
The Multifaceted Roles of Fungal Cutinases during Infection

Department of Vegetable and Field Crops, Institute of Plant Sciences, Agricultural Research Organization (ARO), Volcani Center, Rishon Lezion 7505101, Israel
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(2), 199; https://doi.org/10.3390/jof8020199
Submission received: 23 December 2021
/
Revised: 13 February 2022
/
Accepted: 13 February 2022
/
Published: 18 February 2022
(This article belongs to the Special Issue Plant and Fungal Interactions)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Cuticles cover the aerial epidermis cells of terrestrial plants and thus represent the first line of defence against invading pathogens, which must overcome this hydrophobic barrier to colonise the inner cells of the host plant. The cuticle is largely built from the cutin polymer, which consists of C16 and C18 fatty acids attached to a glycerol backbone that are further modified with terminal and mid-chain hydroxyl, epoxy, and carboxy groups, all cross-linked by ester bonds. To breach the cuticle barrier, pathogenic fungal species employ cutinases—extracellular secreted enzymes with the capacity to hydrolyse the ester linkages between cutin monomers. Herein, we explore the multifaceted roles that fungal cutinases play during the major four stages of infection: (i) spore landing and adhesion to the host plant cuticle; (ii) spore germination on the host plant cuticle; (iii) spore germ tube elongation and the formation of penetrating structures; and (iv) penetration of the host plant cuticle and inner tissue colonisation. Using previous evidence from the literature and a comprehensive molecular phylogenetic tree of cutinases, we discuss the notion whether the lifestyle of a given fungal species can predict the activity nature of its cutinases.
1. Introduction
The terrestrialisation of plants is considered to be a landmark event in plant evolution [1,2]. Key to this major step was the assembly of the cuticle, a lipophilic barrier that covers all plant aerial tissues, including the leaves, stems, flowers, and fruit [3]. The cuticle is made primarily from the polyester cutin, which consists of C16 and C18 fatty acids attached to a glycerol backbone that are further modified with terminal and mid-chain hydroxyl, epoxy, and carboxy groups, all cross-linked by ester bonds [4]. The cutin matrix is immersed in and coated with solvent-extractable cuticular waxes composed of various C20-C40 very-long-chain fatty acids (VLCFAs) as well as their esters and derivatives [5]. Besides fulfilling pivotal roles in the regulation of transpiration and the diffusion of gases, water, and solutes [6,7,8], the cuticle is also the first line of defence against a wide range of pathogens as it covers the outermost epidermal layer of aerial organs. Pathogens must penetrate the cuticle to colonise the nutritional inner cells of the host plant: some gain entry through stomatal pores, natural cracks, or wounds, whereas many others utilise an arsenal of strategies to breach the cuticle either by using mechanical pressure to pierce its surface or by degrading cutin using specialised hydrolytic enzymes known as cutinases. Although the cuticle covers all above ground organs, the tissues that built the outer bark of trees is made of a hetero-polymer called suberin, which is more complex than cutin as it is built from longer-chain fatty acids and aromatic p-hydroxycinnamic acids derived from the core phenylpropanoid pathway. It is assumed that pathogenic fungal species that are capable of penetrating these types of tissues possess unique enzymes called suberinases that are capable of degrading the suberin polymer. Nonetheless, research on suberinases is scarce with a very limited knowledge regarding the mechanisms allowing these enzymes to degrade suberin.
Cutinases [EC 3.1.1.74] are serine esterases belonging to the α/β hydrolase family that can break down cutin polyesters by hydrolysing the ester linkages between monomers [9,10]. The active site of cutinases consists of a catalytic triad of Ser, Asp and His, a sequence order that also corresponds to other types of lipase enzymes (Figure 1A) [11]. This triad is conserved among cutinases and can be found in the protein sequences of cutinases belonging to multiple fungal species with different lifestyles, including necrotrophic (e.g., Botrytis cinerea, Sclerotinia sclerotiorum and Verticilium dahliae), hemibiotrophic (e.g., Fusarium solani, Magnaporthe grisea and Colletotrichum gloeosporioides), biotrophic (e.g., Erysiphe necator and Blumeria graminis), and saprotrophic (e.g., Apergillus flavus, Aspergillus nidulans and Aspergillus oryzae) (Figure 1B). Despite these sequence similarities and conserved motifs, cutinases from fungi with different lifestyles exhibit dissimilar structural and biochemical attributes, including molecular weight, optimum temperature, optimal pH, isoelectric point, kinetic constants, substrate specificity, and thermostability (for more information see [12]). Pioneering work performed in the early 1960s and 1970s on fungal cutinases [13,14,15,16] paved the way for the characterisation of functional cutinases from diverse fungal species with different lifestyles. It has now been established that the progression of above-ground fungal infections relies closely on cutinase activity, which often correlates with phytopathogenicity in plant-associated pathogenic fungal species and is typically not detected in fungi that are not associated with plant hosts [17]. Recent reviews have covered the interactions between plant cuticles and pathogens [8,18,19,20,21] with a focus on the mechanisms via which cuticle-derived components act as chemical signalling molecules to elicit host plant defence responses and how cuticle imperfections affect the nature of plant-pathogen interactions. However, the role of cutinases as the driving force that helps pathogens to breach this barrier have often been overlooked, despite its key role during pathogenic attack.
In this review, we provide a synopsis of the multifaceted roles of fungal cutinases during the main four stages of infection: (i) spore landing and adhesion to the host plant cuticle; (ii) spore germination on the host plant cuticle; (iii) spore germ tube elongation and the formation of penetrating structures; and (iv) penetration of the host plant cuticle and inner tissue colonisation. We then shift our focus to the concept whether the lifestyle of a given fungi is associated with cutinase activity using previous evidence from the literature and a comprehensive molecular phylogeny tree that we have constructed herein.
