
Inadvertent Selection of a Pathogenic Fungus Highlights Areas of Concern in Human Clinical Practices

 , ,
, ,  and
and 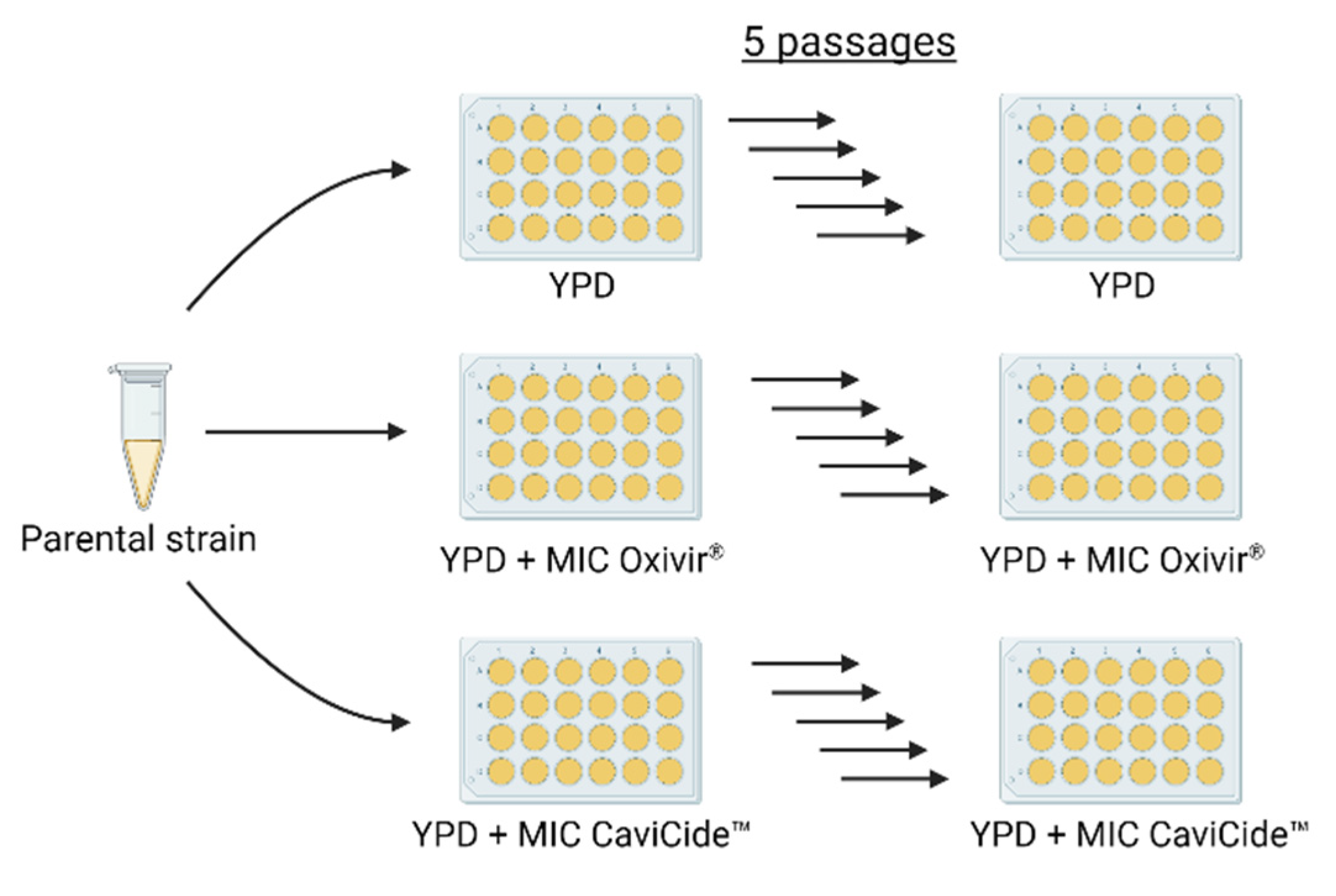{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing
2.2. Passaging Experiments
2.3. Minimum Inhibitory Concentration Assays
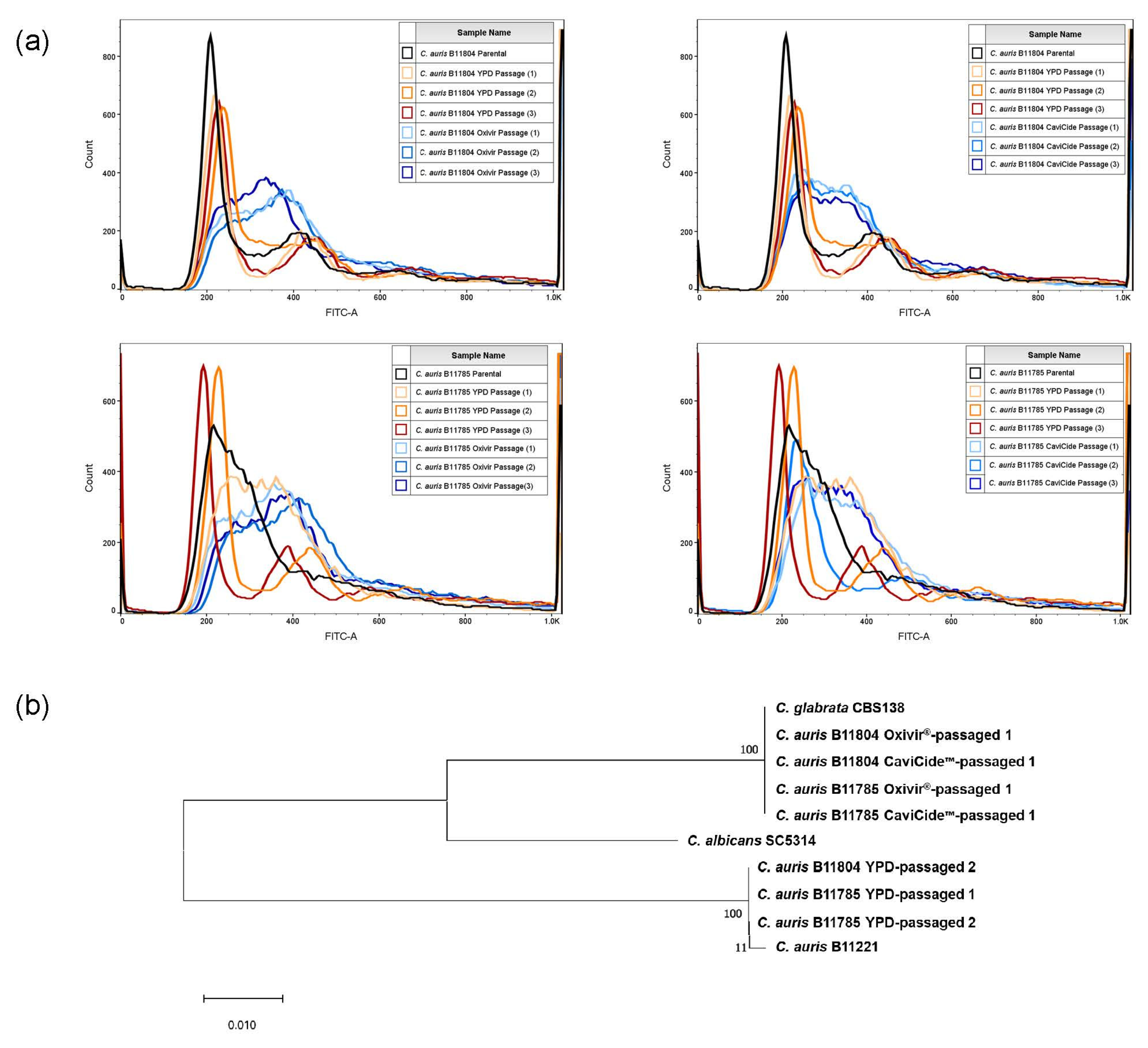
2.4. Flow Cytometry
2.5. Whole-Genome Sequencing
2.6. Purification and Verification of C. auris and C. glabrata Isolates
2.7. Defined Mixture Passaging Experiment and Relative Proportion Determination
3. Results
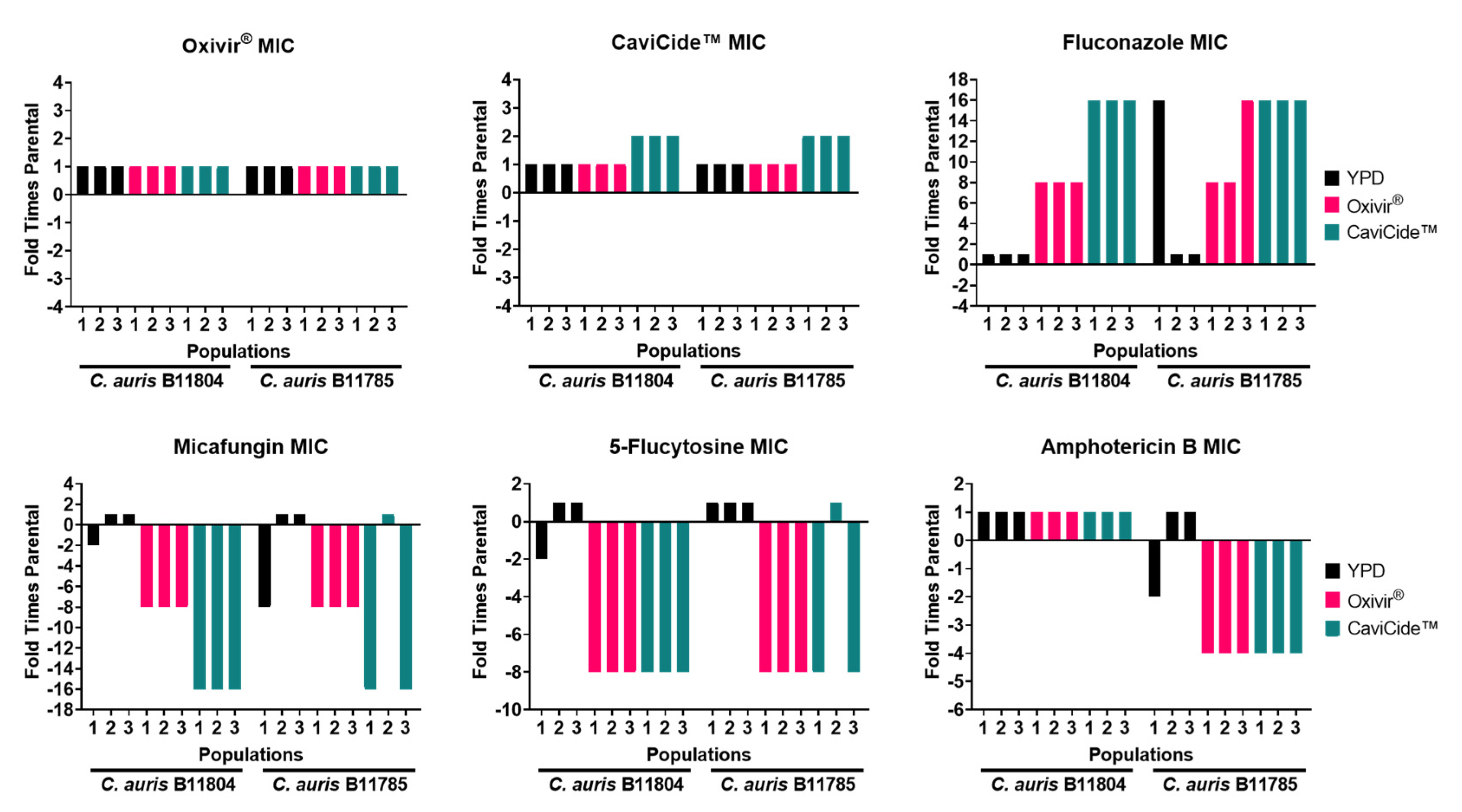
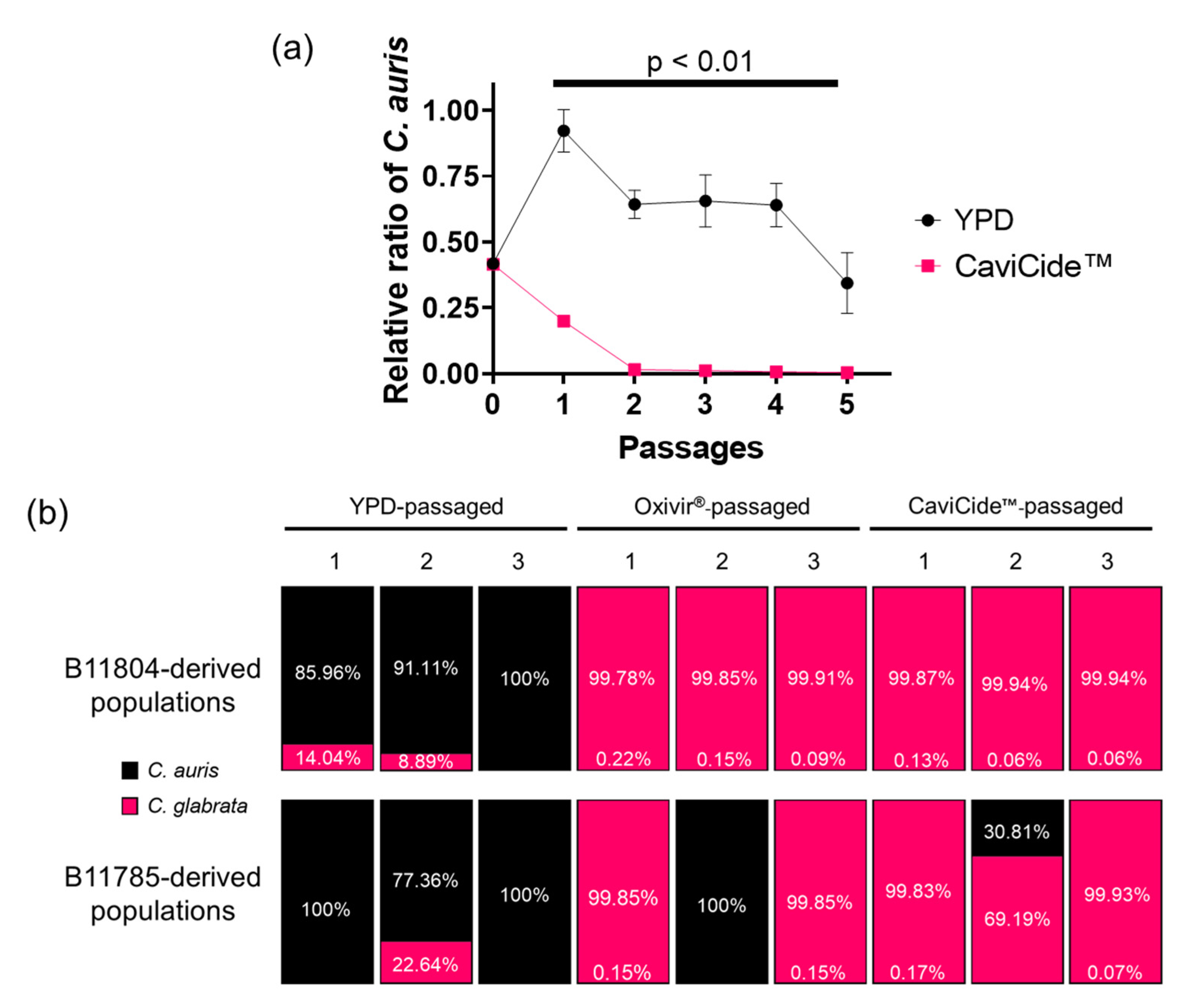
3.1. Serial Exposure of C. auris Clinical Isolates to Hospital Cleaners Selected for C. glabrata Outgrowth
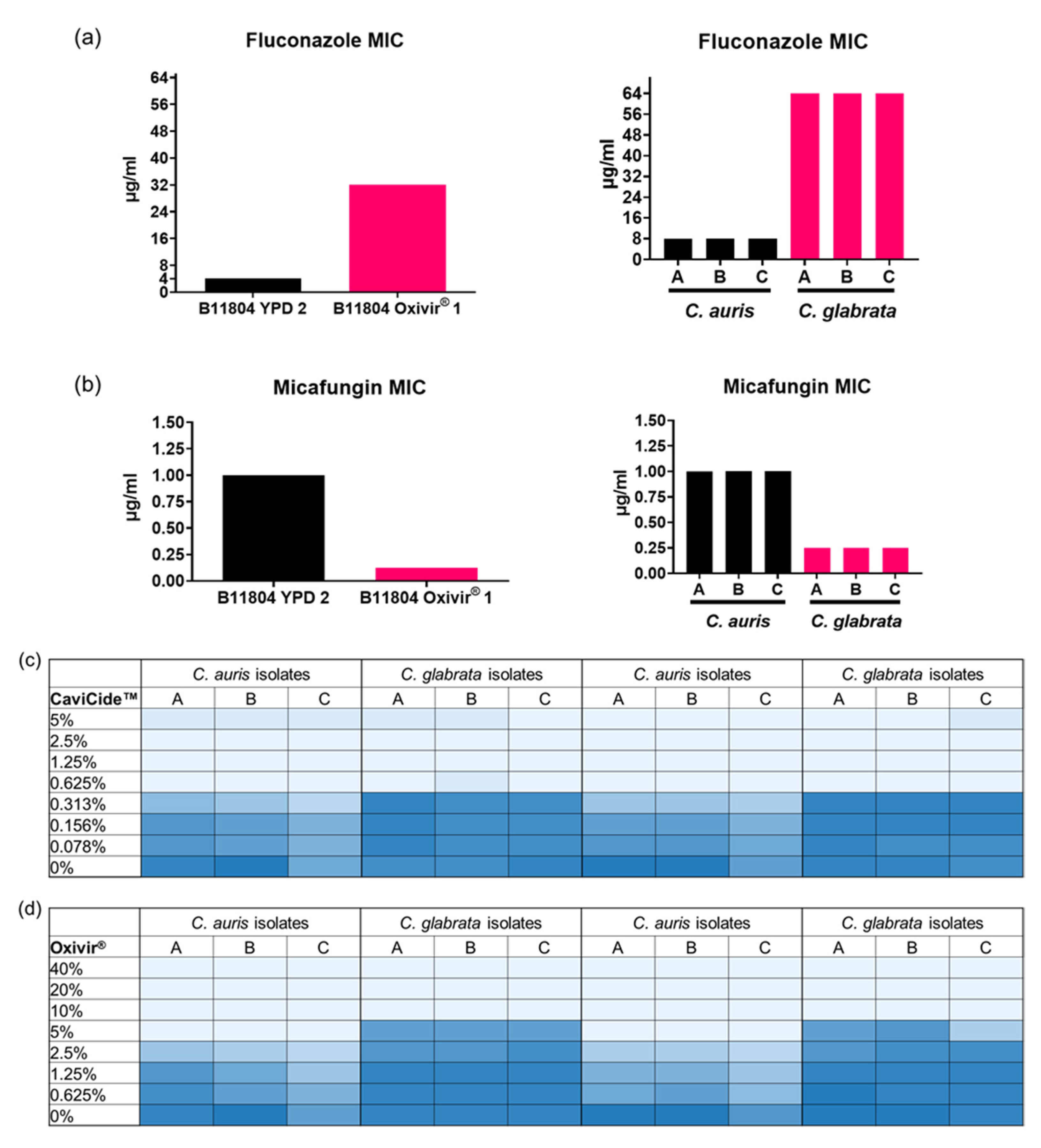
3.2. C. glabrata Tolerated Higher Concentrations of Hospital Cleaner than C. auris
