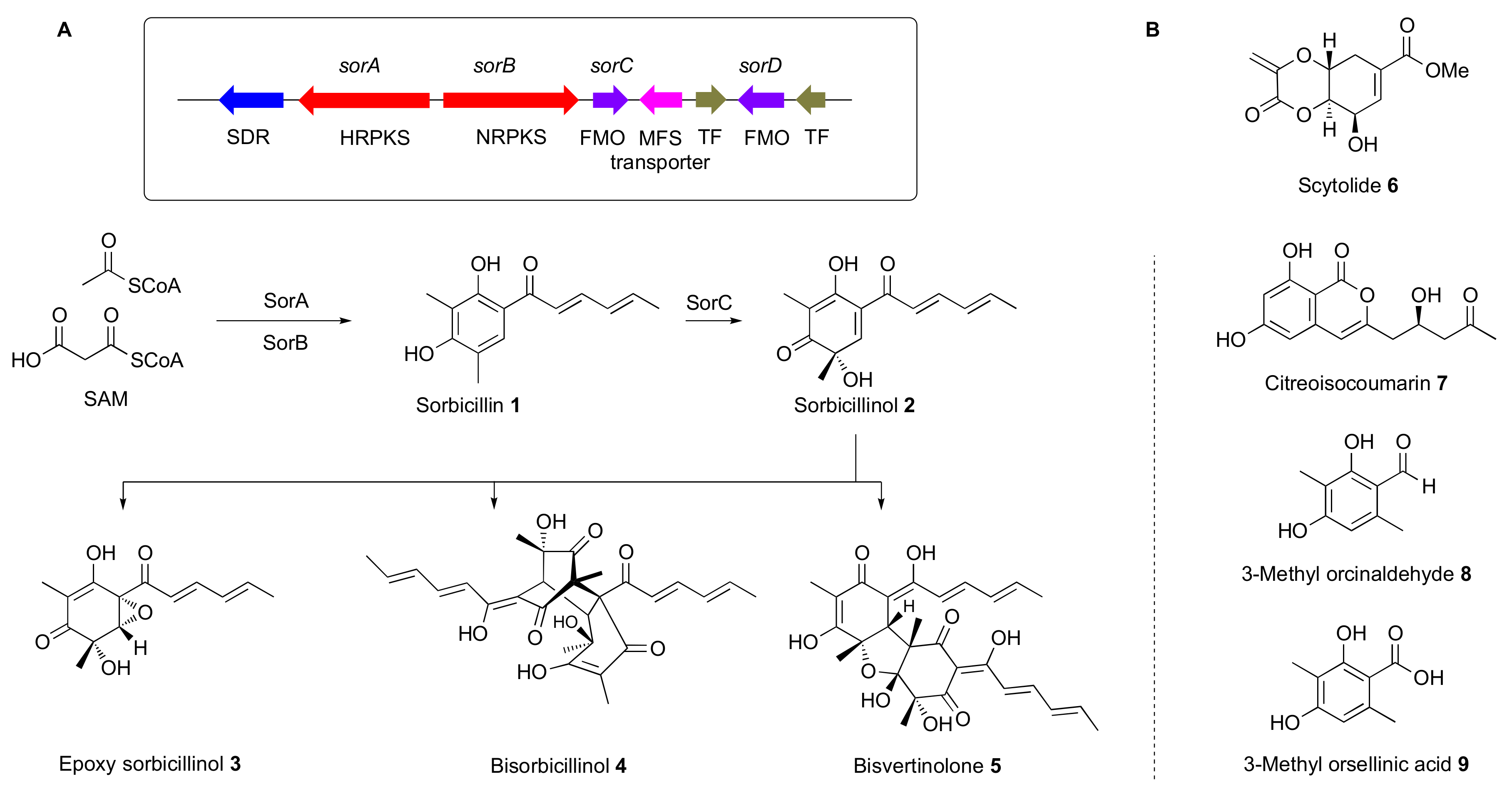
3.1. Construction of a Host with a Cleaner Secondary Metabolic Background
3.2. Construction of a T. reesei Heterologous Expression Vector
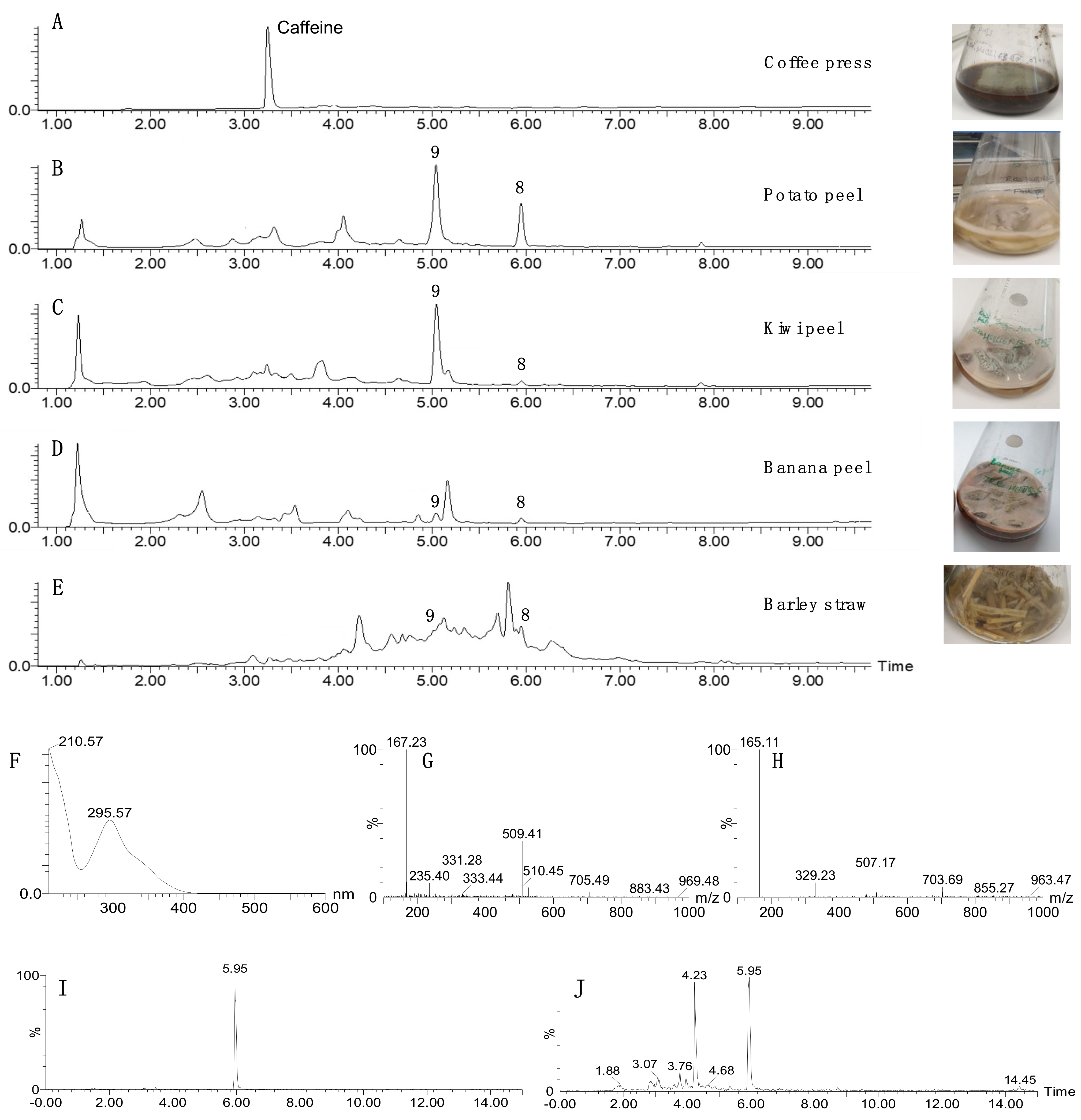
3.3. Growing Recombinant T. reesei Strains on Different Waste Materials
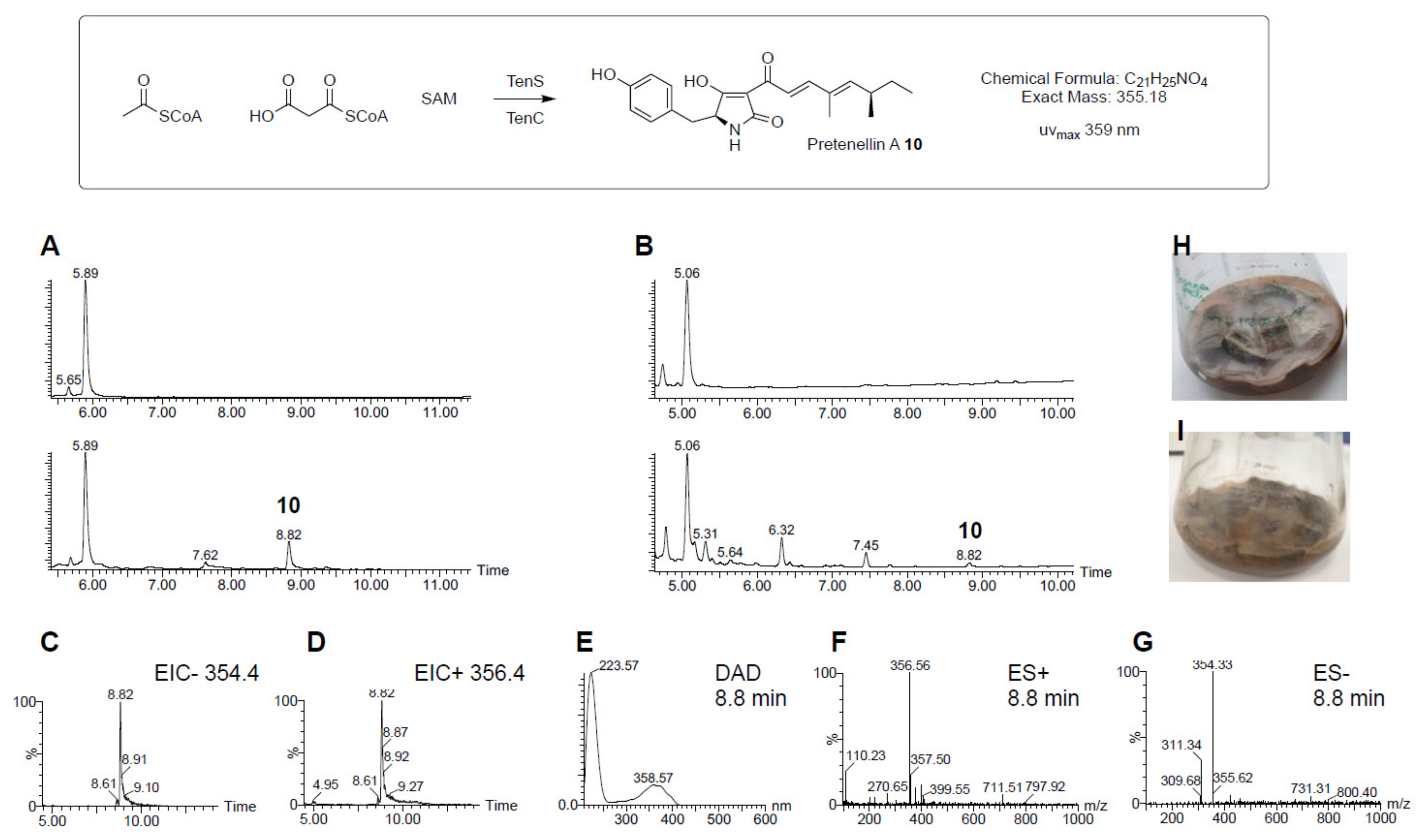
3.4. Construction of a Multiple-Gene Expression System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazarus, C.M.; Williams, K.; Bailey, A.M. Reconstructing fungal natural product biosynthetic pathways. Nat. Prod. Rep. 2014, 31, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Shenouda, M.L.; Cox, R.J. Molecular methods unravel the biosynthetic potential of Trichoderma species. RSC Adv. 2021, 11, 3622–3635. [Google Scholar] [CrossRef]
- Kahlert, L.; Schotte, C.; Cox, R.J. Total Mycosynthesis: Rational Bioconstruction and Bioengineering of Fungal Natural Products. Synthesis 2021, 53, 2381–2394. [Google Scholar] [CrossRef]
- Yuet, K.P.; Khosla, C. Challenges and opportunities for engineering assembly-line polyketide biosynthesis in Escherichia coli. Metab. Eng. Commun. 2020, 10, e00106. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.J.B.; Tang, M.; Schlecht, U.; Horecka, J.; Fischer, C.R.; Lin, H.-C.; Li, J.; Naughton, B.; Cherry, J.; Miranda, M.; et al. HEx: A Heterologous Expression Platform for the Discovery of Fungal Natural Products. Sci. Adv. 2018, 4, eaar5459. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Oakley, C.E.; Ahuja, M.; Entwistle, R.; Schultz, A.; Chang, S.L.; Sung, C.T.; Wang, C.C.C.; Oakley, B.R. An efficient system for heterologous expression of secondary metabolite genes in Aspergillus nidulans. J. Am. Chem. Soc. 2013, 135, 7720–7731. [Google Scholar] [CrossRef] [Green Version]
- Vandova, G.A.; O’Brien, R.V.; Lowry, B.; Robbins, T.F.; Fischer, C.R.; Davis, R.W.; Khosla, C.; Harvey, C.J.; Hillenmeyer, M.E. Heterologous Expression of Diverse Propionyl-CoA Carboxylases Affects Polyketide Production in Escherichia coli. J. Antibiot. 2017, 70, 859–863. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Hauser, M.; Cox, R.J.; Skellam, E. Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase. J. Fungi 2021, 7, 1085. [Google Scholar] [CrossRef]