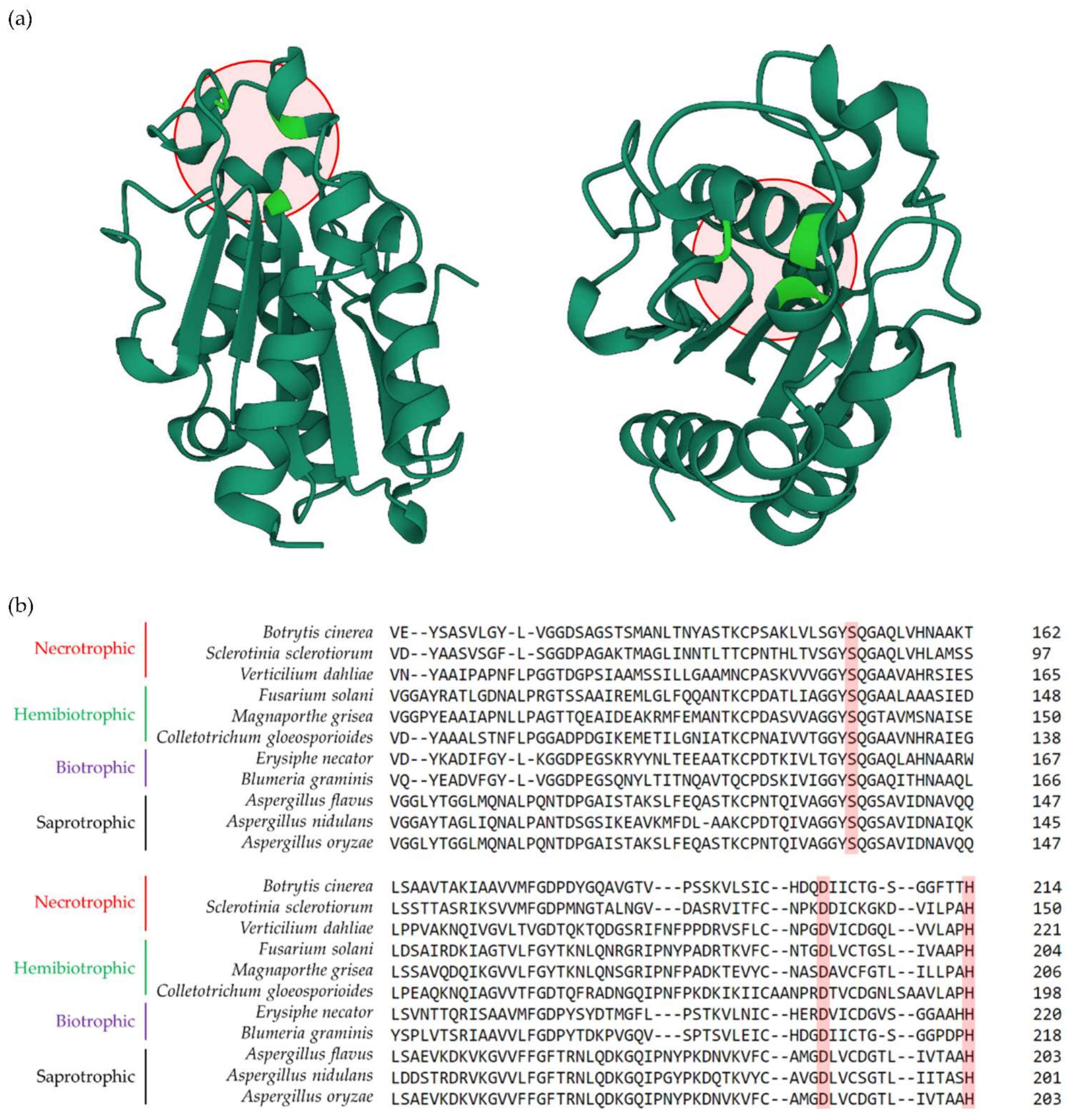
Figure 1.
Structure of the Fusarium solani cutinase and the conserved catalytic triad among fungal species with different lifestyles. (a) The crystal structure of the Fusarium solani CUT enzyme (protein ID: 1CEX) according to the RCSB Protein Data Bank (PDB) (https://www.rcsb.org/ accessed on 20 December 2021). Original structure was elucidated based on X-ray crystallography analyses at atomic resolution of 1.0A° performed by [22]. The structure can be seen in a front (left) and upright (right) overviews. Red circles show the catalytic triad consists of Ser, Asp and His, that are marked in green. (b) Protein alignment of cutinases isolated from fungal species with different lifestyles, including necrotrophic (e.g., Botrytis cinerea, Sclerotinia sclerotiorum and Verticilium dahliae), hemibiotrophic (e.g., Fusarium solani, Magnaporthe grisea and Colletotrichum gloeosporioides), biotrophic (e.g., Erysiphe necator and Blumeria graminis), and saprotrophic (e.g., Apergillus flavus, Aspergillus nidulans and Aspergillus oryzae). The conserved amino acids of Ser, Asp and His that build the catalytic triad are highlighted in red rectangles. Protein sequence alignment was performed using the tools embedded in the ClustalW2 software (https://www.ebi.ac.uk/Tools/msa/clustalw2/ accessed on 12 February 2022).
Figure 1.
Structure of the Fusarium solani cutinase and the conserved catalytic triad among fungal species with different lifestyles. (a) The crystal structure of the Fusarium solani CUT enzyme (protein ID: 1CEX) according to the RCSB Protein Data Bank (PDB) (https://www.rcsb.org/ accessed on 20 December 2021). Original structure was elucidated based on X-ray crystallography analyses at atomic resolution of 1.0A° performed by [22]. The structure can be seen in a front (left) and upright (right) overviews. Red circles show the catalytic triad consists of Ser, Asp and His, that are marked in green. (b) Protein alignment of cutinases isolated from fungal species with different lifestyles, including necrotrophic (e.g., Botrytis cinerea, Sclerotinia sclerotiorum and Verticilium dahliae), hemibiotrophic (e.g., Fusarium solani, Magnaporthe grisea and Colletotrichum gloeosporioides), biotrophic (e.g., Erysiphe necator and Blumeria graminis), and saprotrophic (e.g., Apergillus flavus, Aspergillus nidulans and Aspergillus oryzae). The conserved amino acids of Ser, Asp and His that build the catalytic triad are highlighted in red rectangles. Protein sequence alignment was performed using the tools embedded in the ClustalW2 software (https://www.ebi.ac.uk/Tools/msa/clustalw2/ accessed on 12 February 2022).

2. Fungal Cutinases Play Multifaceted Roles during Different Stages of Infection
2.1. Spore Landing and Adhesion to the Host Plant Cuticle
Spores are microscopic dispersal units produced by fungi through sexual or asexual reproduction that allow them to survive harsh conditions and reproduce. Fungal spores can be carried to the surface of the host plant by wind, water, or animal vectors. During the initial stages of infection, spores sense that they have landed on the host plant surface and remain dormant until they sense suitable conditions for infection [23] (Figure 2). These conditions vary and depend on multiple factors, including nutrient and water availability and the surface biochemistry of the host plant [24]. This initial passive, non-metabolic spore adhesion to the surface of the cuticle then allows active spore adhesion during the advanced stages of infection [25]. Fungal spore adhesion to the host plant surface involves dedicated binding and recognition components along with the secretion of adhesive compounds, such as insoluble proteins, lipids and polysaccharides [26]. These compounds, which already exist in dormant spores, are rapidly secreted towards the surface upon engagement (Figure 2) [27]. Therefore, successful and complete spore adhesion is crucial for later stages of infection and cuticle penetration, and significantly affects overall disease development [28].
The dormant spores of pathogenic fungi contain ‘constitutive-type’ cutinases, also termed previously as “sensing” cutinases, release small amounts of cutin monomers from the host plant cuticle in a spatially-localised manner (Figure 2) [29]. The molecular basis of this type of interaction has been studied in the promoter of the Fusarium solani f.sp. pisi cutinase gene using in vitro and in vivo methodologies. These efforts have identified novel regulatory elements and transcription factors that are involved in cutinase induction, including a silencing sequence that keeps basal gene expression low and affects cutinase gene inducibility; a G-rich positive-acting Sp1-like element that restores high expression levels by antagonising the silencer; a GC-rich palindrome that is vital for cutinase induction by cutin monomers; and two cutinase transcription factors (CTF1 and CTF2) that act as transcriptional inducers by binding to palindromic regions in the cutinase promoter [30,31,32,33,34,35,36]. Constitutive-type cutinase activity has been detected during early stages of infection in the dormant spores of fungal species with different infection strategies such as Botrytis cinerea, Fusarium graminearium, Curvularia lunata, Pyrenopeziza brassicae, Magnaporthe grisea and Colletotrichum spp. [36,37,38,39,40,41,42,43,44]. In the broad bean rust-causing biotrophic fungus Uromyces viciae-fabae, cutinases and other serine esterases were detected on the surface of adhesion pads formed by its spores on the bean leaves. The capacity of these spores to adhere to the host plant surface was significantly reduced when these enzymes were rinsed from the pad surface or chemically inhibited, but restored by exogenous cutinase application [45]. Thus, constitutive-type cutinases play key roles by altering the host plant surface to allow successful spore adhesion and by releasing cutin monomers from the cuticle layer that are essential for subsequent stages of infection.