3.3. C. auris Was Quickly Outgrown in Cleaner-Passaged Mixed Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, J.; Jin, X.; Zhang, Q.; Zheng, Y.; Lin, D.; Yu, Y. Fungicides induced triazole-resistance in Aspergillus fumigatus associated with mutations of TR46/Y121F/T289A and its appearance in agricultural fields. J. Hazard. Mater. 2017, 326, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeanvoine, A.; Rocchi, S.; Bellanger, A.P.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus: A global phenomenon originating in the environment? Méde. Mal. Infect. 2020, 50, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.W.; Carneiro, H.C.S.; Oliveira, L.V.N.; Rocha, K.M.; Freitas, G.J.C.; Costa, M.C.; Magalhães, T.F.F.; Carvalho, V.S.D.; Rocha, C.E.; Ferreira, G.F.; et al. Environmental triazole induces cross-resistance to clinical drugs and affects morphophysiology and virulence of Cryptococcus gattii and C. neoformans. Antimicrob. Agents Chemother. 2017, 62, e01179-17. [Google Scholar] [CrossRef] [Green Version]
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist. 2010, 16, 91–104. [Google Scholar] [CrossRef]
- Kampf, G. Adaptive microbial response to low-level benzalkonium chloride exposure. J. Hosp. Infect. 2018, 100, e1–e22. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, A. The lurking scourge of multidrug resistant Candida auris in times of COVID-19 pandemic. J. Glob. Antimicrob. Resist. 2020, 22, 175–176. [Google Scholar] [CrossRef]
- Hughes, S.; Troise, O.; Donaldson, H.; Mughal, N.; Moore, L.S.P. Bacterial and fungal coinfection among hospitalized patients with COVID-19: A retrospective cohort study in a UK secondary-care setting. Clin. Microbiol. Infect. 2020, 26, 1395–1399. [Google Scholar] [CrossRef]
