
Quantitative PCR Effectively Quantifies Triazole-Susceptible and Triazole-Resistant Aspergillus fumigatus in Mixed Infections

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Cultures, and Sample Preparation
2.2. DNA Extraction
2.3. qPCR Reaction and Calculations
2.4. Galleria Mellonella Infection
2.5. Statistical Analysis
3. Results
3.1. Increased DNA Extraction Yield through Optimized Sample Homogenization Methodology
3.2. Selection of Specific Primers for the Mutated TR34-bp Insertion in A. fumigatus
3.3. Mixed Triazole-Susceptible and Resistant A. fumigatus Detection and Ratio Quantification
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Resendiz-Sharpe, A.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E.; ISHAM/ECMM Aspergillus Resistance Surveillance Working Group. Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56, S83–S92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resendiz-Sharpe, A.; Mercier, T.; Lestrade, P.P.A.; Van Der Beek, M.T.; Borne, P.A.V.D.; Cornelissen, J.J.; De Kort, E.; Rijnders, B.; Schauwvlieghe, A.F.A.D.; Verweij, P.E.; et al. Prevalence of voriconazole-resistant invasive aspergillosis and its impact on mortality in haematology patients. J. Antimicrob. Chemother. 2019, 74, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Lestrade, P.P.; Bentvelsen, R.; Schauwvlieghe, A.F.A.D.; Schalekamp, S.; Van Der Velden, W.J.F.M.; Kuiper, E.J.; Van Paassen, J.; Van Der Hoven, B.; Van Der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole-resistant aspergillosis—Voriconazole resistance and mortality in invasive aspergillosis: A multicenter retrospective cohort study. Clin. Infect. Dis. 2018, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nywening, A.V.; Rybak, J.M.; Rogers, P.D.; Fortwendel, J.R. Mechanisms of triazole resistance in Aspergillus fumigatus. Environ. Microbiol. 2020, 22, 4934–4952. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Carvalho, A.; Houbraken, J.; Lombardi, L.; Garcia-Rubio, R.; Jenks, J.; Rivero-Menendez, O.; Aljohani, R.; Jacobsen, I.; Berman, J.; et al. Aspergillus fumigatus and aspergillosis: From basics to clinics. Stud. Mycol. 2021, 100, 100115. [Google Scholar] [CrossRef]
- Herbrecht, R.; Denning, D.W.; Patterson, T.F.; Bennett, J.E.; Greene, R.E.; Oestmann, J.-W.; Kern, W.V.; Marr, K.A.; Ribaud, P.; Lortholary, O.; et al. Voriconazole versus amphotericin B for primary therapy of invasive aspergillosis. N. Engl. J. Med. 2002, 347, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Schauwvlieghe, A.F.A.D.; Vonk, A.G.; Buddingh, E.P.; Hoek, R.A.S.; Dalm, V.A.; Klaassen, C.H.W.; Rijnders, B. Detection of azole-susceptible and azole-resistant Aspergillus coinfection by cyp51A PCR amplicon melting curve analysis. J. Antimicrob. Chemother. 2017, 72, 3047–3050. [Google Scholar] [CrossRef] [Green Version]
- Chong, G.M.; Van Der Beek, M.T.; von dem Borne, P.A.; Boelens, J.; Steel, E.; Kampinga, G.A.; Span, L.; Lagrou, K.; Maertens, J.; Dingemans, G.J.H.; et al. PCR-based detection of Aspergillus fumigatus Cyp51A mutations on bronchoalveolar lavage: A multicentre validation of the AsperGenius assay ® in 201 patients with haematological disease suspected for invasive aspergillosis. J. Antimicrob. Chemother. 2016, 71, 3528–3535. [Google Scholar] [CrossRef] [Green Version]
