
A Plant-Derived Alkanol Induces Teliospore Germination in Sporisorium scitamineum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sugarcane Varieties and Sporisorium scitamineum Isolate
2.2. Evaluation of the Germination of Smut Teliospores on Sugarcane Buds
2.3. Scanning Electron Microscopy
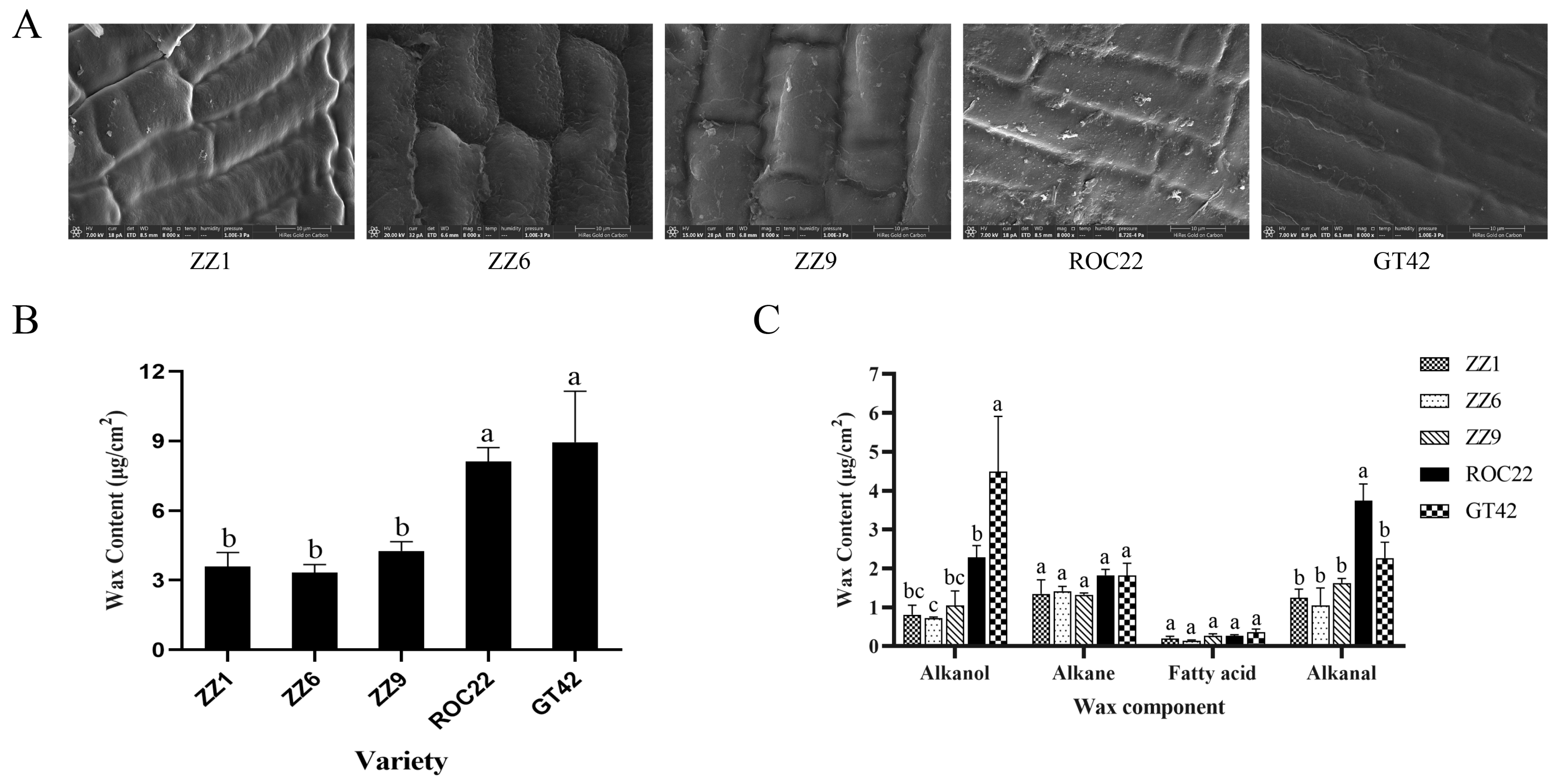
2.4. Determination of Wax Content
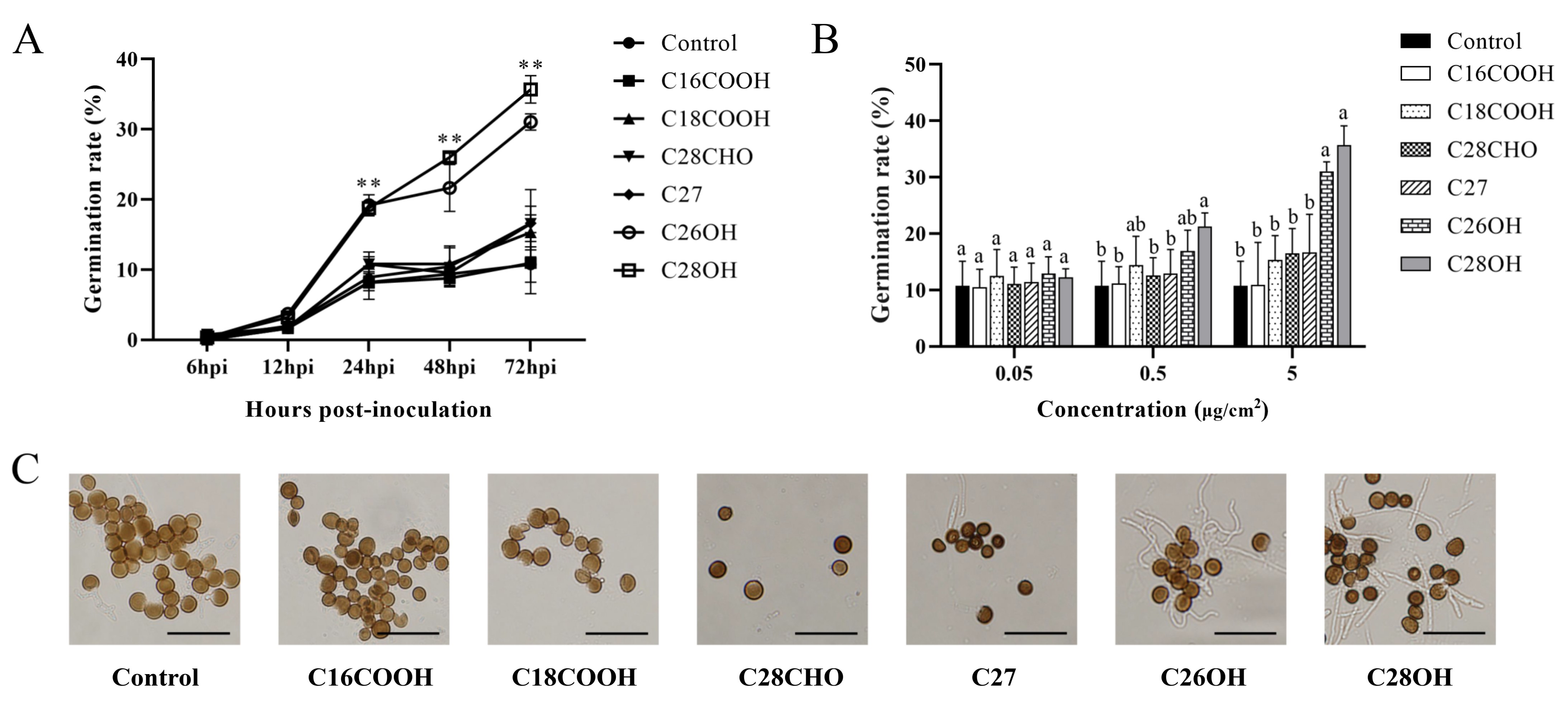
2.5. Smut Teliospores Germination on Pure Wax Coating Surfaces
2.6. RNA Sequencing
2.7. Quantitative Reverse Transcription PCR Analysis
3. Results
3.1. Smut Teliospores Germinates at Varied Rates on the Buds of Smut-Resistant and Smut-Susceptible Sugarcane Varieties
3.2. The Alkanol Contents of the Buds Differ between Smut-Resistant and Smut-Susceptible Sugarcane Varieties
3.3. Correlation between Alkanol Content and Teliospore Germination Is Validated in Clones of Sub-Genetic Populations
3.4. Hexacosanol and Octacosanol Induce Smut Teliospore Germination In Vitro
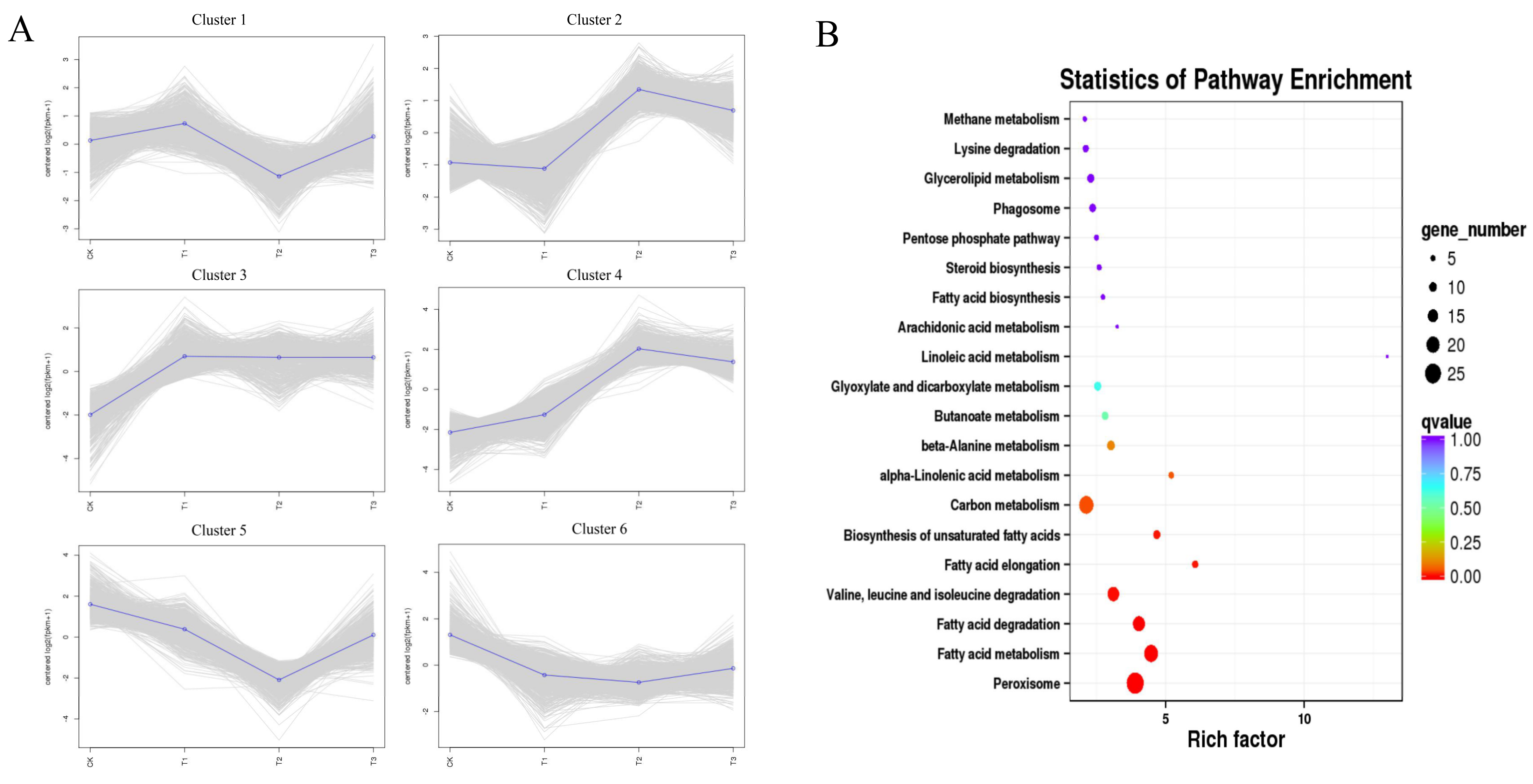
3.5. The MAPK Signaling and Fatty Acid Metabolism Pathways Are Specifically Regulated after Induction by Octacosanol
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Solomon, S. Sugarcane By-Products Based Industries in India. Sugar Tech 2011, 13, 408–416. [Google Scholar] [CrossRef]
- Bhuiyan, S.A.; Magarey, R.; McNeil, M.; Aitken, K. Sugarcane smut, caused by Sporisorium scitamineum, a major disease of sugarcane—A contemporary review. Phytopathology 2021, 111, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Da Gloria, B.A.; Albernas, M.C.C.; Amorim, L. Structural characteristics of buds of sugarcane cultivars with different levels for resistance to smut. J. Plant Dis. Prot. 1995, 102, 502–508. [Google Scholar]
- De Armas, R.; Santiago, R.; Legaz, M.E.; Vicente, C. Levels of phenolic compounds and enzyme activity can be used to screen for resistance of sugarcane to smut (Ustilago scitaminea). Australas. Plant Pathol. 2007, 36, 32–38. [Google Scholar] [CrossRef]
