
Fine Identification and Classification of a Novel Beneficial Talaromyces Fungal Species from Masson Pine Rhizosphere Soil

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of the Strain
2.2. DNA Extraction, PCR Amplification, and Sequencing of Strain JP-NJ4
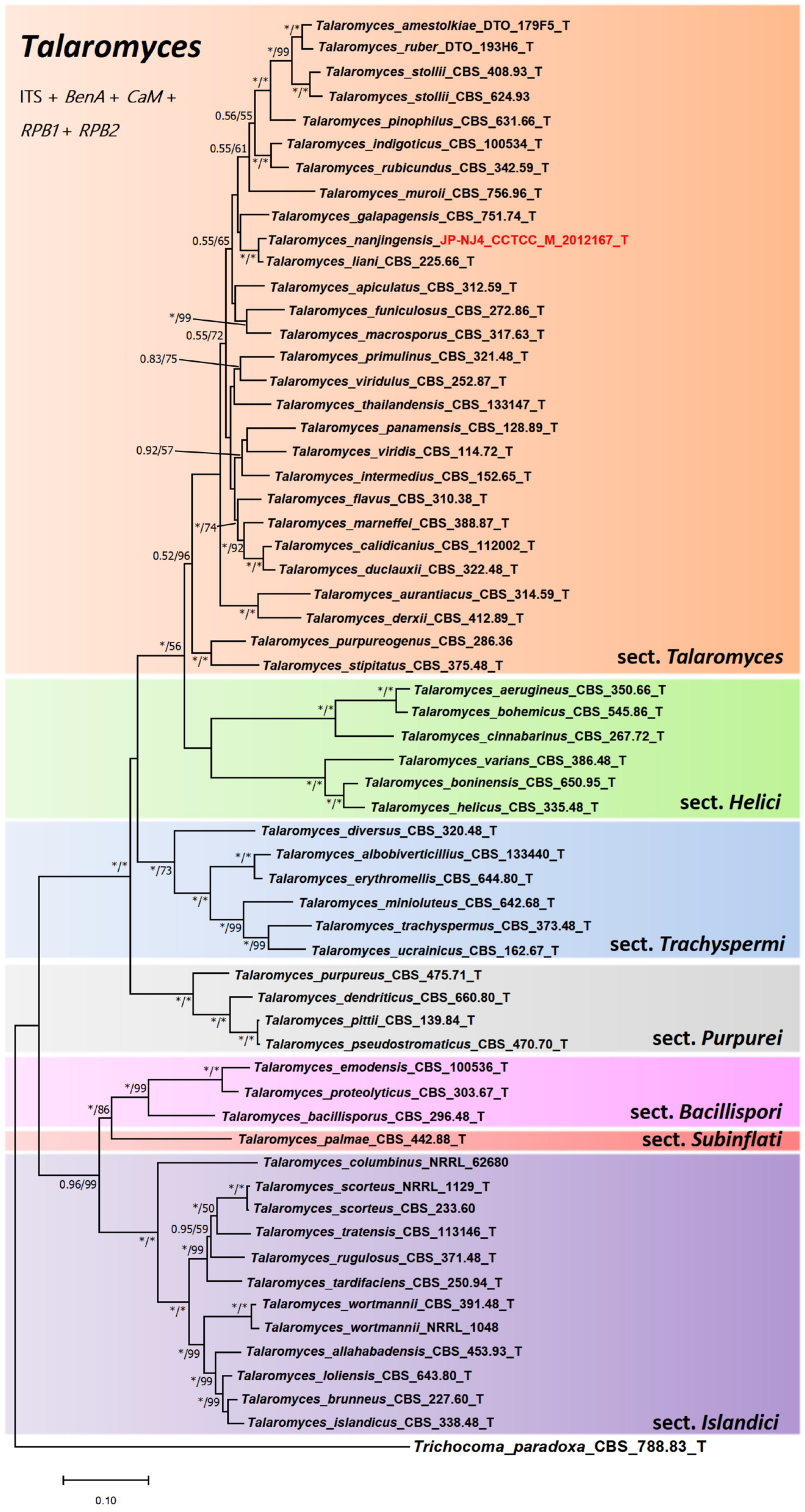
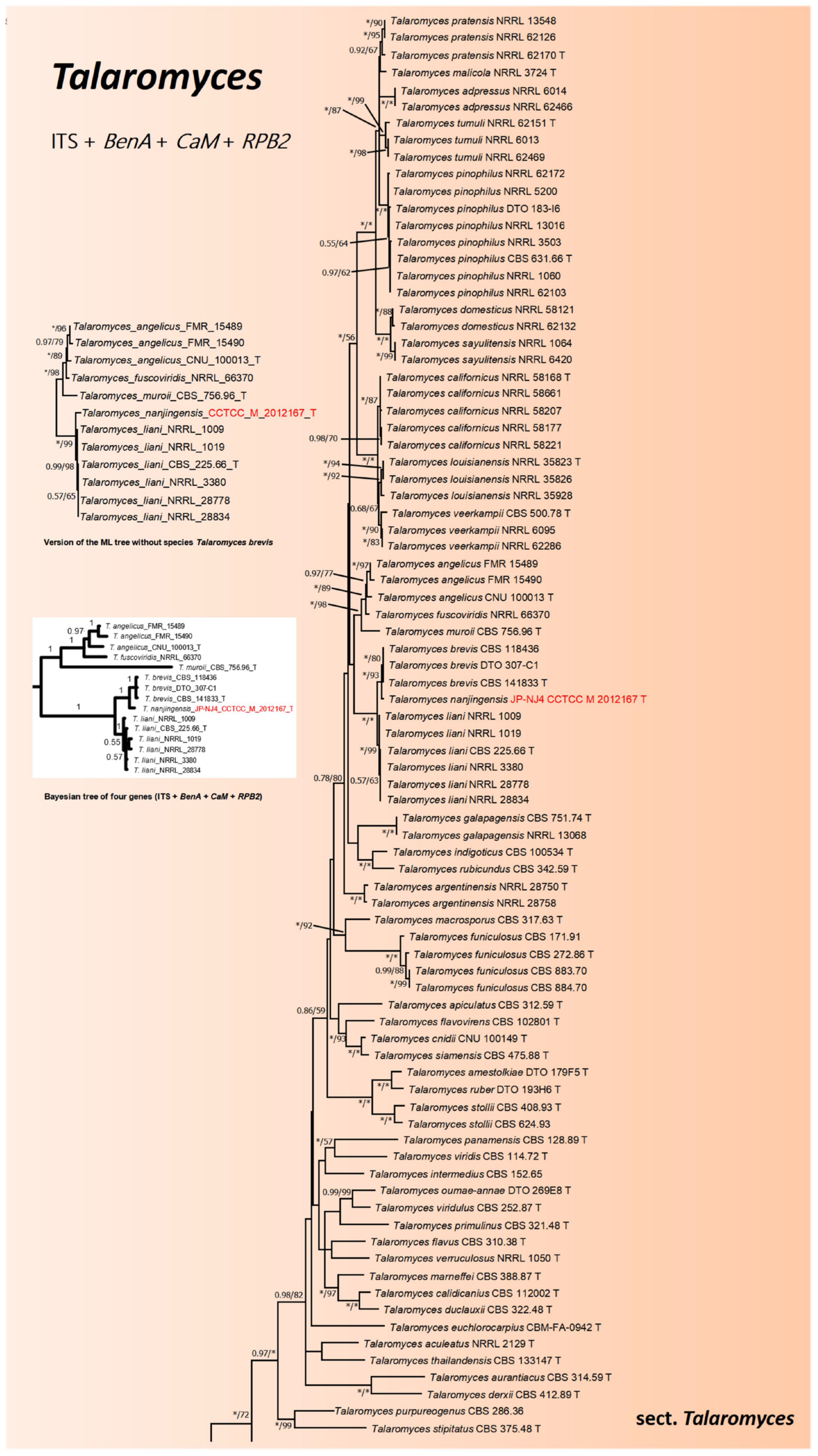
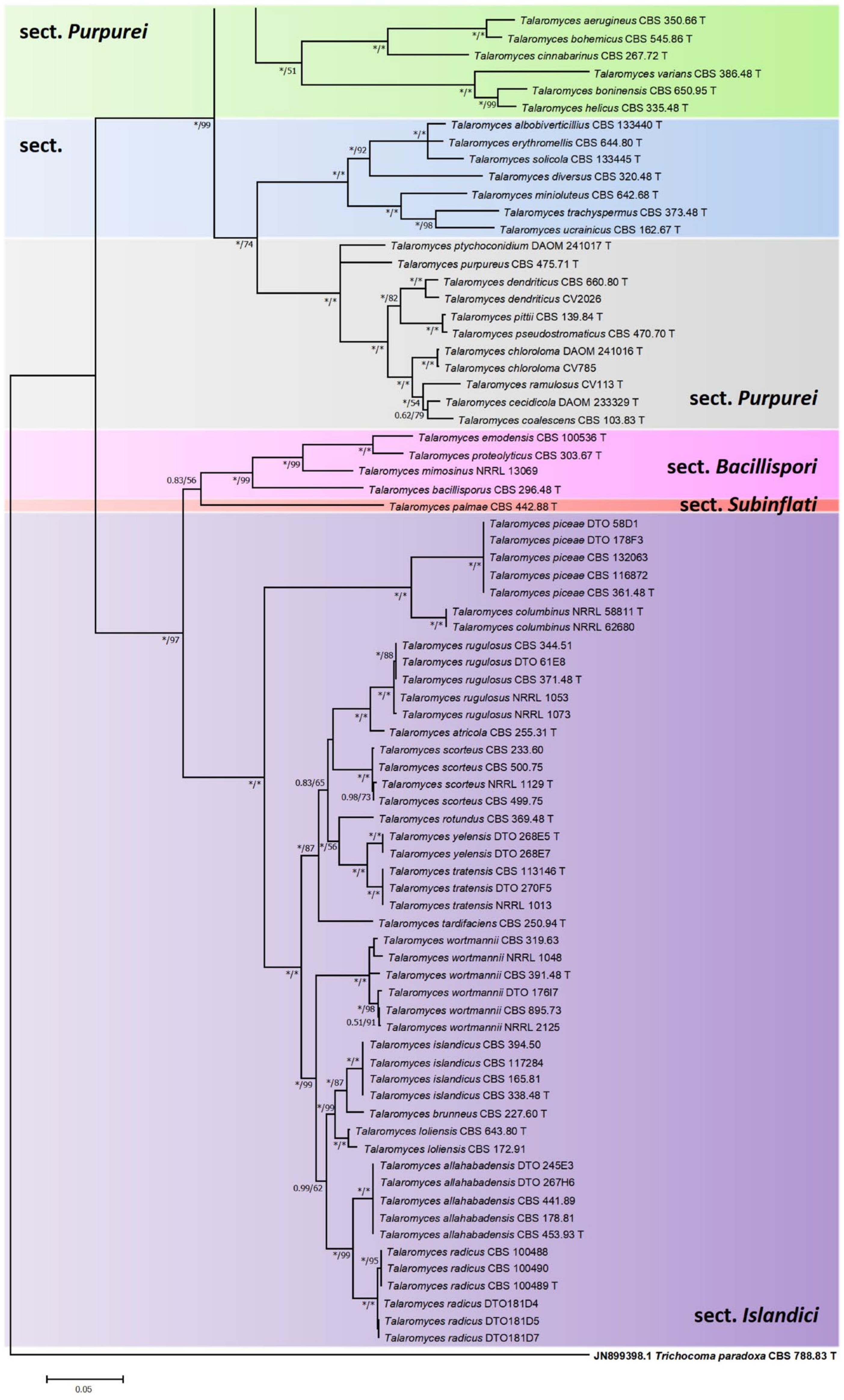
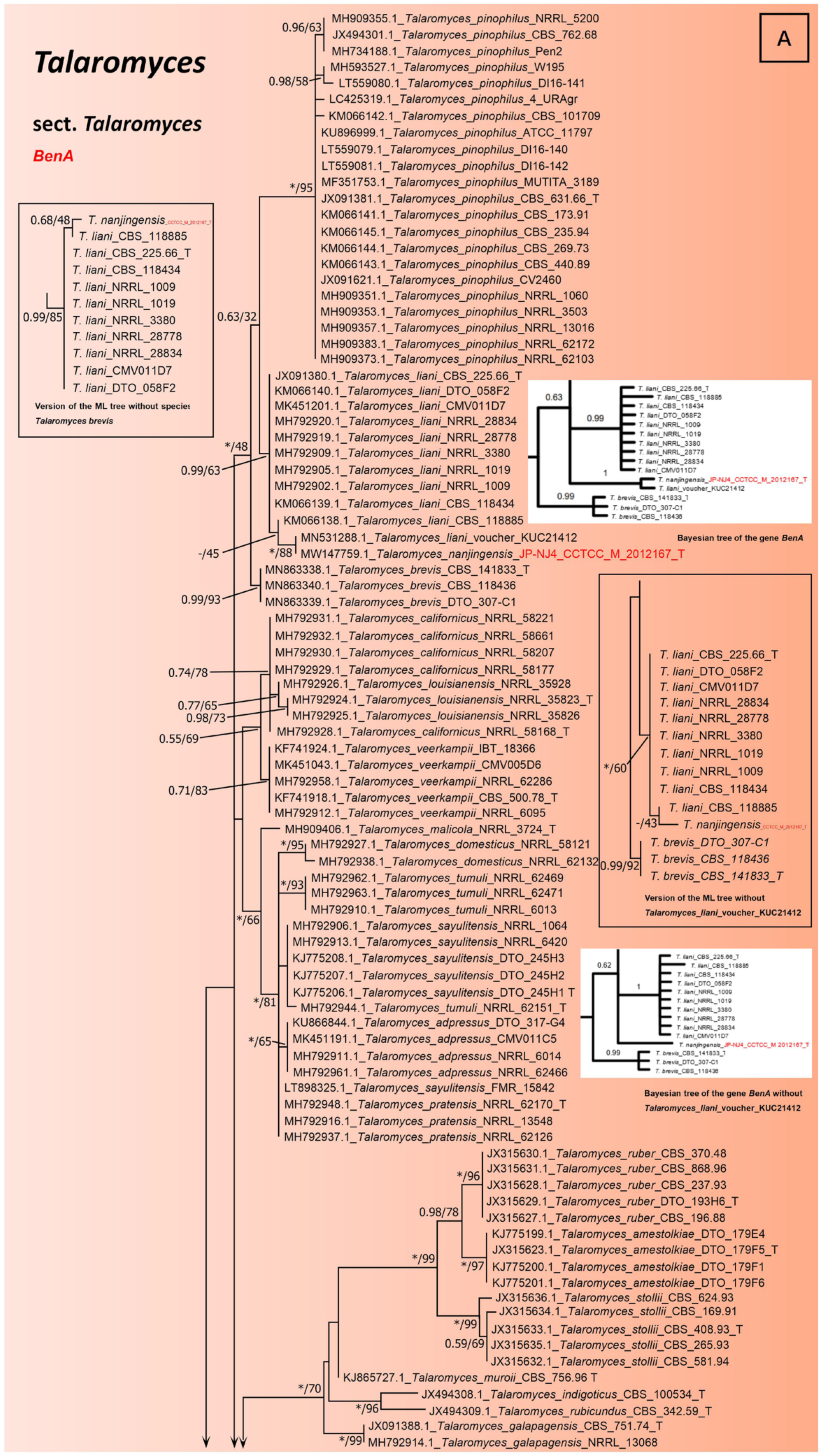
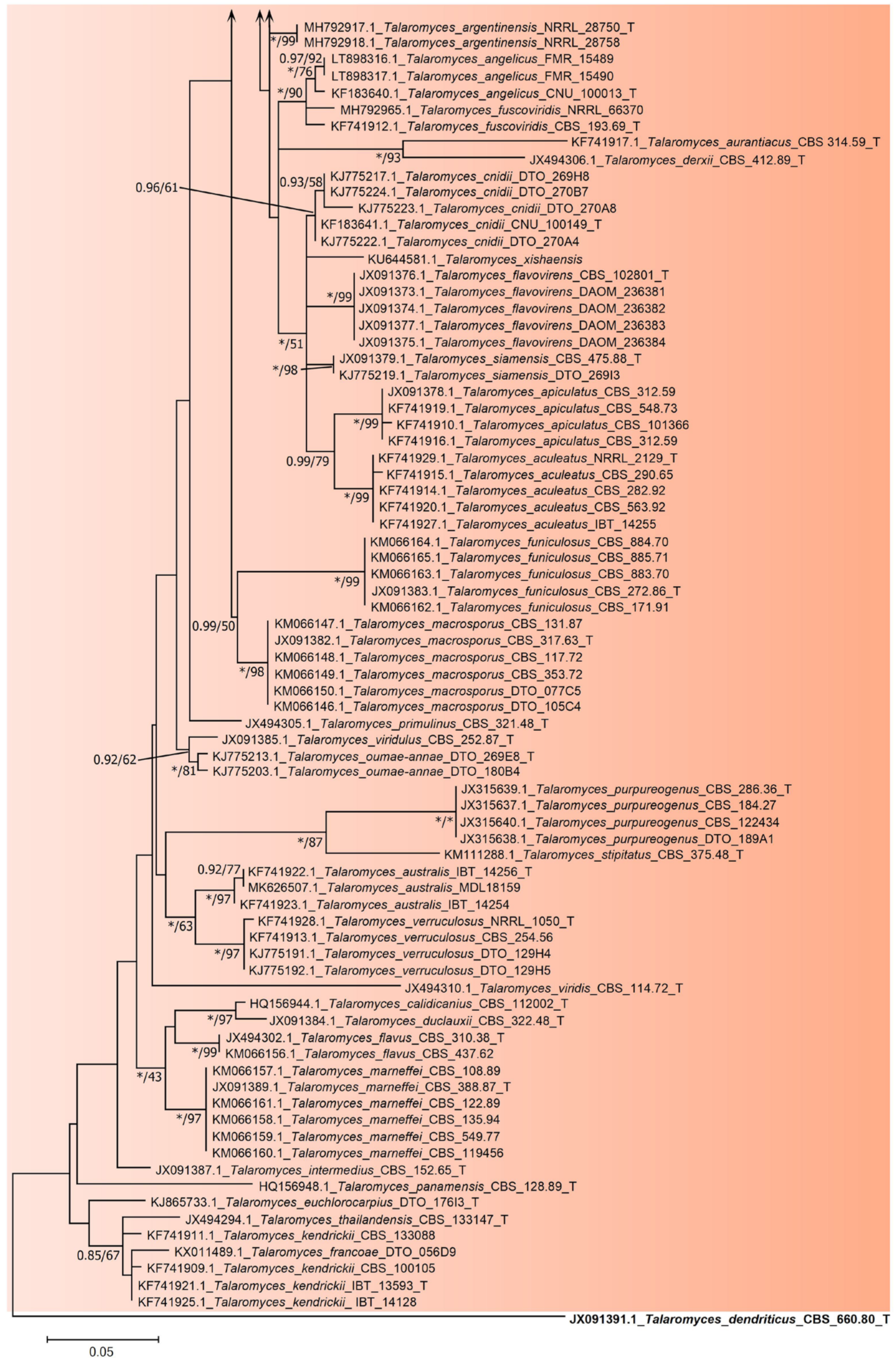
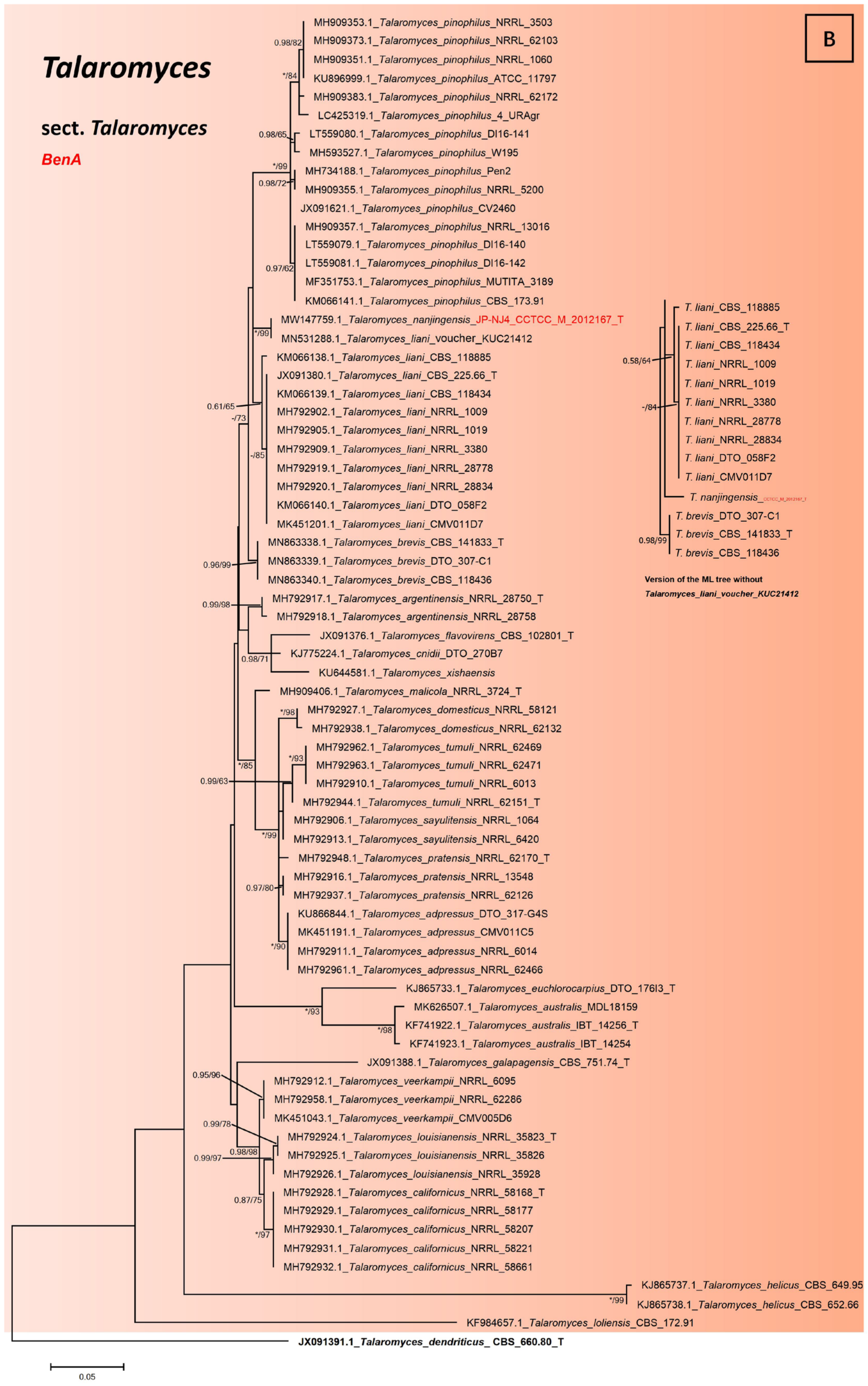
2.3. Phylogenetic Tree Construction of Strain JP-NJ4
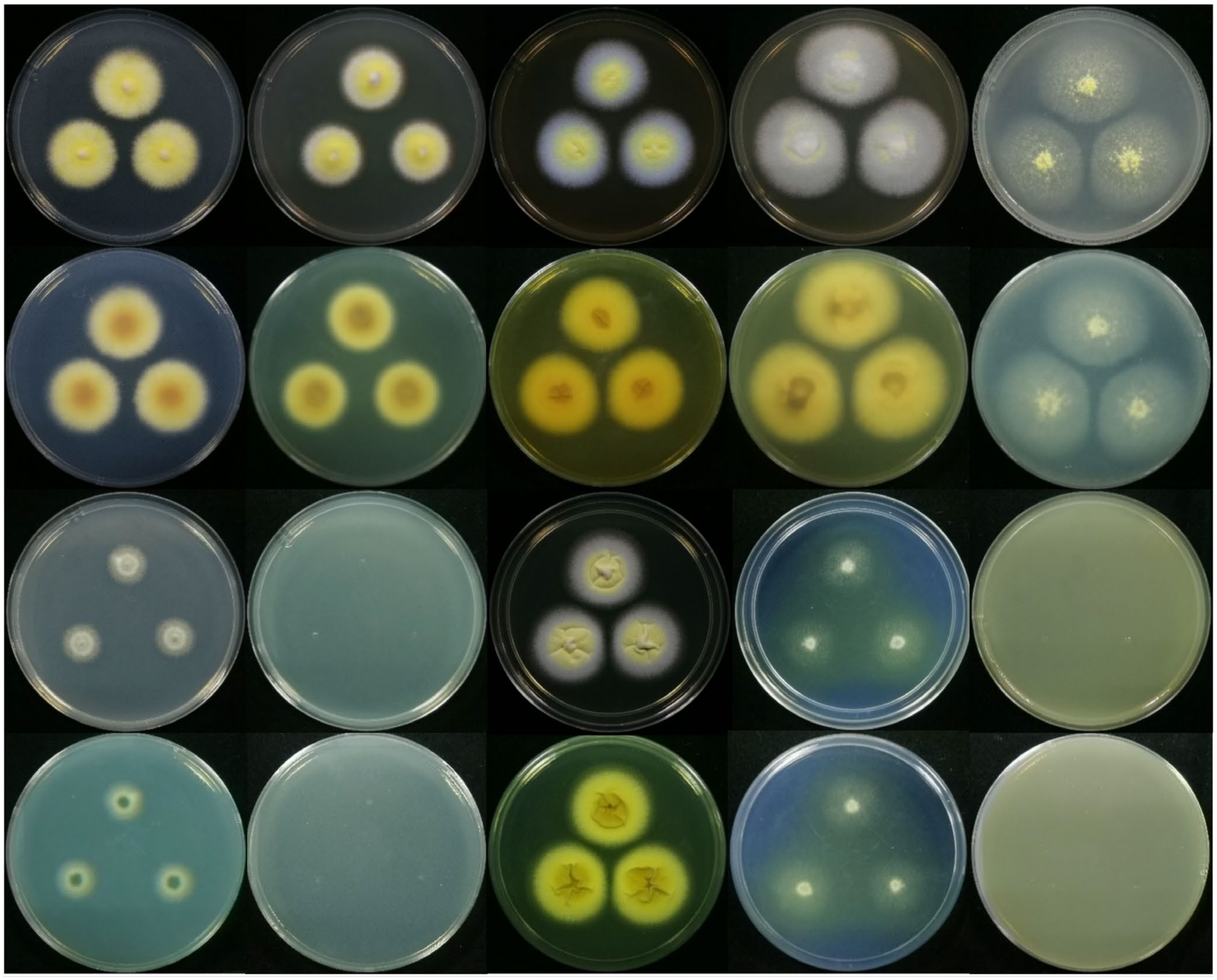
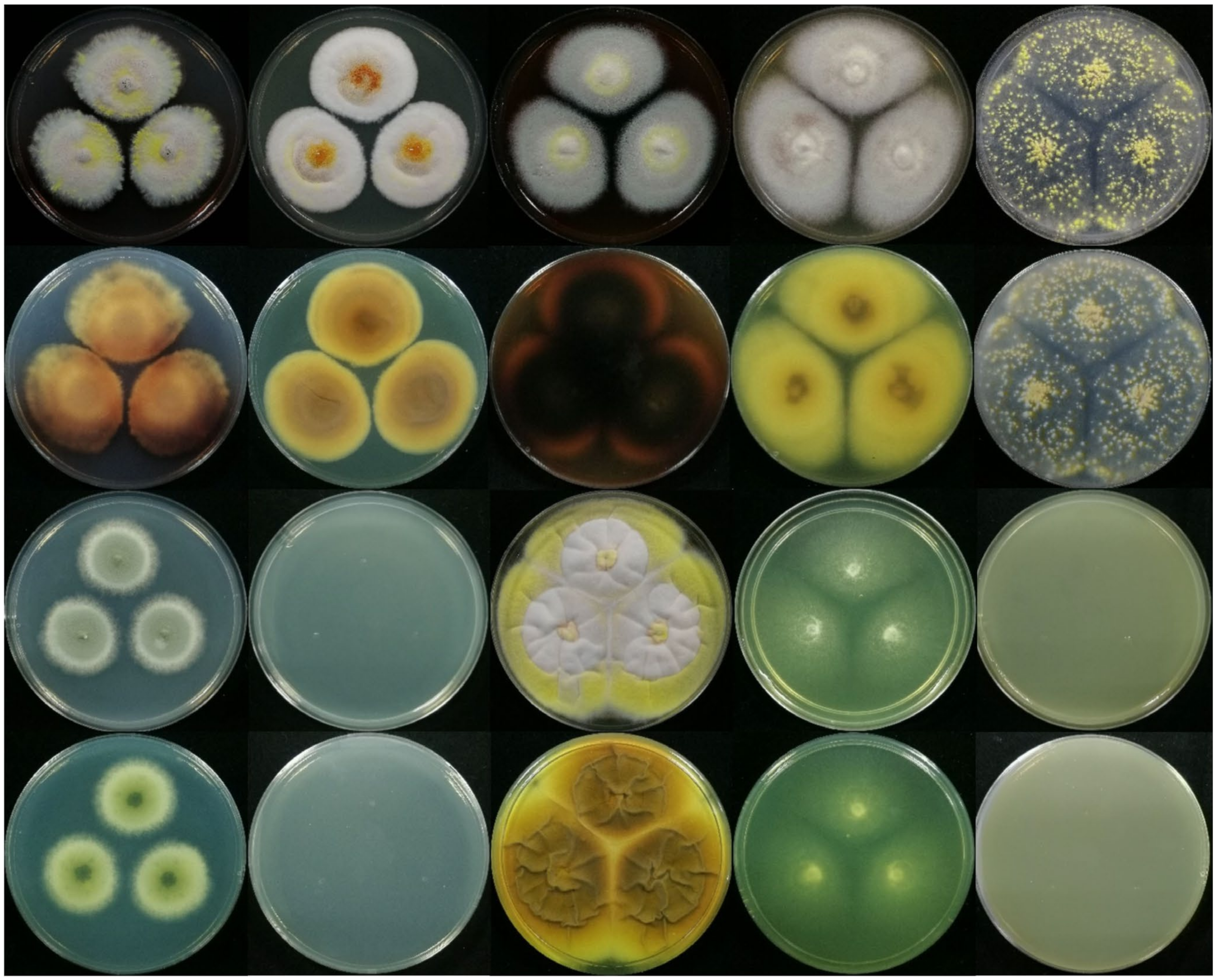
2.4. Observation on the Morphological Characteristics of Strain JP-NJ4
3. Results
3.1. Taxonomy of Strain JP-NJ4
Taxonomy
3.2. Phylogeny-Based Species Identification
3.3. Species Identification Based on Macromorphology and Micromorphology
4. Discussion
- ACCC: Agricultural Culture Collection of China.
- ATCC: American Type Culture Collection, Manassas, VA, USA (WDCM 1) http://www.atcc.org/, accessed on 18 January 2022;
- CABI: Centre for Agriculture and Bioscience International (International Mycological Institute, CABI Genetic Resource Collection).
- CBS: culture collection of the CBS-KNAW Fungal Biodiversity Centre, Utrecht, Netherlands (WDCM 133) http://www.cbs.knaw.nl/databases/index.htm, accessed on 18 January 2022.
- DTO: internal culture collection of CBS-KNAW Fungal Biodiversity Centre; IMI, CABI Genetic Resources Collection, Surrey, UK (WDCM 214) http://www.cabi.org/, accessed on 18 January 2022.
- FERM: (Patent and Bio-Resource Center, National Institute of Advanced Industrial Science and Technology-AIST).
- FMR: facultad de medicina, Universidad de Oviedo. 33071-Oviedo. Spain. Institute de Investigaciones Biomidicas C.S.I.C., Facultad de Medicina UAM, E-28029 Madrid, Spain.
- HMAS: Fungarium of Institute of Microbiology.
- IBT: culture collection of Center for Microbial Biotechnology (CMB) at Department of Systems Biology, Technical University of Denmark (WDCM 758) http://www.biocentrum.dtu.dk/, accessed on 18 January 2022.
- MUCL: Mycotheque de l’Universite catholique de Louvain, Leuven, Belgium (WDCM 308).
- NBRC: Biological Resource Center, NITE.
- NRRL: ARS Culture Collection, U.S. Department of Agriculture, Peoria, Illinois, USA (WDCM 97) http://nrrl.ncaur.usda.gov/, accessed on 18 January 2022.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mehta, P.; Sharma, R.; Putatunda, C.; Walia, A. Endophytic Fungi: Role in Phosphate Solubilization. In Advances in Endophytic Fungal Research; Singh, B., Ed.; Springer: Cham, Switzerland, 2019; pp. 183–209. [Google Scholar]
- Zheng, B.X.; Ibrahim, M.; Zhang, D.P.; Bi, Q.F.; Li, H.Z.; Zhou, G.W.; Ding, K.; Penuelas, J.; Zhu, Y.G.; Yang, X.R. Identification and characterization of inorganic-phosphate-solubilizing bacteria from agricultural fields with a rapid isolation method. AMB Express 2018, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.C.; Chun-Mei, D.U.; Ping, W.X.; Guan, H.Y.; Bao-Xing, X.U. Advance in Phosphorus-Dissolving Microbes. J. Microbiol. 2006, 2, 73–78, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Malviya, J.; Singh, K.; Joshi, V. Effect of Phosphate Solubilizing Fungi on Growth and Nutrient Uptake of Ground nut (Arachis hypogaea) Plants. Adv. Biores. 2011, 2, 110–113. [Google Scholar]
- Saxena, J.; Rawat, J.; Sanwal, P. Enhancement of Growth and Yield of Glycine max Plants with Inoculation of Phosphate Solubilizing Fungus Aspergillus niger K7 and Biochar Amendment in Soil. Commun. Soil. Sci. Plan. 2016, 47, 2334–2347. [Google Scholar] [CrossRef]
- Qiao, H.; Sun, X.R.; Wu, X.Q.; Li, G.E.; Li, D.W. The phosphate-solubilising ability of Penicilium guanacastense and its effects on the growth of Pinus massoniana in phosphate limiting conditions. Biol. Open. 2019, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Taniwaki, M.H.; Hocking, A.D.; Pitt, J.I.; Fleet, G.H. Growth and mycotoxin production by food spoilage fungi under high carbon dioxide and low oxygen atmospheres. Int. J. Food. Microbiol. 2009, 132, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010. [Google Scholar]
- Link, H.F. Observationes in Ordines Plantarum Naturales. Mag. Ges. naturf. Freunde. 1809, 3, 1–42. [Google Scholar]
- Dierckx, R.P. Un essai de revision du genre Penicillium Link. Annales de la Société Scientifique Bruxelles 1901, 25, 90–104. [Google Scholar]
- Breen, J.; Dacre, J.C.; Raistrick, H.; Smith, G. Studies in the biochemistry of micro-organisms. 95. Rugulosin, a crystalline colouring matter of Penicillium rugulosum Thom. Biochem. J. 1955, 60, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Stolk, A.C.; Samson, R.A. The genus Talaromyces—Studies on Talaromyces and related genera II. Stud. Mycol. 1972, 2, 1–65. [Google Scholar] [CrossRef]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces; Academic Press Inc.: London, UK, 1979. [Google Scholar]
- Samson, R.A.; Yilmaz, N.; Houbraken, J.; Spierenburg, H.; Seifert, K.A.; Peterson, S.W.; Varga, J.; Frisvad, J.C. Phylogeny and nomenclature of the genus Talaromyces and taxa accommodated in Penicillium subgenus Biverticillium. Stud. Mycol. 2011, 70, 159–183. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [Green Version]
- Raper, K.B.; Thom, C. A Manual of the Penicillia; The Williams & Wilkins Company: Baltimore, MD, USA, 1949. [Google Scholar]
- Thom, C. The Penicillia; The Williams & Wilkins Company, Baltimore: MD, USA, 1930. [Google Scholar]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Toru Okuda, M.A.K.; Seifert, K.A.; Ando, K. Media and incubation effects on morphological characteristics of Penicillium and Aspergillus. In Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 83–99. [Google Scholar]
- Okuda, T. Variation in colony characteristics of Penicillium strains resulting from minor variations in culture conditions. Mycologia 1994, 86, 259–262. [Google Scholar] [CrossRef]
- Samson, R.A.; Pitt, J.I. General Recommendations. In Advances in Penicillium and Aspergillus Systematics; Samson, R.A., Pitt, J.I., Eds.; Plenum Press: London, UK, 1985; pp. 455–460. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal. Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Xiao-Qin, W.U.; Wang, Z. Phosphate-solubilizing characteristic of a Penicillium pinophilum strain JP-NJ4. Microbiol. China 2014, 9, 1741–1748, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Samson, R.A.; van der Aa, H.A.; de Hoog, G.S. Centraalbureau voor Schimmelcultures: Hundred years microbial resource centre. Stud. Mycol. 2005, 50, 1–8. [Google Scholar] [CrossRef]