- Song, Z.; Bakeer, W.; Marshall, J.W.; Yakasai, A.A.; Khalid, R.M.; Collemare, J.; Skellam, E.; Tharreau, D.; Lebrun, M.H.; Lazarus, C.M.; et al. Heterologous expression of the avirulence gene ACE1 from the fungal rice pathogen Magnaporthe oryzae. Chem. Sci. 2015, 6, 4837–4845. [Google Scholar] [CrossRef] [Green Version]
- Xiang, P.; Ludwig-Radtke, L.; Yin, W.B.; Li, S.M. Isocoumarin formation by heterologous gene expression and modification by host enzymes. Org. Biomol. Chem. 2020, 18, 4946–4948. [Google Scholar] [CrossRef]
- Shenouda, M.L.; Ambilika, M.; Cox, R.J. Trichoderma reesei Contains a Biosynthetic Gene Cluster That Encodes the Antifungal Agent Ilicicolin H. J. Fungi 2021, 7, 1034. [Google Scholar] [CrossRef] [PubMed]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevalainen, H.; Suominen, P.; Taimisto, K. On the safety of Trichoderma reesei. J. Biotechnol. 1994, 37, 193–200. [Google Scholar] [CrossRef]
- Weiss, R.; Eischer, A.; Tadic, T.; Gritsch, S.M.; Ortner, M.; Prall, K.; Neunteufel, E.; Putz, R.F.; Guebitz, G.M.; Nyanhongo, G.S. Valorisation of slaughterhouse and deinking paper waste streams for the production of enzyme by Trichoderma reesei. J. Clean. Prod. 2020, 275, 122882. [Google Scholar] [CrossRef]
- Wang, H.; Kaur, G.; Pensupa, N.; Uisan, K.; Du, C.; Yang, X.; Lin, C.S.K. Textile waste valorization using submerged filamentous fungal fermentation. Process. Saf. Environ. Prot. 2018, 118, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.T.; Pham, T.T.H.; Adjallé, K.; Montplaisir, D.; Brouillette, F.; Barnabé, S. Production of Trichoderma reesei RUT C-30 Lignocellulolytic Enzymes Using Paper Sludge as Fermentation Substrate: An Approach for On-Site Manufacturing of Enzymes for Biorefineries. Waste Biomass Valorization 2016, 8, 1081–1088. [Google Scholar] [CrossRef]
- Mandels, M.; Hontz, L.; Nystrom, J.; Lee, R.; Lynd, I. Enzymatic hydrolysis of waste cellulose. Biotechnol. Bioeng. 2010, 105, 3–25. [Google Scholar] [CrossRef]
- Liu, P.; Lin, A.; Zhang, G.; Zhang, J.; Chen, Y.; Shen, T.; Zhao, J.; Wei, D.; Wang, W. Enhancement of Cellulase Production in Trichoderma Reesei RUT-C30 by Comparative Genomic Screening. Microb. Cell Factories 2019, 18, 81. [Google Scholar] [CrossRef]