- Su, Y.; Wang, Z.; Xu, L.; Peng, Q.; Liu, F.; Li, Z.; Que, Y. Early Selection for Smut Resistance in Sugarcane Using Pathogen Proliferation and Changes in Physiological and Biochemical Indices. Front. Plant Sci. 2016, 7, 1133. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Somai, B.M.; Pillay, D. Smut disease assessment by PCR and microscopy in inoculated tissue cultured sugarcane cultivars. Plant Sci. 2004, 167, 987–994. [Google Scholar] [CrossRef]
- Flores, C.I.; Carpena, A.L.; Rosario, E.L. Evaluation of sugarcane hybrids for resistance to sugarcane smut (Ustilago scitaminea Sydow). Philipp. J. Crop Sci. 1978, 3, 121–125. [Google Scholar]
- Prakasam, P.; Sarma, M.N. Influence of host contact on the germination of Sugarcane smut spores. Sci. Cult. 1960, 3, 644–645. [Google Scholar]
- Marques, J.; Hoy, J.W.; Appezzato-da-Glória, B.; Viveros, A.; Vieira, M.; Baisakh, N. Sugarcane Cell Wall-Associated Defense Responses to Infection by Sporisorium scitamineum. Front. Plant Sci. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Gomez, S.M.; Kumar, S.S.; Ganesh, S.K.; Biji, K.R.; Senthil, A.; Babu, R.C. QTLs linked to leaf epicuticular wax, physio-morphological and plant production traits under drought stress in rice (Oryza sativa L.). Plant Growth Regul. 2008, 56, 245–256. [Google Scholar] [CrossRef]
- Zhu, X.; Xiong, L. Putative megaenzyme DWA1 plays essential roles in drought resistance by regulating stress-induced wax deposition in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 17790–17795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, R.S.; van Staden, J. Towards a rapid near-infrared technique for prediction of resistance to sugarcane borer Eldana saccharina walker (Lepidoptera: Pyralidae) using stalk surface wax. J. Chem. Ecol. 1996, 22, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Purcell, D.E.; Leonard, G.J.; O'Shea, M.G.; Kokot, S. A chemometrics investigation of sugarcane plant properties based on the molecular composition of epicuticular wax. Chemometr. Intell. Lab. 2005, 76, 135–147. [Google Scholar] [CrossRef]