- May, T.W.; Redhead, S.A.; Bensch, K.; Hawksworth, D.L.; Lendemer, J.; Lombard, L.; Turland, N.J. Chapter F of the International Code of Nomenclature for algae, fungi, and plants as approved by the 11th International Mycological Congress, San Juan, Puerto Rico, July 2018. IMA Fungus 2019, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Aime, M.C.; Miller, A.N.; Aoki, T.; Bensch, K.; Cai, L.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.D.; Kirk, P.M.; Lucking, R.; et al. How to publish a new fungal species, or name, version 3.0. IMA Fungus 2021, 12, 11. [Google Scholar] [CrossRef]
- Cubero, O.F.; Crespo, A.; Fatehi, J.; Bridge, P.D. DNA extraction and PCR amplification method suitable for fresh, herbarium-stored, lichenized, and other fungi. Plant. Syst. Evol. 1999, 216, 243–249. [Google Scholar] [CrossRef]
- De Hoog, G.S.; van den Ende, A.H.G.G. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef]
- Houbraken, J.; Spierenburg, H.; Frisvad, J.C. Rasamsonia, a new genus comprising thermotolerant and thermophilic Talaromyces and Geosmithia species. Anton. Leeuw. Int. J. G. 2012, 101, 403–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Lousie, G.N.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from Filamentous Ascomycetes. Appl. Environ. Microb. 1995, 61, 1320–1330. [Google Scholar] [CrossRef]
- Masclaux, F.; Gueho, E.; de Hoog, G.S.; Christen, R. Phylogenetic relationships of human-pathogenic Cladosporium (Xylohypha) species inferred from partial LS rRNA sequences. J. Med. Vet. Mycol. 1995, 33, 327–338. [Google Scholar] [CrossRef]
- Peterson, S.W.; Vega, F.E.; Posada, F.; Nagai, C. Penicillium coffeae, a new endophytic species isolated from a coffee plant and its phylogenetic relationship to P. fellutanum, P. thiersii and P. brocae based on parsimony analysis of multilocus DNA sequences. Mycologia 2005, 97, 659–666. [Google Scholar] [CrossRef]
- Rivera, K.G.; Seifert, K.A. A taxonomic and phylogenetic revision of the Penicillium sclerotiorum complex. Stud. Mycol. 2011, 70, 139–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, F.; White, T.; Lee, S.H.; Taylor, L.; Shawetaylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A.G., Sninsky, D.H., John, J., White, J.J., Thomas, J., Eds.; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- El-Esawi, M.A.; Witczak, J.; Abomohra, A.E.; Ali, H.M.; Elshikh, M.S.; Ahmad, M. Analysis of the Genetic Diversity and Population Structure of Austrian and Belgian Wheat Germplasm within a Regional Context Based on DArT Markers. Genes 2018, 9, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C. Physiological criteria and mycotoxin production as AIDS in identification of common asymmetric penicillia. Appl. Environ. Microbiol. 1981, 41, 568–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, C. Manual and Atlas of the Penicillia.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982. [Google Scholar]
- Blakeslee, A.F. Lindner’s roll tube method of separation cultures. Phytopathology 1915, 5, 68–69. [Google Scholar]
- David, A. Hay Infusion. Tex. Sci. Teach. 1993, 22, 10. [Google Scholar]
- Hocking, A.D.; Pitt, J.I. Dichloran-glycerol medium for enumeration of xerophilic fungi from low-moisture foods. Appl. Environ. Microbiol. 1980, 39, 488–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, M.; Frisvad, J.C.; Tuthill, D.E. Penicillium species diversity in soil and some taxonomic and ecological notes. In Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 309–321. [Google Scholar]
- Pitt, J.I. An appraisal of identification methods for Penicillium species: Novel taxonomic criteria based on temperature and water relations. Mycologia 1973, 65, 1135–1157. [Google Scholar] [CrossRef]
- Kong, H.Z. Flora Fungorum Sinicorum; Science Press: Beijing, China, 2007; Volume 35, pp. 53–54, 187–188. [Google Scholar]
- Schoch, C.L.; Aime, M.C.; Beer, W.D.; Crous, P.W.; Miller, A.N. Using standard keywords in publications to facilitate updates of new fungal taxonomic names. IMA Fungus 2017, 8, 70–73. [Google Scholar] [CrossRef] [Green Version]
- Heo, Y.M.; Lee, H.; Kim, K.; Sun, L.K.; Kim, J.J. Fungal Diversity in Intertidal Mudflats and Abandoned Solar Salterns as a Source for Biological Resources. Mar. Drugs. 2019, 17, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.D.; Chen, A.J.; Houbraken, J.; Frisvad, J.C.; Wu, W.P.; Wei, H.L.; Zhou, Y.G.; Jiang, X.Z.; Samson, R.A. New section and species in Talaromyces. MycoKeys 2020, 68, 75–113. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, R.N.; Bezerra, J.D.P.; Souza-Motta, C.M.; Frisvad, J.C.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Anton. Leeuw. Int. J. G. 2018, 111, 1883–1912. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.J.; Sun, B.D.; Houbraken, J.; Frisvad, J.C.; Yilmaz, N.; Zhou, Y.G.; Samson, R.A. New Talaromyces species from indoor environments in China. Stud. Mycol. 2016, 84, 119–144. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Gene, J.; Guarro, J.; Baseia, I.G.; Garcia, D.; Gusmao, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef]
- Guevara-Suarez, M.; Sutton, D.A.; Gene, J.; Garcia, D.; Wiederhold, N.; Guarro, J.; Cano-Lira, J.F. Four new species of Talaromyces from clinical sources. Mycoses 2017, 60, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.W.; Jurjevic, Z. New species of Talaromyces isolated from maize, indoor air, and other substrates. Mycologia 2017, 109, 537–556. [Google Scholar] [CrossRef]
- Rodriguez-Andrade, E.; Stchigel, A.M.; Terrab, A.; Guarro, J.; Cano-Lira, J.F. Diversity of xerotolerant and xerophilic fungi in honey. IMA Fungus 2019, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Sang, H.; An, T.J.; Kim, C.S.; Shin, G.S.; Sung, G.H.; Yu, S.H. Two novel Talaromyces species isolated from medicinal crops in Korea. J. Microbiol. 2013, 51, 704–708. [Google Scholar] [CrossRef]
- Wang, Q.M.; Zhang, Y.H.; Wang, B.; Wang, L. Talaromyces neofusisporus and T. qii, two new species of section Talaromyces isolated from plant leaves in Tibet, China. Sci. Rep. 2016, 6, 18622. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Lopez-Quintero, C.A.; Vasco-Palacios, A.; Frisvad, J.C.; Theelen, B.; Boekhout, T.; Samson, R.A.; Houbraken, J. Four novel Talaromyces species isolated from leaf litter from Colombian Amazon rain forests. Mycol. Prog. 2016, 15, 1041–1056. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, H.; Koyama, N.; Omura, S.; Tomoda, H. New rugulosins, anti-MRSA antibiotics, produced by Penicillium radicum FKI-3765-2. Org. Lett. 2010, 12, 1572–1575. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Frisvad, J.C.; Knudsen, P.B.; Larsen, T.O. Anticancer and antifungal compounds from Aspergillus, Penicillium and other filamentous fungi. Molecules 2013, 18, 11338–11376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, M.; Taritla, S.; Sharma, A.; Jayabaskaran, C. Antiproliferative and Antioxidative Bioactive Compounds in Extracts of Marine-Derived Endophytic Fungus Talaromyces purpureogenus. Front. Microbiol. 2018, 9, 1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbelia, R.; Louis, B.; Simard, R.R.; Phillipe, T.; Hani, A. Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum and two UV-induced mutants. Fems. Microbiol. Ecol. 1999, 28, 291–295. [Google Scholar] [CrossRef]