- Kolwijck, E.; van der Hoeven, H.; de Sévaux, R.G.L.; Oever, J.T.; Rijstenberg, L.L.; van der Lee, H.A.L.; Zoll, J.; Melchers, W.J.G.; Verweij, P.E. Voriconazole-susceptible and voriconazole-resistant Aspergillus fumigatus coinfection. Am. J. Respir. Crit. Care Med. 2016, 193, 927–929. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, B.; Mahto, K.K.; Meis, J.F.; Chowdhary, A. High-frequency direct detection of triazole resistance in Aspergillus fumigatus from patients with chronic pulmonary fungal diseases in India. J. Fungi 2020, 6, 67. [Google Scholar] [CrossRef]
- Resendiz-Sharpe, A.; Merckx, R.; Verweij, P.E.; Maertens, J.; Lagrou, K. Stable prevalence of triazole-resistance in Aspergillus fumigatus complex clinical isolates in a Belgian tertiary care center from 2016 to 2020. J. Infect. Chemother. 2021, 27, 1774–1778. [Google Scholar] [CrossRef]
- Lackner, M.; Rambach, G.; Jukic, E.; Sartori, B.; Fritz, J.; Seger, C.; Hagleitner, M.; Speth, C.; Lass-Flörl, C. Azole-resistant and-susceptible Aspergillus fumigatus isolates show comparable fitness and azole treatment outcome in immunocompetent mice. Med. Mycol. 2017, 56, 703–710. [Google Scholar] [CrossRef]
- Valsecchi, I.; Mellado, E.; Beau, R.; Raj, S.; Latgé, J.-P. Fitness studies of azole-resistant strains of Aspergillus fumigatus. Antimicrob. Agents Chemother. 2015, 59, 7866–7869. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Mouton, J.W.; Melchers, W.J.G.; Verweij, P.E. Posaconazole prophylaxis in experimental azole-resistant invasive pulmonary aspergillosis. Antimicrob. Agents Chemother. 2015, 59, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Lepak, A.J.; Marchillo, K.; VanHecker, J.; Andes, D.R. Posaconazole pharmacodynamic target determination against wild-type and Cyp51 mutant isolates of Aspergillus fumigatus in an in vivo model of invasive pulmonary aspergillosis. Antimicrob. Agents Chemother. 2013, 57, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Lepak, A.J.; Marchillo, K.; VanHecker, J.; Andes, D.R. Isavuconazole (BAL4815) pharmacodynamic target determination in an in vivo murine model of invasive pulmonary aspergillosis against wild-type and cyp51 mutant isolates of Aspergillus fumigatus. Antimicrob. Agents Chemother. 2013, 57, 6284–6289. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.A.; Bakken, L.R. Viability of soil bacteria: Optimization of plate-counting technique and comparison between total counts and plate counts within different size groups. Microb. Ecol. 1987, 13, 59–74. [Google Scholar] [CrossRef]
- Bowman, J.C.; Abruzzo, G.K.; Anderson, J.W.; Flattery, A.M.; Gill, C.J.; Pikounis, V.B.; Schmatz, D.M.; Liberator, P.A.; Douglas, C.M. Quantitative PCR assay to measure Aspergillus fumigatus burden in a murine model of disseminated aspergillosis: Demonstration of efficacy of caspofungin acetate. Antimicrob. Agents Chemother. 2001, 45, 3474–3481. [Google Scholar] [CrossRef] [Green Version]
- Hartley, M.G.; Ralph, E.; Norville, I.H.; Prior, J.L.; Atkins, T.P. Comparison of PCR and viable count as a method for enumeration of bacteria in an A/j mouse aerosol model of Q Fever. Front. Microbiol. 2019, 10, 1552. [Google Scholar] [CrossRef] [Green Version]
- Vallor, A.C.; Kirkpatrick, W.R.; Najvar, L.K.; Bocanegra, R.; Kinney, M.C.; Fothergill, A.W.; Herrera, M.L.; Wickes, B.L.; Graybill, J.R.; Patterson, T.F. Assessment of Aspergillus fumigatus burden in pulmonary tissue of guinea pigs by quantitative PCR, galactomannan enzyme immunoassay, and quantitative culture. Antimicrob. Agents Chemother. 2008, 52, 2593–2598. [Google Scholar] [CrossRef]