- Lamberton, J.A.; Redcliffe, A.H. The Chemistry of Sugar-Cane Wax. I. The Nature of Sugar-Cane Wax. Aust. J. Chem. 1960, 13, 261–268. [Google Scholar] [CrossRef]
- Shanmugam, M.; Chithra, L.; Nageswari, R.; Venkatesan, M.S.; Mathialagan, M. Sugarcane Wax—A Par Excellent by-Product of Sugar Industry—A Review. Agric. Rev. 2021, 42, 315–321. [Google Scholar]
- Koch, K.; Hartmann, K.D.; Schreiber, L.; Barthlott, W.; Neinhuis, C. Influences of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Environ. Exp. Bot. 2006, 56, 1–9. [Google Scholar] [CrossRef]
- Post-Beittenmiller, D. Biochemistry and Molecular Biology of Wax Production in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 405–430. [Google Scholar] [CrossRef] [Green Version]
- Gniwotta, F.; Vogg, G.; Gartmann, V.; Carver, T.L.; Riederer, M.; Jetter, R. What do microbes encounter at the plant surface? Chemical composition of pea leaf cuticular waxes. Plant Physiol. 2005, 139, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Eigenbrode, S.D.; Espelie, K.E. Effects of Plant Epicuticular Lipids on Insect Herbivores. Annu. Rev. Entomol. 1995, 40, 171–194. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hansjakob, A.; Bischof, S.; Bringmann, G.; Riederer, M.; Hildebrandt, U. Very-long-chain aldehydes promote in vitro prepenetration processes of Blumeria graminis in a dose- and chain length-dependent manner. New Phytol. 2010, 188, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Uppalapati, S.R.; Ishiga, Y.; Doraiswamy, V.; Bedair, M.; Mittal, S.; Chen, J.; Nakashima, J.; Tang, Y.; Tadege, M.; Ratet, P.; et al. Loss of abaxial leaf epicuticular wax in Medicago truncatula irg1/palm1 mutants results in reduced spore differentiation of anthracnose and nonhost rust pathogens. Plant Cell 2012, 24, 353–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.; Ling, H.; Liu, F.; Su, Y.; Su, W.; Mao, H.; Zhang, X.; Wang, L.; Chen, R.; Que, Y. Identification and evaluation of PCR reference genes for host and pathogen in sugarcane-Sporisorium scitamineum interaction system. BMC Genom. 