- Derntl, C.; Rassinger, A.; Srebotnik, E.; Mach, R.L.; Mach-Aigner, A.R. Identification of the main regulator responsible for synthesis of the typical yellow pigment produced by Trichoderma reesei. Appl. Environ. Microbiol. 2016, 82, 6247–6257. [Google Scholar] [CrossRef] [Green Version]
- Derntl, C.; Guzmán-Chávez, F.; Mello-de-Sousa, T.M.; Busse, H.J.; Driessen, A.J.M.; Mach, R.L.; Mach-Aigner, A.R. In vivo study of the sorbicillinoid gene cluster in Trichoderma reesei. Front. Microbiol. 2017, 8, 2037. [Google Scholar] [CrossRef]
- Kahlert, L.; Bassiony, E.F.; Cox, R.J.; Skellam, E.J. Diels–Alder Reactions During the Biosynthesis of Sorbicillinoids. Angew. Chem. 2020, 132, 5865–5871. [Google Scholar] [CrossRef]
- Nielsen, M.L.; Albertsen, L.; Lettier, G.; Nielsen, J.B.; Mortensen, U.H. Efficient PCR-Based Gene Targeting with a Recyclable Marker for Aspergillus nidulans. Fung. Genet. Biol. 2006, 43, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Szwalbe, A.J.; Mulholland, N.P.; Vincent, J.L.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Heterologous Production of Fungal Maleidrides Reveals the Cryptic Cyclization Involved in Their Biosynthesis. Angew. Chem. 2016, 55, 6784–6788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gritz, L.; Davies, J. Plasmid-encoded hygromycin B resistance: The sequence of hygromycin B phosphotransferase gene and its expression in Escherichia coli and Saccharomyces cerevisiae. Gene 1983, 25, 179–188. [Google Scholar] [CrossRef]
- Derntl, C.; Kiesenhofer, D.P.; Mach, R.L.; Mach-Aigner, A.R. Novel Strategies for Genomic Manipulation of Trichoderma reesei with the Purpose of Strain Engineering. Appl. Environ. Microbiol. 2015, 81, 6314–6323. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Ono, Y.; Fujii, I.; Sankawa, U.; Mayorga, M.E.; Timberlake, W.E.; Ebizuka, Y. Product Identification of Polyketide Synthase Coded by Aspergillus nidulans WA Gene. Tetrahedron Lett. 1998, 39, 7733–7736. [Google Scholar] [CrossRef]
- Nofiani, R.; de Mattos-Shipley, K.; Lebe, K.E.; Han, L.-C.; Iqbal, Z.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Strobilurin biosynthesis in Basidiomycete fungi. Nat. Commun. 2018, 9, 3940. [Google Scholar] [CrossRef] [Green Version]