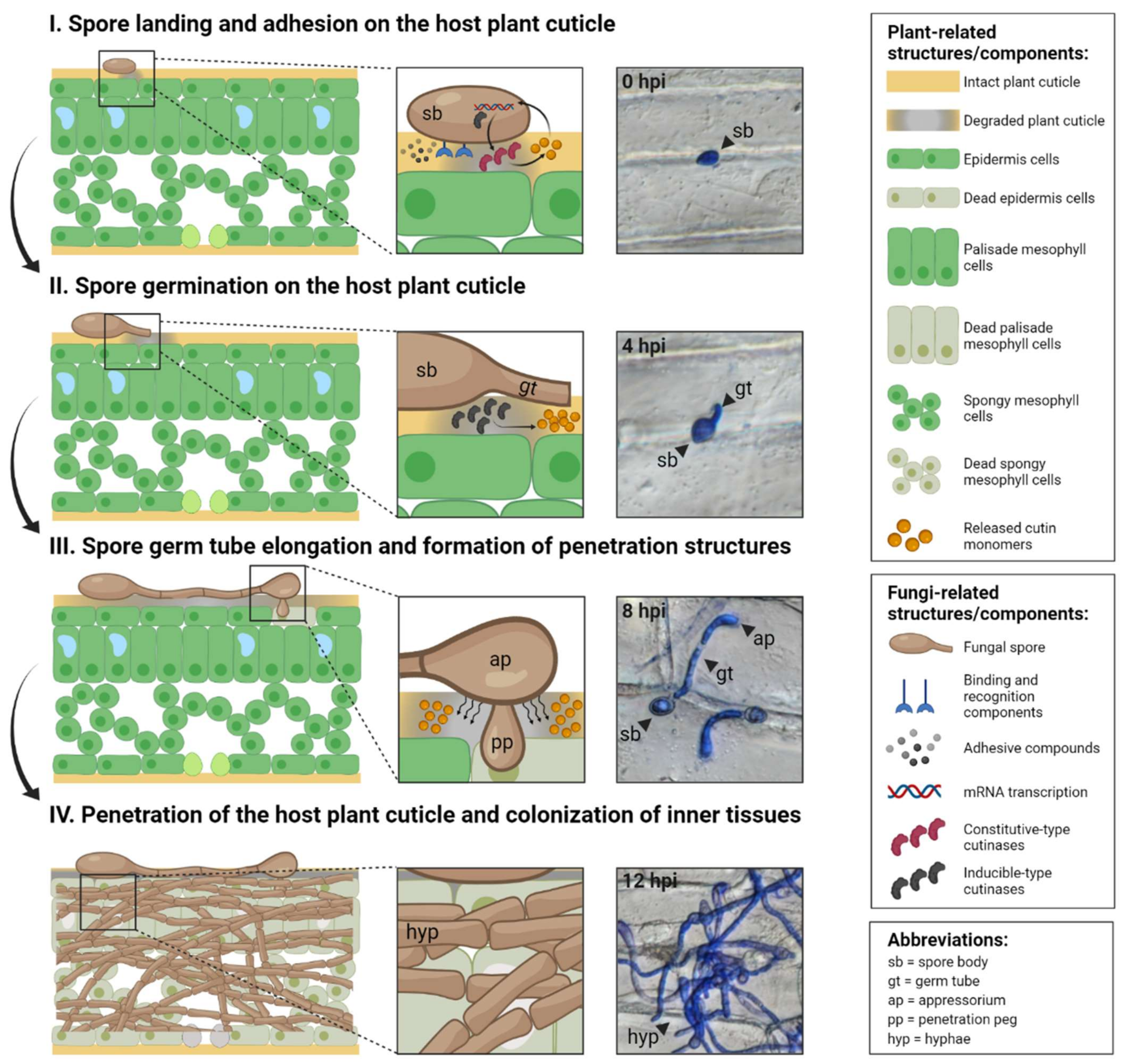
Figure 2.
Role of fungal cutinases in the four fundamental stages of infection. Left panels: key processes in each stage. Middle panels: enlarged regions of interaction between the host plant cuticle and fungi. Right panels: micrographs of the necrotrophic fungus Botrytis cinerea infecting onion (Allium cepa) peel tissues at each stage. Right boxes: explanations of plant- and fungi-related structures/components and abbreviations. Stage (I) spores land and adhere to the host plant surface by secreting adhesive compounds and through dedicated binding and recognition components. ‘Constitutive-type’ cutinases actively release small amounts of cutin monomers from the host plant cuticle in a spatially-localised manner. Stage (II) spores hydrate by absorbing water and nutrients from the plant surface and enter a polar growth stage that leads to the formation of elongated germ tubes on the cuticle surface in preparation for penetration. Previously released cutin monomers are recognised by the spore surface, leading to the production of ‘inducible-type’ cutinases. Stage (III) germ tubes continue outgrowth. In many pathogenic fungi, the germ tube tip forms a swollen structure (appressorium) that facilitates cuticle penetration by applying mechanical force to the surface and piercing the cuticle via a penetration peg. Stage (IV) elongated germ tubes produce extended cylindrical cells (hyphae) that are typically long, filamentous and serve as the main mode of vegetative growth and/or an invasive organ that expedites cuticle penetration. The roles of fungal cutinases in cuticle penetration at these stages are demonstrated in various fungi species with different lifestyles. Figure created using BioRender (https://biorender.com/ accessed on 20 December 2021).
Figure 2.
Role of fungal cutinases in the four fundamental stages of infection. Left panels: key processes in each stage. Middle panels: enlarged regions of interaction between the host plant cuticle and fungi. Right panels: micrographs of the necrotrophic fungus Botrytis cinerea infecting onion (Allium cepa) peel tissues at each stage. Right boxes: explanations of plant- and fungi-related structures/components and abbreviations. Stage (I) spores land and adhere to the host plant surface by secreting adhesive compounds and through dedicated binding and recognition components. ‘Constitutive-type’ cutinases actively release small amounts of cutin monomers from the host plant cuticle in a spatially-localised manner. Stage (II) spores hydrate by absorbing water and nutrients from the plant surface and enter a polar growth stage that leads to the formation of elongated germ tubes on the cuticle surface in preparation for penetration. Previously released cutin monomers are recognised by the spore surface, leading to the production of ‘inducible-type’ cutinases. Stage (III) germ tubes continue outgrowth. In many pathogenic fungi, the germ tube tip forms a swollen structure (appressorium) that facilitates cuticle penetration by applying mechanical force to the surface and piercing the cuticle via a penetration peg. Stage (IV) elongated germ tubes produce extended cylindrical cells (hyphae) that are typically long, filamentous and serve as the main mode of vegetative growth and/or an invasive organ that expedites cuticle penetration. The roles of fungal cutinases in cuticle penetration at these stages are demonstrated in various fungi species with different lifestyles. Figure created using BioRender (https://biorender.com/ accessed on 20 December 2021).