2018, 19, 479. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Peters, L.P.; Carvalho, G.; Vilhena, M.B.; Creste, S.; Azevedo, R.A.; Monteiro-Vitorello, C.B. Functional analysis of oxidative burst in sugarcane smut-resistant and -susceptible genotypes. Planta 2017, 245, 749–764. [Google Scholar] [CrossRef] [Green Version]
- Russin, J.S.; Guo, B.Z.; Tubajika, K.M.; Brown, R.L.; Cleveland, T.E.; Widstrom, N.W. Comparison of Kernel Wax from Corn Genotypes Resistant or Susceptible to Aspergillus flavus. Phytopathology 1997, 87, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Zinsou, V.; Wydra, K.; Ahohuendo, B.; Schreiber, L. Leaf Waxes of Cassava (Manihot Esculenta Crantz) in Relation to Ecozone and Resistance to Xanthomonas Blight. Euphytica 2006, 149, 189–198. [Google Scholar] [CrossRef]
- Sage, R.; Peixoto, M.; Sage, T. Sugarcane: Physiology, Biochemistry, and Functional Biology; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar]
- Asikin, Y.; Takahashi, M.; Hirose, N.; Hou, D.; Takara, K.; Wada, K. Wax, policosanol, and long-chain aldehydes of different sugarcane (Saccharum officinarum L.) cultivars. Eur. J. Lipid Sci. Tech. 2012, 114, 583–591. [Google Scholar] [CrossRef]
- Hansjakob, A.; Riederer, M.; Hildebrandt, U. Wax matters: Absence of very-long-chain aldehydes from the leaf cuticular wax of the glossy11 mutant of maize compromises the prepenetration processes of Blumeria graminis. Plant Pathol. 2011, 60, 1151–1161. [Google Scholar] [CrossRef]
- Tsuba, M.; Katagiri, C.; Takeuchi, Y.; Takada, Y.; Yamaoka, N. Chemical factors of the leaf surface involved in the morphogenesis of Blumeria graminis. Physiol. Mol. Plant Pathol. 2002, 60, 51–57. [Google Scholar] [CrossRef]