- Narikawa, T.; Shinoyama, H.; Fujii, T. A β-Rutinosidase from Penicillium rugulosum IFO 7242 That Is a Peculiar Flavonoid Glycosidase. Biosci. Biotechnol. Bioch. 2000, 64, 1317–1319. [Google Scholar] [CrossRef]
- Pol, D.; Laxman, R.S.; Rao, M. Purification and biochemical characterization of endoglucanase from Penicillium pinophilum MS 20. Indian. J. Biochem. Biophys. 2012, 49, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Hoshino, T.; Inoue, H.; Yano, S. Taxonomic revision of the cellulose-degrading fungus Acremonium cellulolyticus nomen nudum to Talaromyces based on phylogenetic analysis. FEMS. Microbiol. Lett. 2014, 351, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; de Vries, R.P.; Samson, R.A. Modern Taxonomy of Biotechnologically Important Aspergillus and Penicillium Species. Adv. Appl. Microbiol. 2014, 86, 199–249. [Google Scholar] [CrossRef]
- Maeda, R.N.; Barcelos, C.A.; Santa Anna, L.M.; Pereira, N., Jr. Cellulase production by Penicillium funiculosum and its application in the hydrolysis of sugar cane bagasse for second generation ethanol production by fed batch operation. J. Biotechnol. 2013, 163, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahima, T.; Henis, Y. Increasing of Trichoderma hamatum and Talaromyces flavus on root of healthy and useful hosts. In Proceedings of the Biological Control of Soil-Borne Plant Pathogens; CAB International: Wallingford, UK, 1997; pp. 296–322. [Google Scholar]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M. Biocontrol Agent Talaromyces flavus Stimulates the Growth of Cotton and Potato. J. Plant. Growth. Regul. 2012, 31, 471–477. [Google Scholar] [CrossRef]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M.; Afshari-Azad, H. Biological control of Verticillium wilt of greenhouse cucumber by Talaromyces flavus. Phytopathol. Mediterr. 2011, 49, 321–329. [Google Scholar] [CrossRef]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M.; Khaledi, E.M. Biological control of tomato Verticillium disease by Talaromyces flavus. J. Plant Prot. Res. 2010, 50, 360–365. [Google Scholar] [CrossRef]
- Abdel-Rahim, I.R.; Abo-Elyousr, K.A.M. Talaromyces pinophilus strain AUN-1 as a novel mycoparasite of Botrytis cinerea, the pathogen of onion scape and umbel blights. Microbiol. Res. 2018, 212–213, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Khalmuratova, I.; Kim, H.; Nam, Y.J.; Oh, Y.; Jeong, M.J.; Choi, H.R.; You, Y.H.; Choo, Y.S.; Lee, I.J.; Shin, J.H.; et al. Diversity and Plant Growth Promoting Capacity of Endophytic Fungi Associated with Halophytic Plants from the West Coast of Korea. Mycobiology 2015, 43, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Peterson, S.W.; Jurjevic, Z. The Talaromyces pinophilus species complex. Fungal Biol. 2019, 123, 745–762. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Yilmaz, N.; Thrane, U.; Rasmussen, K.B.; Houbraken, J.; Samson, R.A. Talaromyces atroroseus, a new species efficiently producing industrially relevant red pigments. PLoS ONE 2013, 8, e84102. [Google Scholar] [CrossRef] [Green Version]
- Zaccarim, B.R.; de Oliveira, F.; Passarini, M.R.Z.; Duarte, A.W.F.; Sette, L.D.; Jozala, A.F.; Teixeira, M.F.S.; de Carvalho Santos-Ebinuma, V. Sequencing and phylogenetic analyses of Talaromyces amestolkiae from amazon: A producer of natural colorants. Biotechnol. Prog. 2019, 35, e2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Collection Number | Substrate and Origin | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|---|
| ITS | BenA | CaM | RPB1 | RPB2 | |||
| strain JP-NJ4 | M 2012167 | Rhizosphere soil from Pinus massoniana; Nanjing, Jiangsu, China | MW130720 | MW147759 | MW147760 | MW147761 | MW147762 |
| Talaromyces brevis | CBS 141833 (T) = DTO 349-E7 | Soil; Beijing, China | MN864269 | MN863338 | MN863315 | MN863328 | |
| DTO 307-C1 | Soil; Zonguldak, Turkey | MN864270 | MN863339 | MN863316 | MN863329 | ||
| CBS 118436 = DTO 004-D8 | Soil; Maroc | MN864271 | MN863340 | MN863317 | MN863330 | ||
| Talaromyces liani | CBS 225.66 (T) | Soil; China | JN899395 | JX091380 | KJ885257 | JN680280 | KX961277 |
| CBS 118434 | Soil in orchid garden; Sanur, Bali, Indonesia | KM066208 | KM066139 | MK451683= KP453744 | - | - | |
| CBS 118885 | Soil of pepper field; DaeJeon, Korea | KM066210 | KM066138 | - | - | - | |
| NRRL 1009 | Derived from Biourge 368 | MH793030 | MH792902 | MH792966 | - | MH793093 | |
| NRRL 1014 | = 1009 | MH793031 | MH792903 | MH792967 | - | MH793094 | |
| NRRL 1015 | = 1009 | MH793032 | MH792904 | MH792968 | - | MH793095 | |
| NRRL 1019 | USA, Arizona, isol ignotae, KD Butler, 1936. | MH793033 | MH792905 | MH792969 | - | MH793096 | |
| NRRL 3380 | China, isol ex soil, = CBS 225.66 | MH793037 | MH792909 | MH792973 | - | MH793100 | |
| NRRL 28778 | Brazil, isol ex soil, RW Jackson, 1956. | MH793047 | MH792919 | MH792983 | - | MH793110 | |
| NRRL 28834 | India, isol ignotae | MH793048 | MH792920 | MH792984 | - | MH793111 | |
| CMV011D7 | Passiflora edulis; South Africa | - | MK451201 | - | - | - | |
| KUC21412 | Mudflat; South Korea | MN518409 | MN531288 | - | - | - | |
| DTO 058F2 | Heat tretaed corn kernels; the Netherlands | KM066209 | KM066140 | - | - | - | |
| Talaromyces aculeatus | CBS 289.48 (T) = NRRL2129 | Textile; USA | KF741995 | KF741929 | KF741975 = JX140684 = MH792972 | - | KM023271 |
| CBS 282.92 | Soil in secondary forest; Brazil | KF741981 | KF741914 | KF741946 | - | - | |
| CBS 290.65 | Nut; South Africa | KF741982 | KF741915 | KF741948 | - | - | |
| CBS 563.92 | Stem of Dicymbe Altsonii; French Guiana | KF741986 | KF741920 | KF741963 | - | - | |
| CBS 136673 = IBT14255 | Weathering wood stakes; Palmerston North, New Zealand | KF741990 | KF741927 | KF741970 | - | - | |
| Talaromyces adpressus | NRRL 6014 | Peanuts; Unknown | MH793039 | MH792911 | MH792975 | - | MH793102 |
| NRRL 62466 | Peanuts; Unknown | MH793088 | MH792961 | MH793025 | - | MH793152 | |
| CBS 140620 | Indoor air; China | KU866657 | - | - | - | KU867001 | |
| DTO 317-G4 | Indoor air; China | - | KU866844 | KU866741 | - | - | |
| CMV011C5 | Soil; South Africa | MK450741 | MK451191 | MK451673 | - | - | |
| Talaromyces aerugineus | CBS 350.66 (T) | Debris; United Kingdom | AY753346 = NR 147420 | KJ865736 | KJ885285 | JN121657 | JN121502 |
| Talaromyces albobiverticillius | CBS 133440 (T) = Penicillium albobiverticillium isolate 900890701 | Decaying leaves of a broad-leaved tree; Taiwan | HQ605705 = KF114734 | KF114778 | KJ885258 | KF114753 | KM023310 |
| CBS 133441 | Decaying leaves of a broad-leaved tree; Taiwan | KF114733 | KF114777 | - | KF114755 | - | |
| Talaromyces allahabadensis | CBS 453.93 (T) | Cultivated soil; Allahabad, India | KF984873 | KF984614 = JX494298 | KF984768 | JN680309 | KF985006 |