- Schor, R.; Schotte, C.; Wibberg, D.; Kalinowski, J.; Cox, R.J. Three previously unrecognised classes of biosynthetic enzymes revealed during the production of xenovulene A. Nat. Commun. 2018, 9, 1963. [Google Scholar] [CrossRef]
- Pahirulzaman, K.A.K.; Williams, K.; Lazarus, C.M. A Toolkit for Heterologous Expression of Metabolic Pathways in Aspergillus oryzae. Methods Enzymol. 2012, 517, 241–260. [Google Scholar] [CrossRef]
- Bailey, A.M.; Cox, R.J.; Harley, K.; Lazarus, C.M.; Simpson, T.J.; Skellam, E. Characterisation of 3-methylorcinaldehyde synthase (MOS) in Acremonium strictum: First observation of a reductive release mechanism during polyketide biosynthesis. Chem. Commun. 2007, 4053–4055. [Google Scholar] [CrossRef]
- Penttilä, M.; Nevalainen, H.; Rättö, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Wang, S.; Xing, M.; Yu, S.; Liu, G. Achieving efficient protein expression in Trichoderma reesei by using strong constitutive promoters. Microb. Cell Factories 2012, 11, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halo, L.M.; Marshall, J.W.; Yakasai, A.A.; Song, Z.; Butts, C.P.; Crump, M.P.; Heneghan, M.; Bailey, A.M.; Simpson, T.J.; Lazarus, C.M.; et al. Authentic Heterologous Expression of the Tenellin Iterative Polyketide Synthase Nonribosomal Peptide Synthetase Requires Coexpression with an Enoyl Reductase. ChemBioChem 2008, 9, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, M.N.; Yakasai, A.A.; Halo, L.M.; Song, Z.; Bailey, A.M.; Simpson, T.J.; Cox, R.J.; Lazarus, C.M. First heterologous reconstruction of a complete functional fungal biosynthetic multigene cluster. ChemBioChem 2010, 11, 1508–1512. [Google Scholar] [CrossRef]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Fitz, E.; Wanka, F.; Seiboth, B. The promoter toolbox for recombinant gene expression in Trichoderma reesei. Front. Bioeng. Biotechnol. 2018, 6, 135. [Google Scholar] [CrossRef]
- Jørgensen, M.S.; Skovlund, D.A.; Johannesen, P.F.; Mortensen, U.H. A novel platform for heterologous gene expression in Trichoderma reesei (Teleomorph Hypocrea jecorina). Microb. Cell Factories 2014, 13, 33. [Google Scholar] [CrossRef] [Green Version]