- Klose, J.; de Sá, M.M.; Kronstad, J.W. Lipid-induced filamentous growth in Ustilago maydis. Mol. Microbiol. 2004, 52, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Mendoza, A.; Berndt, P.; Djamei, A.; Weise, C.; Linne, U.; Marahiel, M.; Vranes, M.; Kämper, J.; Kahmann, R. Physical-chemical plant-derived signals induce differentiation in Ustilago maydis. Mol. Microbiol. 2009, 71, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Andrews, D.L.; Egan, J.D.; Mayorga, M.E.; Gold, S.E. The Ustilago maydis ubc4 and ubc5 genes encode members of a MAP kinase cascade required for filamentous growth. Mol. Plant-Microbe Interact. 2000, 13, 781–786. [Google Scholar] [CrossRef] [Green Version]
- Irie, K.; Takase, M.; Lee, K.S.; Levin, D.E.; Araki, H.; Matsumoto, K.; Oshima, Y. MKK1 and MKK2, which encode Saccharomyces cerevisiae mitogen-activated protein kinase-kinase homologs, function in the pathway mediated by protein kinase C. Mol. Cell. Biol. 1993, 13, 3076–3083. [Google Scholar]
- Mayorga, M.E.; Gold, S.E. Characterization and molecular genetic complementation of mutants affecting dimorphism in the fungus Ustilago maydis. Fungal Genet. Biol. 1998, 24, 364–376. [Google Scholar] [CrossRef]
- Lee, N.; Kronstad, J.W. ras2 Controls morphogenesis, pheromone response, and pathogenicity in the fungal pathogen Ustilago maydis. Eukaryot Cell 2002, 1, 954–966. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Z.; Zhang, B.; Chang, C.; Wang, Y.; Lu, S.; Sun, S.; Zhang, X.; Chen, B.; Jiang, Z. The MAP Kinase SsKpp2 Is Required for Mating/Filamentation in Sporisorium scitamineum. Front. Microbiol. 2018, 9, 2555. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, K.; Kusunose, E.; Noda, Y.; Kusunose, M. Some properties of the fatty alcohol oxidation system and reconstitution of microsomal oxidation activity in intestinal mucosa. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1986, 878, 412–418. [Google Scholar] [CrossRef]
- Jirausch, M.; Asperger, O.; Kleber, H.P. Alcohol oxidation by Acinetobacter calcoaceticus EB 104—An-alkane-utilizing and cytochrome P-450-producing strain. J. Basic Microbiol. 1986, 26, 351–357. [Google Scholar] [CrossRef]