| CBS 178.81 | Crepis zacintha; Alicante, Spain; Type of Penicillium zacinthae | KF984863 | KF984612 | KF984767 | - | KF985004 | |
| CBS 441.89 | Seed groud; Denmark | KF984872 | KF984613 | KF984759 | - | KF985005 | |
| CBS 137397 = DTO245E3 | House dust; Mexico | KF984864 | KF984605 | KF984761 | - | KF984998 | |
| CBS 137399 = DTO267H6 | House dust; Thailand | KF984866 | KF984607 | KF984762 | - | KF984997 | |
| Talaromyces amestolkiae | CBS 132696 (T) = DTO179F5 | House dust; South Africa | JX315660 = NR 120179 | JX315623 | KF741937 = JX315650 | JX315679 | JX315698 |
| DTO179E4 | House dust; South Africa | KJ775706 | KJ775199 | JX140685 | - | - | |
| DTO179F1 | House dust; South Africa | KJ775707 | KJ775200 | JX140686 | - | - | |
| DTO179F6 | House dust; South Africa | KJ775708 | KJ775201 | - | - | - | |
| Talaromyces angelicus | KACC 46611 (T) = CNU 100013 = DTO303E2 | Dried roots of Angelica gigas; Pyeongchang, Korea | KF183638 | KF183640 | KJ885259 | - | KX961275 |
| FMR 15489 | Unknown | LT899791 | LT898316 | LT899773 | - | LT899809 | |
| FMR 15490 | Unknown | LT899792 | LT898317 | LT899774 | - | LT899810 | |
| Talaromyces apiculatus | CBS 312.59 (T) | Soil; Japan | JN899375 = NR 121530 | KF741916 = JX091378 | KF741950 | JN680293 | KM023287 |
| CBS 548.73 | Soil; Suriname | KF741985 | KF741919 | KF741962 | - | - | |
| CBS 101366 | Soil; Hong Kong, China | KF741977 | KF741910 | KF741932 | - | - | |
| Talaromyces argentinensis | NRRL 28750 (T) | Soil; Unknown | MH793045 = NR 165525 | MH792917 | MH792981 | - | MH793108 |
| NRRL 28758 | Soil; Unknown | MH793046 | MH792918 | MH792982 | - | MH793109 | |
| Talaromyces assiutensis | CBS 147.78 (T) | Soil; Egypt | JN899323 | KJ865720 | KJ885260 | JN680275 | KM023305 |
| CBS 645.80 | Gossypium; India; Type of Talaromyces gossypii | JN899334= NR 147423 | KF114802 | - | JN680317 | - | |
| CBS 116554 | Pasteurised canned strawberries; the Netherlands | KM066167 | KM066124 | MK451674 | - | - | |
| CBS 118440 | Soil; Fes, Marocco | KM066168 | KM066125 | MK451675 | - | - | |
| Talaromyces atricola | CBS 255.31 (T) | Unknown | KF984859 | KF984566 | KF984719 | - | KF984948 |
| Talaromyces atroroseus | CBS 133442 (T) | House dust; South Africa | KF114747 = NR 137815 | KF114789 | KJ775418 | KF114763 | KM023288 |
| DTO267I1 | House dust; Thailand | KJ775716 | KJ775209 | - | - | - | |
| DTO270D5 | House dust; Mexico | KJ775734 | KJ775227 | - | - | - | |
| DTO270D6 | House dust; Mexico | KJ775735 | KJ775228 | - | - | - | |
| Talaromyces aurantiacus | CBS 314.59 (T) | Soil; Georgia | JN899380 = NR103681.2 | KF741917 | KF741951 | JN680294 | KX961285 |
| Talaromyces australis | IBT14256 (T) | Unknown | KF741991 = NR 147431 | KF741922 | KF741971 | - | - |
| IBT14254 | Unknown | KF741989 | KF741923 | KF741969 | - | - | |
| MDL18159 | Bronchoscopy; USA | MK601840 | MK626507 | - | MK626517 | - | |
| Talaromyces austrocalifornicus | CBS 644.95 (T) | Soil; California, USA | JN899357 = NR 137079 | KJ865732 | KJ885261 | JN680316 | - |
| Talaromyces bacillisporus | CBS 296.48 (T) | Leaf; New York, USA | JN899329 | AY753368 | KJ885262 | JN121634 | JF417425 |
| CBS 102389 | Sludge of anaerobic pasteurised organic household waste; Sweden | KM066179 | KM066135 | - | - | - | |
| CBS 110774 | Rye bread; the Netherlands | KM066180 | KM066136 | - | - | - | |
| CBS 116927 | Soil; the Netherlands | KM066181 | KM066137 | - | - | - | |
| Talaromyces bohemicus | CBS 545.86 (T) | Peloids for balneological purposes; Czech Republic | JN899400 = NR 137081 | KJ865719 | KJ885286 | JN121699 | JN121532 |
| Talaromyces boninensis | CBS 650.95 (T) | Peloids for balneological purposes; Czech Republic | JN899356 = NR 145157 | KJ865721 | KJ885263 | JN680319 | KM023276 |
| Talaromyces brunneus | CBS 227.60 (T) | Milled rice imported into Japan; Thailand | JN899365 = NR 111688 | KJ865722 = JX494296 | KJ885264 | JN680281 | KM023272 |
| Talaromyces calidicanius | CBS 112002 (T) | Soil; Nantou County, Taiwan | JN899319 = HQ149324 = NR 103665.2 | HQ156944 | KF741934 = JX140688 | JN899305 | KM023311 |
| ACCC:39162 | Luffa; Beijing; China | KY225703 | KY225714 | - | KY225712 | - | |
| ACCC:39164 | Cucumber; Beijing; China | KY225702 | KY225715 | - | KY225711 | - | |
| Talaromyces californicus | NRRL 58168 (T) | Air sample; Unknown | MH793056 = NR 165527 | MH792928 | MH792992 | - | MH793119 |
| NRRL 58177 | Air sample; Unknown | MH793057 | MH792929 | MH792993 | - | MH793120 | |
| NRRL 58207 | Air sample; Unknown | MH793058 | MH792930 | MH792994 | - | MH793121 | |
| NRRL 58221 | Air sample; Unknown | MH793059 | MH792931 | MH792995 | - | MH793122 | |
| NRRL 58661 | Air sample; Unknown | MH793060 | MH792932 | MH792996 | - | MH793123 | |
| Talaromyces cecidicola | CBS 101419 (T) = Penicillium cecidicola strain DAOM 233329 = Penicillium cecidicola isolate KAS504 | Cynipid insect galls on Quercus pacifica twigs; Oregon, USA | AY787844 = MH862736 | FJ753295 | KJ885287 | - | KM023309 |
| Talaromyces cellulolyticus | Y-94 = FERM: BP-5826 | Unknown; A synonym of Talaromyces pinophilus | AB474749 | AB773823 | - | AB856422 | - |
| Talaromyces chloroloma | DAOM 241016 (T) = Penicillium sp. CMV-2008a isolate Pen389 = Penicillium sp. CMV-2008a isolate CV389 | Fynbos soil; Western Cape, South Africa | FJ160273 | GU385736 | KJ885265 | - | KM023304 |
| DTO 180-F4 = Penicillium sp. CMV-2008a isolate CV390 = Penicillium sp. CMV-2008a isolate Pen390 | Fynbos soil; South Africa | FJ160272 | GU385737 | - | - | - | |
| DTO 182-A5 = CV785 = CV0785 | Air sample; Malmesbury, South Africa | JX091485 | JX091597 | JX140689 | - | MK450871 | |
| Talaromyces cinnabarinus | CBS 267.72 (T) | Soil, Japan | JN899376 | AY753377 | KJ885256 | JN121625 | JN121477 |
| CBS 357.72 | Soil, Japan | KM066178 = MH860496 = AY753347 | KM066134 = AY753376 | - | - | - | |
| Talaromyces cnidii | KACC 46617 (T) = DTO 303-E1 = CNU 100149 | Dried roots of Cnidium officinale; Jecheon, Korea | KF183639 | KF183641 | KJ885266 | - | KM023299 |
| DTO 269-H8 | House dust; Thailand | KJ775724 | KJ775217 | KJ775426 | - | - | |
| DTO 270-A4 | House dust; Thailand | KJ775729 | KJ775222 | KJ775430 | - | - | |
| DTO 270-A8 | House dust; Thailand | KJ775730 | KJ775223 | KJ775431 | - | - | |
| DTO 270-B7 | House dust; Thailand | KJ775731 | KJ775224 | KJ775432 | - | - | |
| Talaromyces coalescens | CBS 103.83 (T) | Soil under Pinus sp.; Spain | JN899366 = NR 120008 | JX091390 | KJ885267 | - | KM023277 |
| Talaromyces columbinus | NRRL 58811 (T) | Air; Loisiana, USA | KJ865739 = NR 147433 | KF196843 | KJ885288 | - | KM023270 |
| CBS 137393 = DTO 189-A5 | Chicken feed (Unga); Nairobi, Kenya | KF984794 | KF984659 | KF984671 | - | KF984897 | |
| NRRL 58644 | Air; Maryland, USA | KF196899 | KF196842 | KF196880 | - | KF196987 | |
| NRRL 62680 | Corn grits; Illinois, USA | KF196901 | KF196844 | KF196882 | KF196949 | KF196988 | |
| Talaromyces convolutus | CBS 100537 (T) | Soil; Kathmandu, Nepal | JN899330 = NR 137157 | KF114773 | - | JN121553 | JN121414 |