2.2. Spore Germination on the Host Plant Cuticle
Prolonged adhesion promotes tighter and steadier attachment between the plant cuticle and spores as they secrete a polysaccharide-based extracellular mucilaginous matrix towards the plant surface. During this process, spores absorb water and nutrients from the plant surface, resulting in rapid swelling and increased volume as they become hydrated (Figure 2) [46]. Omics-based studies have indicated that the secreted extracellular matrix contains a mixture of hydrolytic enzymes, including pectinases, cellulases, and cutinases. Indeed, cutinases have been detected in the adherent spores of many fungal species, such as Uromyces viciae-fabae [45], Erysiphe graminis f. sp. hordei [47], Colletotrichum gramminicola [48], Botrytis cinerea [49,50], and Fusarium solani [29].
During this infection stage, the cutin monomers released earlier by constitutive-type cutinases are recognised by the spore surface, resulting in the protracted production of ‘inducible-type’ cutinases within minutes, which is the core driving force of cuticle penetration during later stages of infection (Figure 2) [51,52,53,54]. We have recently reviewed the signalling roles played by the released cutin monomers as well as epicuticular waxes deposited on top of the cuticle surface, during the progression of infection [18]. Consistently, various in vitro studies have shown that cutinases from different pathogenic fungi respond positively to the exogenous application of naturally extracted or chemically synthesised cutin monomers to their enzymatic buffers [16,54,55,56]. For instance, the expression of Fusarium solani cutinases was induced by supplementation with cutin n-aliphatic C16 and C18 primary alcohols [30,51], while cutinases from Botrytis cinerea, Monilinia fructicola, Pyrenopeziza brassicae, Phytopthera spp., Ascochyta rabiei, Sclerotinia sclerotiorum and Alternaria brassicicola were induced by the predominant cutin monomer, 16-hydroxyhexadecanoic acid [40,50,57,58,59,60]. Following complete adhesion, spores enter a polar growth phase that leads to the formation of elongated tube-like structures, known as spore germ tubes, accompanied by fundamental morpho- and cytokinetic cellular processes [46,61]. At this stage, the germ tubes lie on the surface of the cuticle in preparation for penetration. Hence, an increased production of inducible-type cutinases at this stage of infection represents the elementary phase of cuticle degradation that would be accelerated at later stages of infection.
2.3. Spore Germ Tube Elongation and the Formation of Penetrating Structures
As infection progresses, spore germ tubes continue their outgrowth and, in many pathogenic fungal species, their tips transform into a swollen structure known as the appressorium. This structure facilitates cuticle penetration at later stages of infection by applying a mechanical force towards the surface and piercing the cuticle using a penetration peg [26] (Figure 2). These two structures are considered to be the most specialised penetrating structures that breach the cuticle barrier, allowing the fungus to reach the inner tissue of the host plant.
Studies have shown that the cutin monomers released by both types of cutinases during earlier stages of infection promote germ tube assembly and appressoria differentiation in Erysiphe graminis f. sp hordei and Blumeria graminis [52,62] and Magnaporthe grisea [53,63]. In the latter species, upregulated expression of the cutinase gene, MgCUT2, was detected during appressorium maturation, while lower cutinase activity was measured in a cut2 mutant strain that failed to form a structurally-active appressorium [42]. Similarly, appressorium disruption was detected in Venturia ineaequalis when cutinases were chemically-inhibited [64]. Together, these findings suggest that cutinases operate in a localised manner in germ tubes and appressoria, emphasising their central roles prior to the final penetration of the host plant cuticle barrier and the colonisation of inner tissues.
2.4. Penetration of the Host Plant Cuticle and Inner Tissue Colonisation
Throughout most of their life cycle, the majority of pathogenic fungi exist as extended cylindrical cells that arise from elongated spore germ tubes, known as hyphae (Figure 2). These cells are typically long and filamentous, and are the main mode of fungal vegetative growth; however, in some species hyphae act as an invasive organ that accelerates the penetration of the host plant cuticle. The role of fungal cutinases in host plant cuticle penetration has been inferred from studies showing that cutinase activity is highest in the tips of germ tubes and appressoria penetration pads, which serve as penetrating structures [65]. These crucial effects of cutinases during the final stage of infection have been demonstrated using a wide range of approaches, including reducing cutinase activity using synthetic chemical inhibitors, lowering its expression by mutation or genetic manipulation, and increasing its activity using ectopic integration or overexpression. For example, the chemical inhibition or mutation of cutinases reduced infection rates and cuticle penetration capacity in species belonging to the necrotrophic fungi Venturia, Verticillium, Alternaria, and Curvularia [39,62,66,67,68]; the hemibiotrophic Fusarium, Pyrenopeziza, Magnaporthe, and Colletotrichum [29,40,42,43,49,69,70,71,72,73]; and the biotrophic Erysiphe and Uromyces [45,52]. Although spores from some of these mutants were able to germinate on the cuticle surface, they were unable to penetrate the cuticle and their hyphal tips were augmented and abnormal, indicating ineffective penetration attempts. Remarkably, these mutants easily penetrated the cuticle when placed on wounded surfaces, further emphasising the importance of cutinase activity for this process. Consistently, studies have also shown that cutinase overexpression results in heightened and more efficient pathogenicity. For example, the overexpression of cutinase MfCUT1 from Monilinia fructicola increased brown rot lesion sizes in peaches, nectarines, and cherry flower petals, and was generally more virulent than the wild-type strain [74]. Another study examined Mycosphaerella spp., a fungus that is naturally unable to penetrate the cuticle of papaya fruit and instead infects through wounds. Interestingly, introducing Fusarium cutinase into Mycosphaerella yielded transformants that could successfully infect intact papaya fruit and penetrate their cuticle [75]. Overall, penetration through the host plant cuticle occurs directly or via the assistance of penetrating structures or both and is mediated by and tightly relies on cutinase activity. Following effective penetration, ramified invasive hyphae are established over the inner cells of the host plant.