- Nagashima, H.; Inoue, J.; Sasaki, E.; Yamamoto, S.; Sasaki, Y.; Yamauchi-Inomata, Y.; Harayama, S. Long-chain n-alkanol dehydrogenase from Pseudomonas putida. J. Ferment. Bioeng. 1996, 82, 328–333. [Google Scholar] [CrossRef]
- Lei, W.; Liu, X.; Lu, F. Thermophilic Long Chain Alkanol Dehydrogenase and Its Encoding Gene and Uses Thereof. Available online: https://www.freepatentsonline.com/wo2009030074.html (accessed on 10 December 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard | Molecular Weight (g/mol) | RT Time (min) |
|---|---|---|
| Palmitic Acid (C16H32O2) | 256.42 | 14.916 |
| Stearic Acid (C18H36O2) | 284.50 | 16.418 |
| Pentacosane (C25H32) | 352.70 | 18.332 |
| Heptacosane (C27H56) | 380.70 | 19.795 |
| Nonacosane (C29H60) | 408.80 | 21.079 |
| Tetracosanol (C24H50O) | 354.70 | 20.286 |
| Hexacosanol (C26H54O) | 382.70 | 21.493 |
| Octacosanol (C28H58O) | 410.80 | 23.036 |
| Triacontanol (C30H62O) | 438.80 | 24.899 |
| Octacosanal (C28H56O) | 408.70 | 22.164 |
| Clone | Smut Incidence Rate (%) |
|---|---|
| 3–33 | 68.08 |
| 3–69 | 58.22 |
| 23–15 | 62.80 |
| 26–122 | 60.60 |
| 25–113 | 48.59 |
| 43–51 | 0 |
| 45–23 | 0 |
| 45–53 | 0 |
| 46–33 | 0 |
| 79–40 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Lan, X.; Li, X.; Zhao, H.; Gan, J.; Li, R.; Chen, B. A Plant-Derived Alkanol Induces Teliospore Germination in Sporisorium scitamineum. J. Fungi 2022, 8, 209. https://doi.org/10.3390/jof8020209
Liu Z, Lan X, Li X, Zhao H, Gan J, Li R, Chen B. A Plant-Derived Alkanol Induces Teliospore Germination in Sporisorium scitamineum. Journal of Fungi. 2022; 8(2):209. https://doi.org/10.3390/jof8020209
Chicago/Turabian StyleLiu, Zongling, Xianruan Lan, Xiufang Li, Haiyun Zhao, Jiaming Gan, Ru Li, and Baoshan Chen. 2022. "A Plant-Derived Alkanol Induces Teliospore Germination in Sporisorium scitamineum" Journal of Fungi 8, no. 2: 209. https://doi.org/10.3390/jof8020209