| Talaromyces dendriticus | CBS 660.80 (T) | Eucalyptus pauciflora leaf litter; New South Wales, Australia | JN899339 | JX091391 | KF741965 | JN121714 | KM023286= JN121547 |
| DAOM 226674 = Penicillium dendriticum isolate KAS849 | Doryanthes excelsa spathes; Mangrove Mountain, New South Wales, Australia | AY787842 | FJ753293 | - | - | - | |
| DAOM 233861 = Penicillium dendriticum isolate KAS1190 | Unindentified insect gall on Eucalyptus leaf; Kalnura, New South Wales, Australia | AY787843 | FJ753294 | - | - | - | |
| DTO 183-G3 = CV2026 | Mite; Struisbaai, South Africa | JX091486 | JX091619 | JX140692 | - | MK450872 | |
| Talaromyces derxii | CBS 412.89 (T) | Cultivated soil; Japan | JN899327 = NR 145152 | JX494306 | KF741959 | JN680306 | KM023282 |
| Talaromycesdiversus | CBS 320.48 (T) | Leather; USA | KJ865740 | KJ865723 | KJ885268 | JN680297 | KM023285 |
| DTO 133-A7 | House dust; Thailand | KJ775701 | KJ775194 | - | - | - | |
| DTO 133-E4 | House dust; Thailand | KJ775702 | KJ775195 | - | - | - | |
| DTO 133-I6 | Lotus tea; produced in Vietnam, imported to the Netherlands | KJ775700 | KJ775193 | - | - | - | |
| DTO 244-E6 | House dust; New Zealand | KJ775712 | KJ775205 | - | - | - | |
| Talaromyces domesticus | NRRL 58121 | Floor swab; Unknown | MH793055 | MH792927 | MH792991 | - | MH793118 |
| NRRL 62132 | Exposed cloth; Unknown | MH793066 | MH792938 | MH793002 | - | MH793129 | |
| Talaromyces duclauxii | CBS 322.48 (T) | Canvas; France | JN899342 = NR 121526 | JX091384 | KF741955 | JN121643 | JN121491 |
| Talaromycesemodensis | CBS 100536 (T) | Soil; Kathmandu, Nepal | JN899337 = NR 137077 | KJ865724 | KJ885269 | JN121552 | JF417445 |
| Talaromyceserythromellis | CBS 644.80 (T) | Soil from creek bank; New South Wales | JN899383 | HQ156945 | KJ885270 | JN680315 | KM023290 |
| Talaromyces euchlorocarpius | PF 1203 (T) = DTO 176I3 = CBM-FA-0942 | Soil; Yokohama, Japan | AB176617 | KJ865733 | KJ885271 | - | KM023303 |
| Talaromyces flavovirens | CBS 102801 (T) | Dead leaves of Quercus ilex; Parque del Retiro, Madrid, Spain | JN899392 | JX091376 | KF741933 | - | KX961283 |
| DAOM236381 | Leaves of Quercus suber; port de la Selva, Girona, Spain | JX013912 | JX091373 | - | - | - | |
| DAOM236382 | Leaves of Quercus suber; Selva de Mar, Girona, Spain | JX013913 | JX091374 | - | - | - | |
| DAOM236383 | Leaves of Quercus suber; Barraca d’en Rabert, Paau, Girona, Spain | JX013914 | JX091377 | - | - | - | |
| DAOM236384 | Leaves of Quercus suber; Xovar, Alt Palacia, Valencia | JX013915 | JX091375 | - | - | - | |
| Talaromyces flavus | CBS 310.38 (T) | Unknown; New Zealand | JN899360 | JX494302 | KF741949 = FJ530982 | JN121639 | JF417426 |
| CBS 437.62 | Compost; Bonn, Germany | KM066202 | KM066156 | - | - | - | |
| Talaromyces francoae | CBS 113134 (T) | Leaf litter; Colombia | NR 154940 | - | - | - | - |
| DTO 056D9 | Leaf litter; Colombia | KX011510 | KX011489 | KX011501 | - | - | |
| Talaromyces funiculosus | CBS 272.86 (T) | Lagenaria vulgaris; India | JN899377 = NR 103678.2 | JX091383 | KF741945 | JN680288 | KM023293 |
| CBS 171.91 | Unknown | KM066193 | KM066162 | MK451679 | - | MK450873 | |
| CBS 883.70 | Unknown; Java | KM066196 | KM066163 | MK451680 | - | MK450874 | |
| CBS 884.70 | Unknown; Java | KM066195 | KM066164 | MK451681 | - | MK450875 | |
| CBS 885.71 | Air; Java, Jakarta | KM066194 | KM066165 | - | - | MK450876 | |
| Talaromyces fuscoviridis | CBS 193.69 (T) | Unknown | KF741979 = NR 153227 | KF741912 | KF741942 | - | - |
| NRRL 66370 | Unknown | MH793092 | MH792965 | MH793029 | - | MH793156 | |
| Talaromyces galapagensis | CBS 751.74 (T) | Shaded soil under Maytenus obovate; Galapagos Islands, Isla, Santa Cruz, Ecuador | JN899358 = NR 147426 | JX091388 = KF114770 | KF741966 | JN680321 | KX961280 |
| NRRL 13068 | Maytenus obovata | MH793042 | MH792914 | MH792978 | - | MH793105 | |
| Talaromyces hachijoensis | IFM 53624 (T) = PF 1174 = CBM-FA-0948 | Soil; Hachijojima, Japan | AB176620 | - | - | - | - |
| Talaromyces helicus | CBS 335.48 (T) | Soil; Sweden | JN899359 = NR 147427 | KJ865725 | KJ885289 | JN680300 | KM023273 |
| CBS 134.67 | Green house soil under Lycopersicon esculentum; Wageningen, the Netherlands | KM066176 | KM066133 | - | - | - | |
| CBS 550.72 | Saline soil; Vallee de la Seille, France | KM066177 = MH860565 | KM066132 | - | - | - | |
| CBS 649.95 = Talaromyces barcinensis | Unknown | JN899349 = MH862547 = NR 137078 | KJ865737 | - | JN680318 | - | |
| CBS 652.66 | Unknown | JN899335 | KJ865738 | - | JN680320 | - | |
| Talaromyces indigoticus | CBS 100534 (T) | Soil; Japan | JN899331 = NR 137076 | JX494308 | KF741931 | JN680323 | KX961278 |
| Talaromyces intermedius | CBS 152.65 (T) | Allauvial pasture and swamp soil; Nottingham, England | JN899332 = NR 145154 | JX091387 | KJ885290 | JN680276 | KX961282 |
| Talaromyces islandicus | CBS 338.48 (T) | Unknown; Cape Town, South Africa | KF984885 | KF984655 = JX494293 | KF984780 | JN121648 | KF985018 = JN121495 |
| CBS 165.81 | spice mixture used in sausage making industry; Spain; Type of Penicillium aurantioflammiferum | KF984883 | KF984653 | KF984778 | - | KF985016 | |
| CBS 394.50 | Kapok fibre; unkown | KF984886 | KF984656 | KF984781 | - | KF985019 | |
| CBS 117284 | Wheat flour; the Netherlands | KF984882 | KF984652 | KF984777 | - | KF985015 | |
| Talaromyces kabodanensis | DI16-149 | Unknown | - | - | LT795598 | - | LT795599 |
| Talaromyces kendrickii | IBT13593 (T) | Unknown | KF741987 = NR 147430 | KF741921 | KF741967 | - | - |
| IBT14128 | Unknown | KF741988 | KF741925 | KF741968 | - | - | |
| CBS 100105 | Unknown | KF741976 | KF741909 | KF741930 | - | - | |
| CBS 133088 | Unknown | KF741978 | KF741911 | KF741939 | - | - | |
| Talaromyces loliensis | CBS 643.80 (T) | Rye grass (Lolium); New Zealand | KF984888 | KF984658 | KF984783 | JN680314 | KF985021 |
| CBS 172.91 | Soil; New Zealand | KF984887 | KF984657 | KF984782 | - | KF985020 | |
| Talaromyces louisianensis | NRRL 35823 (T) | Air sample; Unknown | MH793052= NR 165526 | MH792924 | MH792988 | - | MH793115 |
| NRRL 35826 | Air sample; Unknown | MH793053 | MH792925 | MH792989 | - | MH793116 | |
| NRRL 35928 | Air sample; Unknown | MH793054 | MH792926 | MH792990 | - | MH793117 | |
| Talaromyces macrosporus | CBS 317.63 (T) | Apple juice; Stellenbosch, South Africa | JN899333 = NR 145155 | JX091382 | KF741952 | JN680296 | KM023292 |
| CBS 117.72 | Cotton fabric; USA | KM066188 | KM066148 | - | - | - | |
| CBS 131.87 | Faecal pellet of grasshopper; Malaysia | KM066191 | KM066147 | - | - | - | |
| CBS 353.72 | Tentage; New Guinea | KM066189 | KM066149 | - | - | - | |
| DTO 077-C5 | Pine apple concentrate; the Netherlands | KM066192 | KM066150 | - | - | - | |
| DTO 105-C4 | Unknown | KM066190 | KM066146 | - | - | - | |
| BCC 14364 | Unknown | AY753345 | AY753373 | - | - | - | |