- Marra, R.; Nicoletti, R.; Pagano, E.; DellaGreca, M.; Salvatore, M.M.; Borrelli, F.; Lombardi, N.; Vinale, F.; Woo, S.L.; Andolfi, A. Inhibitory effect of trichodermanone C, a sorbicillinoid produced by Trichoderma citrinoviride associated to the green alga Cladophora sp., on nitrite production in LPS-stimulated macrophages. Nat. Prod. Res. 2018, 33, 3389–3397. [Google Scholar] [CrossRef]
- Sugaya, K.; Koshino, H.; Hongo, Y.; Yasunaga, K.; Onose, J.; Yoshikawa, K.; Abe, N. The biosynthesis of sorbicillinoids in Trichoderma sp. USF-2690: Prospect for the existence of a common precursor to sorbicillinol and 5-epihydroxyvertinolide, a new sorbicillinoid member. Tetrahedron Lett. 2008, 49, 654–657. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Kubicek, E.M.; Kubicek, C.P. Several steps of lateral gene transfer followed by events of “birth-and-death” evolution shaped a fungal sorbicillinoid biosynthetic gene cluster. BMC Evol. Biol. 2016, 16, 269. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Tian, L.; Huang, J.; Ma, H.Y.; Zheng, Z.; Lv, A.L.; Yasukawa, K.; Pei, Y.H. Trichodermatides A-D, novel polyketides from the marine-derived fungus Trichoderma reesei. Org. Lett. 2008, 10, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tian, L.; Huang, Y.F.; Sha, Y.; Pei, Y.H. A new cyclotetrapeptide from marine fungus Trichoderma reesei. Pharmazie 2006, 61, 809–810. [Google Scholar] [CrossRef] [PubMed]
- Salo, O.; Guzmán-Chávez, F.; Ries, M.I.; Lankhorst, P.P.; Bovenberg, R.A.L.; Vreeken, R.J.; Driessen, A.J.M. Identification of a Polyketide Synthase Involved in Sorbicillin Biosynthesis by Penicillium chrysogenum. Appl. Environ. Microbiol. 2016, 82, 3971–3978. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; An, N.; Guo, J.; Wang, Z.; Meng, X.; Liu, W. Influences of genetically perturbing synthesis of the typical yellow pigment on conidiation, cell wall integrity, stress tolerance, and cellulase production in Trichoderma reesei. J. Microbiol. 2021, 59, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Zhao, J.; Ding, L.; Huang, C.; Naman, C.B.; He, S.; Wu, B.; Zhu, P.; Luo, Q.; Gerwick, W.H.; et al. 5-Hydroxycyclopenicillone, a New β-Amyloid Fibrillization Inhibitor from a Sponge-Derived Fungus Trichoderma sp. HPQJ-34. Mar. Drugs 2017, 15, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, M.; Chiang, Y.-M.; Chang, S.-L.; Praseuth, M.B.; Entwistle, R.; Sanchez, J.F.; Lo, H.-C.; Yeh, H.-H.; Oakley, B.R.; Wang, C.C.C. Illuminating the Diversity of Aromatic Polyketide Synthases in Aspergillus nidulans. J. Am. Chem. Soc. 2012, 134, 8212–8221. [Google Scholar] [CrossRef] [Green Version]