- Villanueva-Lozano, H.; Treviño-Rangel, R.d.J.; González, G.M.; Ramírez-Elizondo, M.T.; Lara-Medrano, R.; Aleman-Bocanegra, M.C.; Guajardo-Lara, C.E.; Gaona-Chávez, N.; Castilleja-Leal, F.; Torre-Amione, G.; et al. Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin. Microbiol. Infect. 2021, 27, 813–816. [Google Scholar] [CrossRef]
- Mehta, S.; Pandey, A. Rhino-orbital mucormycosis associated with COVID-19. Cureus 2020, 12, e10726. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Liang, G.; Liu, W. Fungal co-infections associated with global COVID-19 pandemic: A clinical and diagnostic perspective from China. Mycopathologia 2020, 185, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Askari, F.; Sahu, M.S.; Kaur, R. Candida glabrata: A lot more than meets the eye. Microorganisms 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Alfouzan, W. Candida auris: Epidemiology, diagnosis, pathogenesis, antifungal susceptibility, and infection control measures to combat the spread of infections in healthcare facilities. Microorganisms 2021, 9, 807. [Google Scholar] [CrossRef] [PubMed]
- Antifungal Susceptibility Testing and Interpretation |Candida auris|Fungal Diseases| CDC. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 4 October 2021).
- Ademe, M.; Girma, F. Candida auris: From multidrug resistance to pan-resistant strains. Infect. Drug Resist. 2020, 13, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Singh, P.; Wang, Y.; Yadav, A.; Pawar, K.; Singh, A.; Padmavati, G.; Xu, J.; Chowdhary, A. Environmental isolation of Candida auris from the coastal wetlands of Andaman Islands, India. mBio 2021, 12, e03181-20. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Pereira, D.; Formosa-Dague, C.; Leitão, C.; Pais, P.; Ndlovu, E.; Viana, R.; Pimenta, A.I.; Santos, R.; Takahashi-Nakaguchi, A.; et al. From the first touch to biofilm establishment by the human pathogen Candida glabrata: A genome-wide to nanoscale view. Commun. Biol. 2021, 4, 886. [Google Scholar] [CrossRef]
- Kean, R.; Sherry, L.; Townsend, E.; McKloud, E.; Short, B.; Akinbobola, A.; Mackay, W.G.; Williams, C.; Jones, B.L.; Ramage, G. Surface disinfection challenges for Candida auris: An in-vitro study. J. Hosp. Infect. 2018, 98, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Protano, C.; Cammalleri, V.; Romano Spica, V.; Valeriani, F.; Vitali, M. Hospital environment as a reservoir for cross transmission: Cleaning and disinfection procedures. Ann. Ig. 2019, 31, 436–448. [Google Scholar] [CrossRef]
- Infection Prevention and Control for Candida auris|Candida auris|Fungal Diseases|CDC. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-infection-control.html (accessed on 4 October 2021).