3. The Association between Lifestyles of Fungi and Cutinase Activity
Cutinases have been identified in multiple fungal species with different lifestyles. Necrotrophic fungal species actively kill host plant cells and feed on their dead tissues. These species are mostly non-obligate, have a wide range of hosts, and are known to secrete copious amounts of cell-wall degrading enzymes as well as toxins [41,44]. Hemibiotrophic fungal species initiate biotrophic interactions with host plant cells followed by a necrotrophic interaction that kills the cells to feed on their dead tissues 40,43]. Moreover, it has been reported that necrotrophic and hemibiotrophic fungal species possess a cryptic biotrophic phase allowing them to persist in intercellular spaces without causing disease symptoms [76,77,78]. Biotrophic fungal species colonise living host plant cells without killing them and causing relatively little damage. These species can be obligate or facultative, typically have a narrow host range, and secrete limited amounts of cell-wall degrading enzymes [41,44]. Saprotrophic fungal species also do not kill the host plant as they can feed from entirely dead plant tissues [79]. Even though the latter two lifestyles share a similar pattern of not taking part in killing the plant cells during infection, the distinct phylogenetic separation between them is fairly predictable as biotrophic fungi still need to penetrate the cuticle, while saprotrophic fungi encounter a plant tissue that is already degraded and decayed, in which the cuticle does not present any sort of a barrier for penetration. The existence of cutinase encoding-genes in saprotrophic fungal genomes, on the other hand, plainly indicates that, although surviving on dead and decaying plant materials, they still require cutinase activity to some degree to further catalyse the complex cuticle existing in these tissues. Although each of the abovementioned lifestyles of fungi are considered distinct and are used to define a fungal species, there is a wide continuum between saprotrophy and parasitism, as well as between each of these lifestyles.
Previous studies have suggested that plant-pathogenic fungi contain a conserved set of cutinases irrespective of their lifestyle [39,59,80]. In order to further investigate whether there is an association between the origin and lifestyle of plant-pathogenic fungi and cutinase type and/or activity, we generated an inclusive protein maximum-likelihood phylogenetic tree encompassing 68 functionally characterized and/or putative cutinases isolated from fungi with different lifestyles. We employed the Whelan and Goldman + Frequencies + Gamma distribution model (i.e., WAG + F + G), as suggested by a model-testing step performed prior the generation of phylogenetic tree. Finally, two bacterial cutinases and one plant-type cutinase were included in the tree as reference outgroups allowing the outcome of reliable evolutionary relationships between fungal cutinases (Figure 3).
By and large, the resulting molecular phylogenetic tree inferred relatively distinct separations of cutinases originated from saprophytic and biotrophic fungal species. For instance, all nine cutinases belonging to the saprophytic Aspergillus genus (comprising of A. nidulans, A. flavus, and A. oryzae) were grouped together forming a distinct sub-clade (Figure 3). This may suggest that fungal species belonging to this genus have similar types of cutinases that are apparently different from cutinases belonging to fungal species with different lifestyles. A similar trend was detected in the three cutinases isolated from the biotrophic fungal species of Blumeria graminis and Erysiphe necator, that formed a separate sub-clade (Figure 3). Supporting these separation patterns are the high percentages of trees in which these associated fungal species clustered together following bootstrapping (Figure 3). We fully aware that our phylogenetic tree contains a relatively low number of cutinases originating from saprophytic and biotrophic fungal species, as these were the only cutinases obtained following blast assays against the well-characterized and structurally-resolved Fusarium solani cutinase (>30% protein sequence identity). Alternatively, our blast assays yielded many cutinases from necrotrophic and hemibiotrophic fungal species belonging to various genera (Figure 3). Some of these were divided polyphyletically, meaning that different cutinases from the same genus/species were found in different sub-clades. Examples include nine isolated cutinases of the hemibiotrophic fungal species Magnaporthe grisea found in four distinct sub-clades; six cutinases of the necrotrophic fungal species Alternaria alternata found in four sub-clades; and eight cutinases of the necrotrophic fungal species Verticillium dahliae found in three sub-clades (Figure 3). On the other hand, all 10 cutinases of the necrotrophic fungal species Botrytis cinerea and Sclerotinia sclerotiorum were grouped at the same sub-clade, implying that cutinases from these two species are relatively of a similar type (Figure 3).
We also classified each of the fungal species incorporated in the molecular phylogenetic tree to different classes of the Ascomycota phylum including Sordariomycetes, Dothideomycetes, Leotiomycetes and Eurotiomycetes, all of which are evolutionarily-distinct based on their ascomatal (i.e., fruiting body) characters [81]. The saprotrophic Aspergillus spp. included in the phylogenetic tree all belong to the Eurotiomycetes class, and therefore may suggest that species belonging to this specific class may have a single type of a cutinase. The other three classes, Sordariomycetes, Dothideomycetes and Leotiomycetes, span throughout different clades in the phylogenetic tree, inferring that they include species with numerous phylogenetically distinct types of cutinases (Figure 3). Taken together, it appears that necrotrophic and hemibiotrophic fungal species have multiple types of cutinases, while biotrophic and saprotrophic fungal species seem to have one type of cutinase. This is an important piece of evidence that requires further attention. Phylogenetic incorporation of additional putative cutinases from fungal species with different lifestyles and Ascomycota classes will likely provide greater insight towards these remarkable concepts.
Figure 3.
Molecular phylogenetic tree of cutinases of fungal species with different lifestyles. The phylogenetic tree was inferred by using the Maximum Likelihood method and Whelan and Goldman + Freq. model [82]. Gamma distribution was used to model evolutionary rate differences among sites [five categories (+G, parameter = 1.1745)]. The tree with the highest log likelihood (−26,692.25) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches following 1000 bootstrap replications. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. There was a total of 612 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [83]. The analysis involved the full protein sequences of 68 characterised and/or putative cutinases belonging to fungal species with different lifestyles based on blast analysis against the well-characterised and structurally resolved Fusarium solani cutinase (AAL18697). All of these proteins exhibited >30% sequence identity vs. the Fusarium solani cutinase. The isolated proteins belong to 32 necrotrophic (●), 24 hemibiotrophic (●), three biotrophic (●), and nine saprotrophic (●) fungal species. In addition, cutinases from each fungal species are classified to the following Ascomycota classes: Sordariomycetes (∆), Dothideomycetes (◻), Leotiomycetes (♢), and Eurotiomycetes (ο). For outgroups, we have used the full protein sequences of cutinases isolated from Arabidopsis thaliana, Streptomyces scabiei and Mycobacterium spp. Protein IDs are given in parenthesis following the species name.
Figure 3.
Molecular phylogenetic tree of cutinases of fungal species with different lifestyles. The phylogenetic tree was inferred by using the Maximum Likelihood method and Whelan and Goldman + Freq. model [82]. Gamma distribution was used to model evolutionary rate differences among sites [five categories (+G, parameter = 1.1745)]. The tree with the highest log likelihood (−26,692.25) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches following 1000 bootstrap replications. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. There was a total of 612 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [83]. The analysis involved the full protein sequences of 68 characterised and/or putative cutinases belonging to fungal species with different lifestyles based on blast analysis against the well-characterised and structurally resolved Fusarium solani cutinase (AAL18697). All of these proteins exhibited >30% sequence identity vs. the Fusarium solani cutinase. The isolated proteins belong to 32 necrotrophic (●), 24 hemibiotrophic (●), three biotrophic (●), and nine saprotrophic (●) fungal species. In addition, cutinases from each fungal species are classified to the following Ascomycota classes: Sordariomycetes (∆), Dothideomycetes (◻), Leotiomycetes (♢), and Eurotiomycetes (ο). For outgroups, we have used the full protein sequences of cutinases isolated from Arabidopsis thaliana, Streptomyces scabiei and Mycobacterium spp. Protein IDs are given in parenthesis following the species name.