- Chooi, Y.-H.; Muria-Gonzalez, M.; Mead, O.L.; Solomon, P.S. SnPKS19 Encodes the Polyketide Synthase for Alternariol Mycotoxin Biosynthesis in the Wheat Pathogen Parastagonospora nodorum. Appl. Environ. Microbiol. 2015, 81, 5309–5317. [Google Scholar] [CrossRef] [Green Version]
- Velázquez-Nuñez, M.J.; Avila-Sosa, R.; Palou, E.; López-Malo, A. Antifungal activity of orange (Citrus sinensis var. Valencia) peel essential oil applied by direct addition or vapor contact. Food Control 2013, 31, 1–4. [Google Scholar] [CrossRef]
- Caccioni, D.R.L.; Guizzardi, M.; Biondi, D.M.; Renda, A.; Ruberto, G. Relationship between volatile components of citrus fruit essential oils and antimicrobial action on Penicillium digitatum and Penicillium italicum. Int. J. Food Microbiol. 1998, 43, 73–79. [Google Scholar] [CrossRef]
- Dashtban, M.; Schraft, H.; Syed, T.A.; Qin, W. Fungal Biodegradation and Enzymatic Modification of Lignin. Int. J. Biochem. Mol. Biol. 2010, 1, 36–50. [Google Scholar]
- Matsuda, Y.; Wakimoto, T.; Mori, T.; Awakawa, T.; Abe, I. Complete Biosynthetic Pathway of Anditomin: Nature’s Sophisticated Synthetic Route to a Complex Fungal Meroterpenoid. J. Am. Chem. Soc. 2014, 136, 15326–15336. [Google Scholar] [CrossRef] [PubMed]
- Van Dijken, J.P.; Bauer, J.; Brambilla, L.; Duboc, P.; Francois, J.M.; Gancedo, C.; Giuseppin, M.L.F.; Heijnen, J.J.; Hoare, M.; Lange, H.C.; et al. An interlaboratory comparison of physiological and genetic properties of four Saccharomyces cerevisiae strains. Enzyme Microb. Technol. 2000, 26, 706–714. [Google Scholar] [CrossRef]
- Yang, X.-L.; Friedrich, S.; Yin, S.; Piech, O.; Williams, K.; Simpson, T.J.; Cox, R.J. Molecular basis of methylation and chain-length programming in a fungal iterative highly reducing polyketide synthase. Chem. Sci. 2019, 10, 8478–8489. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Gruber, F.; Visser, J.; Kubicek, C.P.; de Graaff, L.H. The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain. Curr. Genet. 1990, 18, 71–76. [Google Scholar] [CrossRef]