- M27Ed4: Broth Dilution Antifungal Susceptibility, Yeasts. Available online: https://clsi.org/standards/products/microbiology/documents/m27/ (accessed on 17 September 2021).
- Almeida, A.J.; Martins, M.; Carmona, J.A.; Cano, L.E.; Restrepo, A.; Leão, C.; Rodrigues, F. New insights into the cell cycle profile of Paracoccidioides brasiliensis. Fungal Genet. Biol. 2006, 43, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Welsh, R.M.; Bentz, M.L.; Shams, A.; Houston, H.; Lyons, A.; Rose, L.J.; Litvintseva, A.P. Survival, persistence, and isolation of the emerging multidrug-resistant pathogenic yeast Candida auris on a plastic health care surface. J. Clin. Microbiol. 2017, 55, 2996–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.-J.; Nelliat, A. A Double-edged sword: Aneuploidy is a prevalent strategy in fungal adaptation. Genes 2019, 10, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revie, N.M.; Iyer, K.R.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Evolution, mechanisms and impact. Curr. Opin. Microbiol. 2018, 45, 70–76. [Google Scholar] [CrossRef]
- Cormack, B.P.; Falkow, S. Efficient homologous and illegitimate recombination in the opportunistic yeast pathogen Candida glabrata. Genetics 1999, 151, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, S.; Griffiths, W.; Dietz, L.; Horve, P.; Nunez, S.; Hu, J.; Shen, J.; Fretz, M.; Bi, C.; Xu, Y.; et al. From one species to another: A review on the interaction between chemistry and microbiology in relation to cleaning in the built environment. Indoor Air 2019, 29, 880–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, R.A.; Hota, B. Contamination, disinfection, and cross-Colonization: Are hospital surfaces reservoirs for nosocomial infection? Clin. Infect. Dis. 2004, 39, 1182–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piedrahita, C.T.; Cadnum, J.L.; Jencson, A.L.; Shaikh, A.A.; Ghannoum, M.A.; Donskey, C.J. Environmental surfaces in healthcare facilities are a potential source for transmission of Candida auris and other Candida species. Infect. Control Hosp. Epidemiol. 2017, 38, 1107–1109. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eagan, J.L.; Steffan, B.N.; Ortiz, S.C.; Drott, M.T.; Goldman, G.H.; Hull, C.M.; Keller, N.P.; Bastos, R.W. Inadvertent Selection of a Pathogenic Fungus Highlights Areas of Concern in Human Clinical Practices. J. Fungi 2022, 8, 157. https://doi.org/10.3390/jof8020157
Eagan JL, Steffan BN, Ortiz SC, Drott MT, Goldman GH, Hull CM, Keller NP, Bastos RW. Inadvertent Selection of a Pathogenic Fungus Highlights Areas of Concern in Human Clinical Practices. Journal of Fungi. 2022; 8(2):157. https://doi.org/10.3390/jof8020157
Chicago/Turabian StyleEagan, Justin L., Breanne N. Steffan, Sébastien C. Ortiz, Milton T. Drott, Gustavo H. Goldman, Christina M. Hull, Nancy P. Keller, and Rafael W. Bastos. 2022. "Inadvertent Selection of a Pathogenic Fungus Highlights Areas of Concern in Human Clinical Practices" Journal of Fungi 8, no. 2: 157. https://doi.org/10.3390/jof8020157