4. Conclusions and Future Perspective
Plant–fungi interaction is a multidimensional process governed by the secretion of various components from both the host plant and the fungus towards the cuticle surface. In particular, fungal cutinases among other enzymes are major determinants of pathogenicity and affect the capacity of the fungus to penetrate the cuticle and colonise plant inner tissues. Pathogenic fungi have different lifestyles which might influence the nature of its associated cutinases, however, this notion is poorly understood and requires further attention. As discussed above, the differences among these cutinases might be attributed to the various mechanisms and dissimilar infection strategies employed by fungi. The expression of different isoenzymes of cutinases was detected during saprophytic and parasitic stages of Alternaria brassicicola [84]. This suggests that different temporal infection stages of the very same fungus may also involve unalike cutinases with different characteristics that are required to execute this specific infection stage. Supporting this notion is a study that performed phylogenetic analysis and in-depth characterization of functionally and structurally diverse cutinases, implying that the genome of a single fungus might possess genes encoding different cutinase with low sequence similarities and broad functionality [85]. One study demonstrated that the secretomes (that typically include cutinases) of fungi with similar lifestyles share common attributes, but the phylogenetic history of the fungi during evolution has greater impact on the secretome compared to its lifestyle adaptation [86]. Additional bioinformatics study performed a large-scale sequence-similarity network analysis to identify common components of virulence mechanisms of major pathogenic fungi lifestyles. This approach suggested that the exploitation of cutinases is a significantly enriched function of hemibiotrophic and necrotrophic lifestyles [87].
All in all, a more comprehensive knowledge on fungal cutinases’ activity and properties is crucial not only for understanding plant-fungi pathosystems, but also due to the emerging roles of cutinases as efficient biocatalysts with manifold of applications in industrial biotechnology. These applications range from the degradation of numerous plastics synthetic polymers, enzymatic de-polymerization in treatment of post-used synthetic polymers, to bioremediation of many other substances and pollutants [12].
Author Contributions
G.C.A. collected and analysed the data for reconstructing the cutinases’ protein alignments and molecular phylogenetic tree. G.C.A. performed inoculation experiments of the necrotrophic fungus Botrytis cinerea and onion (Allium cepa) peel tissues. G.C.A. and H.C. drafted and wrote the manuscript. Both authors have agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cohen, H.; Szymanski, J.; Aharoni, A. Assimilation of “omics” strategies to study the cuticle layer and suberin lamellae in plants. J. Exp. Bot. 2017, 68, 5389–5400. [Google Scholar] [CrossRef] [PubMed]
- Philippe, G.; Geneix, N.; Petit, J.; Guillon, F.; Sandt, C.; Rothan, C.; Lahaye, M.; Marion, D.; Bakan, B. Assembly of tomato fruit cuticles: A cross-talk between the cutin polyester and cell wall polysaccharides. New Phytol. 2020, 226, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Cobb, E.D.; Matas, A.J. The evolution of hydrophobic cell wall biopolymers: From algae to angiosperms. J. Exp. Bot. 2017, 68, 5261–5269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Komatsuda, T.; Ma, J.F.; Li, C.; Yamaji, N.; Nevo, E. A functional cutin matrix is required for plant protection against water loss. Plant Signal. Behav. 2011, 6, 1297–1299. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, T.; Kosma, D.K.; Matas, A.J.; Buda, G.J.; He, Y.; Yu, B.; Pravitasari, A.; Batteas, J.D.; Stark, R.E.; Jenks, M.A.; et al. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 60, 363–377. [Google Scholar] [CrossRef]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional roles of plant cuticle during plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Enzymatic penetration of the plant cuticle by fungal pathogens. Annu. Rev. Phytopathol. 1985, 23, 223–250. [Google Scholar] [CrossRef]
- Longhi, S.; Cambillau, C. Structure-activity of cutinase, a small lipolytic enzyme. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids. 1999, 1441, 185–196. [Google Scholar] [CrossRef]
- Martinez, C.; de Geus, P.; Lauwereys, M.; Matthyssens, G.; Cambillau, C. Fusarium solani cutinase is a lipolytic enzyme with a catalytic serine accessible to solvent. Nature 1992, 356, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Nikolaivits, E.; Kanelli, M.; Dimarogona, M.; Topakas, E. A middle-aged enzyme still in its prime: Recent advances in the field of cutinases. Catalysts 2018, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Heinen, W. Über Den Enzymatischen Cutin-Abbau v. Mitteilung: Die Lyse von Peroxyd-Brücken Im cutin durch eine peroxydase aus Penicillium spinulosum Thom. Acta Bot. Neerl. 1963, 12, 51–57. [Google Scholar] [CrossRef]
- Shishiyama, J.; Araki, F.; Akai, S. Studies on cutin-esterase II. characteristics of cutin-esterase from Botrytis cinerea and its activity on tomato-cutin. Plant Cell Physiol. 1970, 11, 937–947. [Google Scholar] [CrossRef]
- Purdy, R.E.; Kolattukudy, P.E. Depolymerization isolation of a hydroxy enzyme some and fatty from acid Fusarium solani f. sp. pisi. isolation and some properties of the enzyme. Arch. Biochem. Biophys. 1973, 159, 61–69. [Google Scholar] [CrossRef]
- Purdy, R.E.; Kolattukudy, P.E. Hydrolysis of plant cuticle by plant pathogens. properties of cutinase i, cutinase ii, and a nonspecific esterase isolated from Fusarium solani pisi. Biochemistry 1975, 14, 2832–2840. [Google Scholar] [CrossRef]
- Morid, B.; Zare, R.; Rezaee, S.; Zamani-Zadeh, H.; Hajmansour, S. The relationship between cutinases and the pathogenicity/virulence of Fusarium solani in potato tubers. Phytopathol. Mediterr. 2009, 48, 403–410. [Google Scholar] [CrossRef]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The plant cuticle: An ancient guardian barrier set against long-standing rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef]
- Aragón, W.; Reina-Pinto, J.J.; Serrano, M. The intimate talk between plants and microorganisms at the leaf surface. J. Exp. Bot. 2017, 68, 5339–5350. [Google Scholar] [CrossRef]
- Fich, E.A.; Segerson, N.A.; Rose, J.K.C. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Métraux, J.P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhi, S.; Czjzek, M.; Lamzin, V.; Nicolas, A.; Cambillau, C. Atomic resolution (1.0 Å) crystal structure of Fusarium solani cutinase: Stereochemical analysis. J. Mol. Biol. 1997, 268, 779–799. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.J.; Howard, R.J. Adhesion of fungal spores and germlings to host plant surfaces. Protoplasma 1994, 181, 202–212. [Google Scholar] [CrossRef]