| AS3.6680 | Unknown | - | - | AY678608 | - | - | |
| Talaromyces malicola | NRRL 3724 (T) | Soil under apple tree; Unknown | MH909513= NR 165531 | MH909406 | MH909459 | - | MH909567 |
| Talaromyces marneffei | CBS 388.87 (T) | Bamboo rat (Rhizomys sinensis); Vietnam | JN899344 = NR 103671.2 | JX091389 | KF741958 | JN899298 | KM023283 |
| CBS 108.89 | Human (male); China | KM066187 | KM066157 | - | - | - | |
| CBS 122.89 | Male AIDS patient after travel to Indonesia | KM066183 | KM066161 | - | - | - | |
| CBS 135.94 | Haemoculture; Nonthaburi, Thailand | KM066184 | KM066158 | - | - | - | |
| CBS 549.77 | Man spleen; unknown | KM066185 | KM066159 | - | - | - | |
| CBS 119456 | Male blood; Thailand | KM066186 | KM066160 | - | - | - | |
| Talaromyces mimosinus | CBS 659.80 (T) | Soil from creek bank, New South Wales | JN899338 | KJ865726 | KJ885272 | JN899302 | - |
| NRRL 13069 = NRRL 13609 (BenA) | Unknown | KX946911 | KX946880 | KX946897 | - | KX946926 | |
| Talaromyces minioluteus | CBS 642.68 (T) | Unknown | JN899346 = NR 121527 | KF114799 | KJ885273 | JN121709 | JF417443 |
| CBS 137.84 = Penicillium samsonii strain CBS137.84 | Fruit, damaged by insect; Valladolid, Spain | KM066171 | KF114798 | - | JN680273 | - | |
| CBS 270.35 | Zea mays; Castle Rock, Virginia, USA; Type of Penicillium purpurogenum var. rubrisclerotium | KM066172 | KM066129 | - | JN680287 | - | |
| Talaromyces muroii | CBS 756.96 (T) | Soil; Taiwan | JN899351 = NR 103672.2 | KJ865727 | KJ885274 | JN680322 | KX961276 |
| CBS 261.55 | Clematis; Boskoop, the Netherlands | KM066200 | KM066153 | - | - | - | |
| CBS 283.58 | Jute potato bag, treated with copper oxide ammonia; unknown | KM066197 | KM066151 | - | - | - | |
| CBS 284.58 | Unknown; the Netherlands | KM066199 | KM066152 | - | - | - | |
| CBS 351.61 | Chicken crop; the Netherlands | KM066198 | KM066155 | - | - | - | |
| CBS 889.96 | Dung of sheep; Papua New Guinea | KM066201 | KM066154 | - | - | - | |
| Talaromyces oumae-annae | CBS 138208 (T) = DTO 269-E8 | House dust; South Africa | KJ775720 = NR 147432 | KJ775213 | KJ775425 | - | KX961281 |
| CBS 138207 = DTO 180-B4 | House dust; South Africa | KJ775710 | KJ775203 | KJ775421 | - | - | |
| Talaromyces palmae | CBS 442.88 (T) | Chrysalidocarpus lutescens seed; Wageningen, the Netherlands | JN899396 | HQ156947 | KJ885291 | JN680308 | KM023300 |
| Talaromyces panamensis | CBS 128.89 (T) | Soil; Barro Colorado Island, Panama | JN899362 | HQ156948 = JX091386 | KF741936 = JX140695 | JN899291 | KM023284 |
| Talaromyces paucisporus | PF 1150 (T) = IFM 53616 = CBM-FA-0944 | Soil; Aso-machi, Japan | AB176603 | - | - | - | - |
| Talaromyces piceus= Talaromyces piceae? | CBS 361.48 (T) | Unknown | KF984792 | KF984668 | KF984680 | - | KF984899 |
| CBS 116872 | Production plant; the Netherlands | KF984788 | KF984660 | KF984678 | - | KF984903 | |
| CBS 132063 | Straw used in horse stable; the Netherlands | KF984789 | KF984665 | KF984674 | - | KF984904 | |
| CBS 137363 = DTO58D1 | Pectin; unknown | KF984787 | KF984664 | KF984677 | - | KF984902 | |
| CBS 137377 = DTO178F3 | House dust; South Africa | KF984784 | KF984661 | KF984676 | - | KF984900 | |
| Talaromyces pinophilus | CBS 631.66 (T) | PVC; France | JN899382 = NR 111691 | JX091381 | KF741964 | JN680313 | KM023291 |
| CBS 173.91 | Unknown; USA | KM066206 | KM066141 | - | - | - | |
| CBS 235.94 | Unknown; USA | KM066204 | KM066145 | - | - | - | |
| CBS 269.73 | Unknown; Germany | KM066207 | KM066144 | KM520392= MK451686 | - | - | |
| CBS 440.89 | Zea mays; India | KM066203 | KM066143 | - | - | - | |
| CBS 762.68 | Rhizosphere; India; Type of Penicillium korosum | JN899347 | JX494301 | - | - | - | |
| CBS 101709 | Soil; Japan | KM066205 | KM066142 | KM520391 = MK451685 | - | - | |
| DTO183-I6 = CV2460 | Protea repens infructescense; Struisbaai, South Africa | JX091488 | JX091621 | JX140697 | - | MK450878 | |
| NRRL 1060 | Seed; Unknown | MH909460 | MH909351 | MH909407 | - | MH909514 | |
| NRRL 3503 | Radio set; Unknown | MH909462 | MH909353 | MH909409 | - | MH909516 | |
| NRRL 5200 | Unknown; Type of Penicillium korosum | MH909464 | MH909355 | MH909411 | - | MH909518 | |
| NRRL 13016 | Dung ball; Unknown | MH909466 | MH909357 | MH909413 | - | MH909520 | |
| NRRL 62103 | Canvas cloth; Unknown | MH909482 | MH909373 | MH909429 | - | MH909535 | |
| NRRL 62172 | Wheat; Unknown | MH909492 | MH909383 | MH909439 | - | MH909545 | |
| ATCC 11797 | Unknown | KU729085 | KU896999 | - | - | - | |
| CABI IMI114933 | Unknown; France | KC962105 | KC992266 | - | - | - | |
| Talaromyces pittii | CBS 139.84 (T) | Clay soil under poplar trees; Spain | JN899325 = NR 103667.2 | KJ865728 | KJ885275 | JN680274 | KM023297 |
| Talaromyces pratensis | NRRL 62170 (T) | Unknown | MH793075 = NR 165529 | MH792948 | MH793012 | - | MH793139 |
| NRRL 13548 | Corn; Unknown | MH793044 | MH792916 | MH792980 | - | MH793107 | |
| NRRL 62126 | River water; Unknown | MH793065 | MH792937 | MH793001 | - | MH793128 | |
| Talaromyces primulinus | CBS 321.48 (T) | Unknown; USA | JN899317 = NR 145151 | JX494305 | KF741954 | JN680298 | KM023294 |
| Talaromyces proteolyticus | CBS 303.67 (T) | Granite soil; Ukraine | JN899387 = NR 103685.2 | KJ865729 | KJ885276 | JN680292 | KM023301 |
| Talaromyces pseudostromaticus | CBS 470.70 (T) | Feather of Hylocichla fuscescens; Minnesota, USA | JN899371 | HQ156950 | KJ885277 | JN899300 | KM023298 |
| Talaromyces ptychoconidium | DAOM 241017 (T) = DTO 180-E7 = CV2808 = Penicillium sp. CMV-2008c isolate CV319 = Penicillium sp. CMV-2008c isolate Pen322 | Fynbos soil; Malmesbury, South Africa | FJ160266 | GU385733 | JX140701 | - | KM023278 |
| DTO 180-E9 = Penicillium sp. CMV-2008c isolate Pen319 = Penicillium sp. CMV-2008c isolate CV322 | Fynbos soil; Malmesbury, South Africa | FJ160267 | GU385734 | - | - | MK450879 | |
| DTO 180-F1 = Penicillium sp. CMV-2008c isolate CV323 | Fynbos soil; Malmesbury, South Africa | GQ414762 | GU385735 | - | - | - | |
| Talaromyces purpureus | CBS 475.71 (T) | Soil; France | JN899328 = NR 145153 | GU385739 | KJ885292 | JN121687 | JN121522 |
| Talaromyces purpurogenus | CBS 286.36 (T) | Parasitic on a culture of Aspergillus oryzae; Japan | JN899372 = NR 121529 | JX315639 | KF741947 = JX315655 | JN680271 | JX315709 |
| CBS 184.27 | Soil; Lousiana, USA | JX315665 = MH854924 | JX315637 | JX315658 | JX315684 = JN680270 | - | |
| CBS 122434 | Unknown | JX315663 | JX315640 | JX315659 | JX315682 | - | |
| CBS 132707 = DTO189A1 | Moulded field corn; Wisconsin, USA | JX315661 | JX315638 | JX315642 | JX315680 | - | |
| Talaromyces rademirici | CBS 140.84 (T) | Air under willow tree; Valladolid, Spain | JN899386 = NR 103684.2 | KJ865734 | - | - | KM023302 |
| Talaromyces radicus | CBS 100489 (T) | Root seadling; New South Wales | KF984878 | KF984599 | KF984773 | - | KF985013 |
| CBS 100488 | Wheat root; New South Wales | KF984877 | KF984598 | KF984772 | - | KF985012 | |