- Dockrey, S.A.B.; Narayan, A.R.H. Photocatalytic Oxidative Dearomatization of Orcinaldehyde Derivatives. Org. Lett. 2020, 22, 3712–3716. [Google Scholar] [CrossRef]
- Shang, Z.; Khalil, Z.; Li, L.; Salim, A.A.; Quezada, M.; Kalansuriya, P.; Capon, R.J. Roseopurpurins: Chemical Diversity Enhanced by Convergent Biosynthesis and Forward and Reverse Michael Additions. Org. Lett. 2016, 18, 4340–4343. [Google Scholar] [CrossRef]
- Hantke, V.; Wang, C.; Skellam, E.J.; Cox, R.J. Function of pathway specific regulators in the ACE1 and pyrichalasin H biosynthetic gene clusters. RSC Adv. 2019, 9, 35797–35802. [Google Scholar] [CrossRef] [Green Version]
- Uzbas, F.; Sezerman, U.; Hartl, L.; Kubicek, C.P.; Seiboth, B. A homologous production system for Trichoderma reesei secreted proteins in a cellulase-free background. Appl. Microbiol. Biotechnol. 2012, 93, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Ohashi, M.; Hung, Y.-S.; Scherlach, K.; Watanabe, K.; Hertweck, C.; Tang, Y. AoiQ Catalyzes Geminal Dichlorination of 1,3-Diketone Natural Products. J. Am. Chem. Soc. 2021, 143, 7267–7271. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of Microbial Natural Products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Nakari-Setala, T.; Penttila, M. Production of Trichoderma reesei cellulases on glucose-containing media. Appl. Environ. Microbiol. 1995, 61, 3650–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shenouda, M.L.; Ambilika, M.; Skellam, E.; Cox, R.J. Heterologous Expression of Secondary Metabolite Genes in Trichoderma reesei for Waste Valorization. J. Fungi 2022, 8, 355. https://doi.org/10.3390/jof8040355
Shenouda ML, Ambilika M, Skellam E, Cox RJ. Heterologous Expression of Secondary Metabolite Genes in Trichoderma reesei for Waste Valorization. Journal of Fungi. 2022; 8(4):355. https://doi.org/10.3390/jof8040355
Chicago/Turabian StyleShenouda, Mary L., Maria Ambilika, Elizabeth Skellam, and Russell J. Cox. 2022. "Heterologous Expression of Secondary Metabolite Genes in Trichoderma reesei for Waste Valorization" Journal of Fungi 8, no. 4: 355. https://doi.org/10.3390/jof8040355