- Osherov, N.; May, G.S. The molecular mechanisms of conidial germination. FEMS Microbiol. Lett. 2001, 199, 153–160. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef] [Green Version]
- Tucker, S.L.; Talbot, N.J. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu. Rev. Phytopathol. 2001, 39, 385–417. [Google Scholar] [CrossRef]
- Tudzynski, P.; Sharon, A. Fungal Pathogenicity Genes. In Applied Mycology and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2003; pp. 187–212. [Google Scholar]
- Nicholson, R.L.; Epstein, L. Adhesion of Fungi to the Plant Surface. In The Fungal Spore and Disease Initiation in Plants and Animals; Springer US: Boston, MA, USA, 1991; pp. 3–23. [Google Scholar]
- Köller, W.; Allan, C.R.; Kolattukudy, P.E. Protection of Pisum sativum from Fusarium solani f. sp. pisi by inhibition of cutinase with organophosphorus pesticides. Phytopathology 1982, 72, 1425–1430. [Google Scholar] [CrossRef]
- Lin, T.S.; Kolattukudy, P.E. Induction of a biopolyester hydrolase (cutinase) by low levels of cutin monomers in Fusarium solani f. sp. pisi. J. Bacteriol. 1978, 133, 942–951. [Google Scholar] [CrossRef] [Green Version]
- Soliday, C.L.; Dickman, M.B.; Kolattukudy, P.E. Structure of the cutinase gene and detection of promoter activity in the 5’-flanking region by fungal transformation. J. Bacteriol. 1989, 171, 1942–1951. [Google Scholar] [CrossRef] [Green Version]
- Bajar, A.; Podila, G.K.; Kolattukudy, P.E. Identification of a fungal cutinase promoter that is inducible by a plant signal via a phosphorylated trans-acting factor. Proc. Natl. Acad. Sci. USA 1991, 88, 8208–8212. [Google Scholar] [CrossRef] [Green Version]
- Kämper, J.T.; Kämper, U.; Rogers, L.M.; Kolattukudy, P.E. Identification of regulatory elements in the cutinase promoter from Fusarium solani f. sp. pisi (Nectria haematococca). J. Biol. Chem. 1994, 269, 9195–9204. [Google Scholar] [CrossRef]
- Kolattukudy, P.E.; Rogers, L.M.; Li, D.; Hwang, C.S.; Flaishman, M.A. Surface signaling in pathogenesis. Proc. Natl. Acad. Sci. USA 1995, 92, 4080–4087. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Kolattukudy, P.E. Cloning of cutinase transcription factor 1, a transactivating protein containing Cys6Zn2 binuclear cluster dna-binding motif. J. Biol. Chem. 1997, 272, 12462–12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Sirakova, T.; Rogers, L.; Ettinger, W.F.; Kolattukudy, P.E. Regulation of constitutively expressed and induced cutinase genes by different zinc finger transcription factors in Fusarium solani f. sp. pisi (Nectria Haematococca). J. Biol. Chem. 2002, 277, 7905–7912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindro, K.; Pezet, R. Evidence for a constitutive cytoplasmic cutinase in ungerminated conidia of Botrytis cinerea Pers.: Fr. FEMS Microbiol. Lett. 1997, 149, 89–92. [Google Scholar] [CrossRef]
- Leroch, M.; Kleber, A.; Silva, E.; Coenen, T.; Koppenhöfer, D.; Shmaryahu, A.; Valenzuela, P.D.T.; Hahn, M. Transcriptome profiling of Botrytis cinerea conidial germination reveals upregulation of infection-related genes during the prepenetration stage. Eukaryot. Cell 2013, 12, 614–626. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hou, J.; Wang, Y.; Jin, Y.; Borth, W.; Zhao, F.; Liu, Z.; Hu, J.; Zuo, Y. Genome-wide identification, classification and expression analysis in fungal–plant interactions of cutinase gene family and functional analysis of a putative ClCUT7 in Curvularia lunata. Mol. Genet. Genom. 2016, 291, 1105–1115. [Google Scholar] [CrossRef]
- Davies, K.A.A.; De Lorono, I.; Foster, S.J.J.; Li, D.; Johnstone, K.; Ashby, A.M.M. Evidence for a role of cutinase in pathogenicity of Pyrenopeziza brassicae on Brassicas. Physiol. Mol. Plant Pathol. 2000, 57, 63–75. [Google Scholar] [CrossRef]
- Oliver, R.P.; Ipcho, S.V.S. Arabidopsis pathology breathes new life into the necrotrophs-vs.-biotrophs classification of fungal pathogens. Mol. Plant Pathol. 2004, 5, 347–352. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [Green Version]
- Auyong, A.S.; Ford, R.; Taylor, P.W. The Role of Cutinase and its impact on pathogenicity of Colletotrichum truncatum. J. Plant Pathol. Microbiol. 2015, 06, 1000259. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Deising, H.; Nicholson, R.L.; Haug, M.; Howard, R.J.; Mendgen, K. Adhesion pad formation and the involvement of cutinase and esterases in the attachment of uredospores to the host cuticle. Plant Cell. 1992, 4, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2019, 84, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Pascholati, S.F.; Yoshioka, H.; Kunoh, H.; Nicholson, R.L. Preparation of the infection court by Erysiphe graminis f. sp. hordei: Cutinase is a component of the conidial exudate. Physiol. Mol. Plant Pathol. 1992, 41, 53–59. [Google Scholar] [CrossRef]
- Pascholati, S.F.; Deising, H.; Leiti, B.; Anderson, D.; Nicholson, R.L. Cutinase and non-specific esterase activities in the conidial mucilage of Colletotrichum graminicola. Physiol. Mol. Plant Pathol. 1993, 42, 37–51. [Google Scholar] [CrossRef]
- Van Kan, J.A.L.; Van’t Klooster, J.W.; Wagemakers, C.A.M.; Dees, D.C.T.; Van Der Vlugt-Bergmans, C.J.B. Cutinase A of Botrytis cinerea is expressed, but not essential, during penetration of gerbera and tomato. Mol. Plant Microbe Interact. 1997, 10, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vlugt-Bergmans, C.J.B.; Wagemakers, C.A.M.; Van Kan, J.A.L. Cloning and expression of the cutinase a gene of Botrytis cinerea. Mol. Plant Microbe Interact. 1997, 10, 21–29. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Kolattukudy, P.E. Mechanism by which contact with plant cuticle triggers cutinase gene expression in the spores of Fusarium solani f. sp. pisi. Proc. Natl. Acad. Sci. USA 1986, 83, 1704–1708. [Google Scholar] [CrossRef] [Green Version]
- Francis, S.A.; Dewey, F.M.; Gurr, S.J. The Role of Cutinase in germling development and infection by Erysiphe graminis f. sp. hordei. Physiol. Mol. Plant Pathol. 1996, 49, 201–211. [Google Scholar] [CrossRef]
- Gilbert, R.D.; Johnson, A.M.; Dean, R.A. Chemical signals responsible for appressorium formation in the rice blast fungus Magnaporthe grisea. Physiol. Mol. Plant Pathol. 1996, 48, 335–346. [Google Scholar] [CrossRef]
- Van den Ende, G.; Linskens, H.F. Cutinolytic enzymes in relation to pathogenesis. Annu. Rev. Phytopathol. 1974, 12, 247–258. [Google Scholar] [CrossRef]
- Lin, T.S.; Kolattukudy, P.E. Structural studies on cutinase, a glycoprotein containing novel amino acids and glucuronic acid amide at the N terminus. Eur. J. Biochem. 1980, 106, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.L.; Aires-Barros, M.R.; Cabral, J.M.S. Cutinase: From molecular level to bioprocess development. Biotechnol. Bioeng. 1999, 66, 17–34. [Google Scholar] [CrossRef]