| CBS 100490 | Wheat root; New South Wales | KF984879 | KF984600 | KF984774 | - | KF985014 | |
| CBS 137382 = DTO181D5 | Fynbos soil; South Africa | KF984875 | KF984602 | KF984775 | - | KF985009 | |
| DTO181D4 | Fynbos soil; South Africa | KF984880 | KF984601 | KF984770 | - | KF985008 | |
| DTO181D7 | Fynbos soil; South Africa | KF984881 | KF984603 | KF984771 | - | KF985010 | |
| Talaromyces ramulosus | DAOM 241660 (T) = CV2837 = CV113 | Soil; Malmesbury, South Africa | EU795706 | FJ753290 | JX140711 | - | KM023281 |
| DTO 181-E3 = CV314 = CV0314 | Mite; Stellenbosch, South Africa | JX091494 | JX091626 | JX140706 | - | - | |
| DTO 181-F6 = CV394 = CV0394 | Protea repens infructescense; Stellenbosch, South Africa | JX091495 | JX091629 | JX140707 | - | - | |
| DTO 182-A3 = CV735 = CV0735 | Protea repens infructescense; Stellenbosch, South Africa | JX091496 | JX091630 | JX140708 | - | - | |
| DTO 182-A6 = CV787 = CV0787 | Air, Malmesbury; South Africa | JX091497 | JX091631 | JX140709 | - | - | |
| DTO 183-A7 = CV1426 | Protea repens infructescense; Malmesbury, South Africa | JX091493 | JX091632 | JX140710 | - | - | |
| Talaromyces rotundus | CBS 369.48 (T) | Cardboard; Norway | JN899353 | KJ865730 | KJ885278 | - | KM023275 |
| Talaromyces ruber | CBS 132704 (T) = DTO193H6 | Air craft fuel tank; United Kingdom | JX315662 = NR 111780 | JX315629 | KF741938 | JX315681 | JX315700 |
| CBS 196.88 | Unknown | JX315666 = JN899312 | JX315627 | JX315657 | JN680278 = JX315685 | - | |
| CBS 237.93 | Unknown | JX315667 | JX315628 | JX315656 | JX315686 = JN899306 | - | |
| CBS 370.48 | Currency paper; Washington, USA | JX315673 | JX315630 | JX315649 | JX315692 | - | |
| CBS 868.96 | Unknown | JX315677 | JX315631 | JX315643 | JX315696 = JN899309 | - | |
| Talaromyces rubicundus | CBS 342.59 (T) | Soil; Georgia | JN899384 | JX494309 | KF741956 | JN680301 | KM023296 |
| Talaromyces rugulosus | CBS 371.48 (T) | Roating potato tubers (Solanum tuberosum), USA | KF984834 | KF984575 = JX494297 | KF984702 | JN680302 | KF984925 |
| CBS 344.51 | Unknown; Japan; Type of Penicillium echinosporum | KF984858 | KF984574 | KF984701 | - | KF984924 | |
| CBS 137366 = DTO61E8 | Air sample, beer producing factory; Kaulille, Belgium; Type of Penicillium chrysitis | KF984850 | KF984572 | KF984700= JX140720 | - | KF984922 | |
| NRRL 1053 | Unknown | KF984848 | KF984577 | KF984710 | - | KF984945 | |
| NRRL 1073 | decaying twigs; France; Type of Penicillium tardum and Penicillium elongatum | KF984832 | KF984579 | KF984711 | - | KF984927 | |
| Talaromyces ryukyuensis | NHL 2917 (T) = DTO 176-I6 = strain: NHL2917 | Soil; Naha, Japan | AB176628 = NR147414 | - | - | - | - |
| Talaromyces sayulitensis | CBS 138204 (T) = DTO 245-H1 | House dust; Mexico | KJ775713 | KJ775206 | KJ775422 | - | - |
| CBS 138205 = DTO 245-H2 | House dust; Mexico | KJ775714 | KJ775207 | KJ775423 | - | - | |
| CBS 138206 = DTO 245-H3 | House dust; Mexico | KJ775715 | KJ775208 | KJ775424 | - | - | |
| NRRL 1064 | Corn; Unknown | MH793034 | MH792906 | MH792970 | - | MH793097 | |
| NRRL 6420 | Corn; Unknown | MH793041 | MH792913 | MH792977 | - | MH793104 | |
| FMR 15842 | Unknown | - | LT898325 | - | - | - | |
| BEOFB2600m | Unknown; Serbia | MH630050 | MH780060 | - | - | - | |
| BEOFB2601m | Unknown; Serbia | MH630051 | MH780061 | - | - | - | |
| Talaromyces scorteus | CBS 340.34 (T) = NRRL 1129 | Military equipment; Japan | KF984892 = NR153234 = KF196908 | KF984565 = KF196851 | KF984684 = KX946895 | KF196953 | KF984916 = KF196961 |
| CBS 233.60 | Milled Californian rice; Japan; Type of Talaromyces phialosporus | KF984895 | KF984562 = HQ156949 | KF984683 | JN680282 | KF984917 | |
| CBS 499.75 | Unknown; Nigeria | KF984894 | KF984563 | KF984685 | - | KF984918 | |
| CBS 500.75 | Unknown; Sierra Leone | KF984896 | KF984564 | KF984687 | - | KF984919 | |
| DTO 270-A6 | House dust; Thailand | KF984893 | KF984561 | KF984686 | - | KF984915 | |
| Talaromyces siamensis | CBS 475.88 (T) | Forest soil; Thailand | JN899385 = NR 103683.2 | JX091379 | KF741960 | - | KM023279 |
| DTO 269-I3 | House dust; Thailand | KJ775726 | KJ775219 | KJ775428 | - | - | |
| Talaromyces solicola | CBS 133445 (T) = DAOM 241015 = Penicillium sp. CMV-2008d isolate Pen193 = Penicillium sp. CMV-2008d isolate CV191 | Soil; Malmesbury, South Africa | FJ160264 | GU385731 | KJ885279 | - | KM023295 |
| CBS 133446 | Soil; Malmesbury, South Africa | KF114730 | KF114775 | - | - | - | |
| Talaromyces stipitatus | CBS 375.48 (T) | Decaying wood; Louisiana, USA | JN899348 = NR 147424 | KM111288 | KF741957 | JN680303 | KM023280 |
| NBRC 100533 | Unknown | - | AB773824 | - | AB856423 | - | |
| Talaromyces stollii | CBS 408.93 (T) | AIDS patient; the Netherlands | JX315674 = NR 111781 | JX315633 | JX315646 | JX315693 | JX315712 |
| CBS 169.91 | Unknown substrate; South Africa | JX315664 | JX315634 | JX315647 | JX315683 | - | |
| CBS 265.93 | Bronchoalveolar lavage of patient after lung transplantation (subclinical); France | JX315670 | JX315635 | JX315648 | JX315689 | - | |
| CBS 581.94 | Unknown | JX315675 | JX315632 | JX315645 | JX315694 | - | |
| CBS 624.93 | Ananas camosus cultivar; Martinique | JX315676 | JX315636 | JX315644 = JX965209 | JX315695 = JX965281 | JX965315 | |
| NRRL 1768 | USA, Georgia, isol ex peanut, RJ Cole, 1974. | - | - | - | - | MH793098 | |
| NRRL 62122 | Unknown | - | - | - | - | MH793127 | |
| NRRL 62160 | Unknown | - | - | - | - | MH793136 | |
| NRRL 62163 | Unknown | - | - | - | - | MH793137 | |
| NRRL 62165 | Soil; Unknown | - | - | - | - | MH793138 | |
| NRRL 62171 | Unknown | - | - | - | - | MH793140 | |
| NRRL 62227 | Corn; Unknown | - | - | - | - | MH793144 | |
| Talaromyces subinflatus | CBS 652.95 (T) | Copse soil; Japan | JN899397 = NR 137080 | KJ865737 = JX494288 | KJ885280 | JN899301 | KM023308 |
| Talaromyces tardifaciens | CBS 250.94 (T) | Paddy soil; Bhaktapur, Nepal | JN899361 | KC202954 = KF984560 | KF984682 | JN680283 | KF984908 |
| Talaromyces thailandensis | CBS 133147 (T) | Soil; Thailand | JX898041 = NR 147428 | JX494294 | KF741940 | JX898043 | KM023307 |
| Talaromyces trachyspermus | CBS 373.48 (T) | Unknown; USA | JN899354 = NR 147425 | KF114803 | KJ885281 | JN121664 | JF417432 |
| CBS 116556 | Pasteurised canned strawberries; Germany | KM066170 | KM066126 | MK451694 | - | - | |
| CBS 118437 | Soil; Marocco | KM066169 | KM066127 | MK451695 | - | - | |
| CBS 118438 | Soil; Marocco | KM066166 | KM066128 | MK451696 | - | - | |
| Talaromyces tratensis | CBS 113146 (T) = CBS 133146 (RPB1)? | Soil; Trat, Thailand | KF984891 | KF984559 | KF984690 | JX898042 | KF984911 |
| CBS 137400 = DTO 270-F5 | House dust; Mexico | KF984889 | KF984557 | KF984688 | - | KF984909 | |
| CBS 137401 = NRRL1013 | Carbonated beverage; Washington D.C., USA | KF984890 | KF984558 | KF984689 | - | KF984910 | |
| Talaromyces tumuli | NRRL 62151 (T) | Soil; Unknown | MH793071= NR 165528 | MH792944 | MH793008 | - | MH793135 |