- Tenhaken, R.; Arnemann, M.; Köhler, G.; Barz, W. Characterization and cloning of cutinase from Ascochyta rabiei. Z. Naturforsch. C J. Biosci. 1997, 52, 197–208. [Google Scholar] [CrossRef]
- Wang, G.-Y.; Hammock, B.; Lee, Y.-M.; Bostock, R. Affinity purification and characterization of a cutinase from the fungal plant pathogen Monilinia fructicola (Wint.) honey. Arch. Biochem. Biophys. 2000, 382, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Belbahri, L.; Calmin, G.; Mauch, F.; Andersson, J.O. Evolution of the cutinase gene family: Evidence for lateral gene transfer of a candidate Phytophthora virulence factor. Gene 2008, 408, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bashi, Z.D.; Roger Rimmer, S.; Khachatourians, G.G.; Hegedus, D.D. Factors governing the regulation of Sclerotinia sclerotiorum cutinase a and polygalacturonase 1 during different stages of infection. Can. J. Microbiol. 2012, 58, 605–616. [Google Scholar] [CrossRef]
- D’Enfert, C. Fungal spore germination: Insights from the molecular genetics of Aspergillus nidulans and Neurospora crassa. Fungal Genet. Biol. 1997, 21, 163–172. [Google Scholar] [CrossRef]
- Zhang, Z.; Henderson, C.; Perfect, E.; Carver, T.L.W.W.; Thomas, B.J.; Skamnioti, P.; Gurr, S.J. Of genes and genomes, needles and haystacks: Blumeria graminis and functionality. Mol. Plant Pathol. 2005, 6, 561–575. [Google Scholar] [CrossRef]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea Pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köller, W.; Smith, F.D.; Reynolds, K.L. Phenotypic instability of flusilazole sensitivity in Venturia inaequalis. Plant Pathol. 1991, 40, 608–611. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; ISBN 0080473784. [Google Scholar]
- Gui, Y.J.; Zhang, W.Q.; Zhang, D.D.; Zhou, L.; Short, D.P.G.; Wang, J.; Ma, X.F.; Li, T.G.; Kong, Z.Q.; Wang, B.L.; et al. A Verticillium dahliae extracellular cutinase modulates plant immune responses. Mol. Plant Microbe Interact. 2018, 31, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zhang, B.; Gai, Y.; Sun, X.; Chung, K.R.; Li, H. Cell-wall-degrading enzymes required for virulence in the host selective toxin-producing necrotroph Alternaria alternata of citrus. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Fu, H.; Chung, K.R.; Gai, Y.; Mao, L.; Li, H. The basal transcription factor II H subunit Tfb5 is required for stress response and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant. Pathol. 2020, 21, 1337–1352. [Google Scholar] [CrossRef] [PubMed]
- Maiti, I.B.; Kolattukudy, P.E. Prevention of fungal infection of plants by specific inhibition of cutinase a relationship between DNA helix stability and recognition sites for RNA polymerase. Sceince 1979, 205, 507–508. [Google Scholar] [CrossRef]
- Dantzig, A.H.; Zuckerman, S.H.; Andonov-Roland, M.M. Isolation of a Fusarium solani mutant reduced in cutinase activity and virulence. J. Bacteriol. 1986, 168, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.M.; Flaishman, M.; Kolattukudy, P.E. Cutinase gene disruption in Fusarium solani f. sp. pisi decreases Its Virulence on Pea. Plant Cell 1994, 6, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ashby, A.M.; Johnstone, K. Molecular evidence that the extracellular cutinase Pbc1 is required for pathogenicity of Pyrenopeziza brassicae on oilseed rape. Mol. Plant Microbe Interact. 2003, 16, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, J.; Li, D.W.; Zheng, L.; Huang, J. CglCUT1 gene required for cutinase activity and pathogenicity of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera. Eur. J. Plant Pathol. 2017, 147, 103–114. [Google Scholar] [CrossRef]
- Lee, M.H.; Chlu, C.M.; Roubtsova, T.; Chou, C.M.; Bostock, R.M. Overexpression of a redox-regulated cutinase gene, MfCUTI, increases virulence of the brown rot pathogen Monilinia fructicola on Prunus spp. Mol. Plant Microbe Interact. 2010, 23, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Dickman, M.B.; Podila, G.K.; Kolattukudy, P.E. Insertion of cutinase gene into a wound pathogen enables it to infect intact host. Nature 1989, 342, 446–448. [Google Scholar] [CrossRef]
- Rajarammohan, S. Redefining Plant-Necrotroph Interactions: The Thin Line Between Hemibiotrophs and Necrotrophs. Front. Microbiol. 2021, 12, 673518. [Google Scholar] [CrossRef] [PubMed]
- Van Kan, J.A.; Shaw, M.W.; Grant-Downton, R.T. Botrytis species: Relentless necrotrophic thugs or endophytes gone rogue? Mol. Plant Pathol. 2014, 15, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Spanu, P.D.; Panstruga, R. Editorial: Biotrophic plant-microbe interactions. Front. Plant Sci. 2017, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Skamnioti, P.; Furlong, R.F.; Gurr, S.J. Evolutionary history of the ancient cutinase family in five filamentous ascomycetes reveals differential gene duplications and losses and in Magnaporthe grisea shows evidence of sub- and neo-functionalization. New Phytol. 2008, 180, 711–721. [Google Scholar] [CrossRef]
- Schmitt, I. Fruiting Body Evolution in the Ascomycota: A Molecular Perspective Integrating Lichenized and Non-Lichenized Groups. In Evolution of Fungi and Fungal-Like Organisms; Springer: Berlin/Heidelberg, Germany, 2011; pp. 187–204. [Google Scholar]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Köller, W.; Yao, C.; Trial, F.; Parker, P.M. Role of cutinase in the invasion of plants. Can. J. Bot. 1995, 73, 1109–1118. [Google Scholar] [CrossRef]
- Novy, V.; Carneiro, L.V.; Shin, J.H.; Larsbrink, J.; Olsson, L. Phylogenetic analysis and in-depth characterization of functionally and structurally diverse CE5 cutinases. J. Biol. Chem. 2021, 297, 101302. [Google Scholar] [CrossRef] [PubMed]
- Krijger, J.J.; Thon, M.R.; Deising, H.B.; Wirsel, S.G.R. Compositions of fungal secretomes indicate a greater impact of phylogenetic histroy than lifestyle adaptation. BMC Genom. 2014, 15, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandaranayaka, E.P.J.; Frenkel, O.; Elad, Y.; Prusky, D.; Harel, A. Network analysis exposes core functions in major lifestyles of fungal and oomycete plant pathognes. BMC Genom. 2019, 20, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arya, G.C.; Cohen, H. The Multifaceted Roles of Fungal Cutinases during Infection. J. Fungi 2022, 8, 199. https://doi.org/10.3390/jof8020199
AMA Style
Arya GC, Cohen H. The Multifaceted Roles of Fungal Cutinases during Infection. Journal of Fungi. 2022; 8(2):199. https://doi.org/10.3390/jof8020199
Chicago/Turabian StyleArya, Gulab Chand, and Hagai Cohen. 2022. "The Multifaceted Roles of Fungal Cutinases during Infection" Journal of Fungi 8, no. 2: 199. https://doi.org/10.3390/jof8020199
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.