| NRRL 6013 | Unknown | MH793038 | MH792910 | MH792974 | - | MH793101 | |
| NRRL 62469 | Peanut; Unknown | MH793089 | MH792962 | MH793026 | - | MH793153 | |
| NRRL 62471 | Peanut; Unknown | MH793090 | MH792963 | MH793027 | - | MH793154 | |
| F-3 | Unknown | MT434004 | - | - | - | - | |
| Talaromyces ucrainicus | CBS 162.67 (T) | Unknown | JN899394 = NR 153205 | KF114771 | KJ885282 | JN680277 | KM023289 |
| CBS 127.64 | soil treated with cyanamide; Germany; Type of Talaromyces ohiensis | KM066173 | KF114772 | - | JN680272 | - | |
| CBS 583.72A | Soil; Japan | KM066174 | KM066130 | - | - | - | |
| CBS 583.72C | Soil; Japan | KM066175 | KM066131 | - | - | - | |
| Talaromyces udagawae | CBS 579.72 (T) | Soil; Misugimura, Japan | JN899350 = NR 145156 | KF114796 | KX961260 | JN680310 | - |
| Talaromyces unicus | CBS 100535 (T) | Soil; Taiwan | JN899336 = NR 157429 | KJ865735 | KJ885283 | JN680324 | - |
| Talaromyces varians | CBS 386.48 (T) | Cotton yarn; England | JN899368 = NR 111689 | KJ865731 | KJ885284 | JN680305 | KM023274 |
| Talaromyces veerkampii | CBS 500.78 (T) | Unknown | KF741984 = NR 153228 | KF741918 | KF741961 | - | KX961279 |
| NRRL 6095 | Unknown | MH793040 | MH792912 | MH792976 | - | MH793103 | |
| NRRL 62286 | Wheat flour; Unknown | MH793085 | MH792958 | MH793022 | - | MH793149 | |
| IBT18366 | Unknown | KF741993 | KF741924 | KF741973 | - | - | |
| CMV005D6 | Soil; South Africa | MK450751 | MK451043 | - | - | - | |
| Talaromyces verruculosus | NRRL 1050 (T) = CBS 388.48 | Soil; Texas, USA | KF741994 | KF741928 | KF741974 | - | KM023306 |
| CBS 254.56 | Unknown; Yangambi, Zaire | KF741980 | KF741913 | KF741944 | - | - | |
| DTO 129-H4 | House dust; Thailand | KJ775698 | KJ775191 | KJ775419 | - | - | |
| DTO 129-H5 | House dust; Thailand | KJ775699 | KJ775192 | KJ775420 | - | - | |
| AX2101 I | Metallic surface; Para, Brazil | KJ413368 | KJ413340 | - | - | KJ476428 | |
| Talaromycesviridis | CBS 114.72 (T) = Sagenoma viride | Soil; Australia | AF285782 = MH860406 = NR160136 | JX494310 | KF741935 | JN121571 | JN121430 |
| Talaromyces viridulus | CBS 252.87 (T) | Soil from bank of creek floading into Little river; New South Wales | JN899314 = NR103663.2 | JX091385 | KF741943 | JN680284 = JN121620 | JF417422 |
| Talaromyces wortmannii | CBS 391.48 (T) | Soil; Denmark | KF984829 | KF984648 | KF984756 | JN121669 | KF984977 = JF417433 |
| CBS 319.63 | Unknown | KF984828 | KF984651 | KF984755 | - | KF984961 | |
| CBS 385.48 = NRRL 1048 | coconut matting; Johannesburg, South Africa; Type of Talaromyces variabilis | KF196915 | KF196853 = JX494295 | KF196878 | JN680304 | KF196975 = KX657552 | |
| CBS 895.73 | Unkown; Japan | KF984811 | KF984626 | KF984737 | - | KF984982 | |
| CBS 137376 = DTO 176-I7 | soil; Japan; Type of Talaromyces sublevisporus | KF984800 | KF984632 | KF984724 | - | KF984979 | |
| NRRL 2125 = DTO 278-E7 | Weathering canvas; Panama | KF984797 | KF984635 | KF984731 | - | KF984991 | |
| Talaromyces xishaensis | HMAS 248732 (T) | China | NR147445 | - | - | - | - |
| - | China | KU644580 | KU644581 | KU644582 | - | - | |
| Talaromyces yelensis | CBS 138210 (T) = DTO 268-E5 | House dust; Micronesia | KJ775717 | KJ775210 | KP119162 | - | KP119164 |
| CBS 138209 = DTO 268-E7 | House dust; Micronesia | KJ775719 = NR 145183 | KJ775212 | KP119161 | - | KP119163 | |
| Morphological Characters | Species | ||||
|---|---|---|---|---|---|
| T. liani (Yilmaz et al., 2014) | Talaromyces Strain JP-NJ4 | T. brevis (Sun et al., 2020) | |||
| Macromorphological Characters | Ascomata | Present after 25 °C, 7 d on OA and MEA (at 30 °C abundant yellow ascomata) | Present after 25 °C, 7 d on OA, 25 °C, 14 d on CZ, and 30 °C, 14 d on CYA and MEA | Present after 25 °C, 7 d on OA | |
| Growth rate (mm) Diam (diameter), 7 d | CZ (25 °C) | Unknown | 29–33 | Unknown | |
| CYA (25 °C) | 20–30 | 25–29 | 30–31 | ||
| CYA (30 °C) | 25–37 | 30–37 | 28–30 | ||
| CYA (37 °C) | 20–25 | 21–31 | 25–26 | ||
| MEA (25 °C) | 35–45 | 31–33 | 50–51 | ||
| MEA (30 °C) | 50–55 | 35–41 | 57–60 | ||
| MEAbl (25 °C) | Unknown | 34–43 | Unknown | ||
| OA (25 °C) | 35–40 | 38–44 | 39–43 | ||
| DG18 (25 °C) | 10–17 | 15–18 | 13–15 | ||
| CYAS (25 °C) | No growth | No growth | No growth | ||
| YES (25 °C) | 35–40 | 30–40 | 42–43 | ||
| CREA (25 °C) | 10–20 | 18–24 | 13–14 | ||
| HAY (25 °C) | Unknown | No growth | Unknown | ||
| Colour of CYA reverse | Light orange and light yellow (5A5–4A5) | Centre pastel yellow (2D4) to pale yellow (1A4) | Ochreous (44) | ||
| Soluble pigment | Absent on CYA (in some isolates yellow) and MEA at 25 °C, 7 d | Weak yellow and orange soluble pigments present on CYA and MEA at 25 °C, 7 d; Strong red soluble pigments present on MEA at 25 °C, 14 d | Absent | ||
| MEA colony texture | Velvety and floccose | Velvety to floccose | Floccose | ||
| Acid production on CREA | Absent (in some isolates very weak) | Present strong | Present | ||
| Micromorphological Characters | Conidiophore | Present | Present | Present | |
| Conidiophore branching | Mono- to biverticillate | Monoverticillate to biverticillate, reduced conidiophores consisting of solitary phialides | Mono- to biverticillate | ||
| Conidium | Shape | Ellipsoidal | Globose to subglobose; (sometimes ovoid) | Subglobose to fusiform | |
| Size (μm) | 2.5–4(–4.5) × 2–3.5 | 2–3 × 3; (3 × 3–3.5) | 3–4(–5) × 2.5–3.5(–4.5) | ||
| Ornamentation | Smooth | Smooth | Smooth | ||
| Ascoma colour | Yellow to orange red | Yellow | Yellow to orange | ||
| Ascoma shape | Globose to subglobose | Globose to subglobose | Globose to subglobose | ||
| Ascoma size (μm) | 150–550 × 150–545 | 300–950 × 300–1000 | 400–550 × 400–550 | ||
| Asci size (μm) | 9–13 × 7.5–11 | 10–12 × 8–10 | Unknown | ||
| Ascospore | Shape | Broadly ellipsoidal | Broadly ellipsoidal | Ellipsoidal | |
| Size (μm) | 4–6 × 2.5–4 | 3.5–5 × 2–3 | 3.5–4.5 × 3–4 | ||
| Ridges | Absent | Absent | Absent | ||
| Ornamentation | Spiny | Spiny | Spiny | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.-R.; Xu, M.-Y.; Kong, W.-L.; Wu, F.; Zhang, Y.; Xie, X.-L.; Li, D.-W.; Wu, X.-Q. Fine Identification and Classification of a Novel Beneficial Talaromyces Fungal Species from Masson Pine Rhizosphere Soil. J. Fungi 2022, 8, 155. https://doi.org/10.3390/jof8020155
Sun X-R, Xu M-Y, Kong W-L, Wu F, Zhang Y, Xie X-L, Li D-W, Wu X-Q. Fine Identification and Classification of a Novel Beneficial Talaromyces Fungal Species from Masson Pine Rhizosphere Soil. Journal of Fungi. 2022; 8(2):155. https://doi.org/10.3390/jof8020155
Chicago/Turabian StyleSun, Xiao-Rui, Ming-Ye Xu, Wei-Liang Kong, Fei Wu, Yu Zhang, Xing-Li Xie, De-Wei Li, and Xiao-Qin Wu. 2022. "Fine Identification and Classification of a Novel Beneficial Talaromyces Fungal Species from Masson Pine Rhizosphere Soil" Journal of Fungi 8, no. 2: 155. https://doi.org/10.3390/jof8020155