- Herrera, M.L.; Vallor, A.C.; Gelfond, J.A.; Patterson, T.F.; Wickes, B.L. Strain-dependent variation in 18S ribosomal DNA copy numbers in Aspergillus fumigatus. J. Clin. Microbiol. 2009, 47, 1325–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Imai, J.; Clemons, K.V.; Stevens, D.A. Efficacy of caspofungin against central nervous system Aspergillus fumigatus infection in mice determined by TaqMan PCR and CFU methods. Antimicrob. Agents Chemother. 2005, 49, 1369–1376. [Google Scholar] [CrossRef] [Green Version]
- Resendiz-Sharpe, A.; da Silva, R.P.; Geib, E.; Vanderbeke, L.; Seldeslachts, L.; Hupko, C.; Brock, M.; Lagrou, K.; Velde, G.V. Longitudinal multimodal imaging-compatible mouse model of triazole-sensitive and -resistant invasive pulmonary aspergillosis. Dis. Model. Mech. 2022, 15, 049165. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, L.J.; Anyim, M.; Doffman, S.R.; Wilks, M.; Millar, M.R.; Agrawal, S.G. Comparison of DNA extraction methods for Aspergillus fumigatus using real-time PCR. J. Med Microbiol. 2006, 55, 1187–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Linden, J.W.M.; Snelders, E.; Arends, J.P.; Daenen, S.M.; Melchers, W.J.G.; Verweij, P.E. Rapid diagnosis of azole-resistant aspergillosis by direct PCR using tissue specimens. J. Clin. Microbiol. 2010, 48, 1478–1480. [Google Scholar] [CrossRef] [Green Version]
- Snelders, E.; Van Der Lee, H.A.L.; Kuijpers, J.; Rijs, A.J.M.M.; Varga, J.; Samson, R.A.; Mellado, E.; Donders, A.R.T.; Melchers, W.; Verweij, P.E. Emergence of azole resistance in Aspergillus fumigatus and spread of a single resistance mechanism. PLoS Med. 2008, 5, e219. [Google Scholar] [CrossRef]
- Kim, W.-B.; Park, C.; Cho, S.-Y.; Chun, H.-S.; Lee, D.-G. Development of multiplex real-time PCR for rapid identification and quantitative analysis of Aspergillus species. PLoS ONE 2020, 15, e0229561. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, F.; Hashemi, S.J.; Zoll, J.; Melchers, W.J.G.; Rafati, H.; Dehghan, P.; Rezaie, S.; Tolooe, A.; Tamadon, Y.; van der Lee, H.A.; et al. Quantitative analysis of single-nucleotide polymorphism for rapid detection of TR34/L98H- and TR46/Y121F/T289A-positive Aspergillus fumigatus isolates obtained from patients in Iran from 2010 to 2014. Antimicrob. Agents Chemother. 2016, 60, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Martínez, L.; Gil, H.; Rivero-Menéndez, O.; Gago, S.; Cuenca-Estrella, M.; Mellado, E.; Alastruey-Izquierdo, A. Development and validation of a high-resolution melting assay to detect azole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2017, 61, e01083-17. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.W.; Kellogg, C.A.; Peak, K.K.; Shinn, E. A rapid and efficient assay for extracting DNA from fungi. Lett. Appl. Microbiol. 2002, 34, 210–214. [Google Scholar] [CrossRef]
- Jones, A.; Torkel, C.; Stanley, D.; Nasim, J.; Borevitz, J.; Schwessinger, B. High-molecular weight DNA extraction, clean-up and size selection for long-read sequencing. PLoS ONE 2021, 16, e0253830. [Google Scholar] [CrossRef]
- Goldschmidt, P.; Degorge, S.; Merabet, L.; Chaumeil, C. Enzymatic treatment of specimens before DNA extraction directly influences molecular detection of infectious agents. PLoS ONE 2014, 9, e94886. [Google Scholar] [CrossRef] [Green Version]
- Qasim, M.A.; Taha, M. Investigation of the mechanism of protein denaturation by guanidine hydrochloride-induced dissociation of inhibitor-protease complexes. Protein Pept. Lett. 2012, 20, 187–191. [Google Scholar] [CrossRef]
- Diaz-Guerra, T.M.; Mellado, E.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. A point mutation in the 14alpha-sterol demethylase gene cyp51A contributes to itraconazole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2003, 47, 1120–1124. [Google Scholar] [CrossRef]




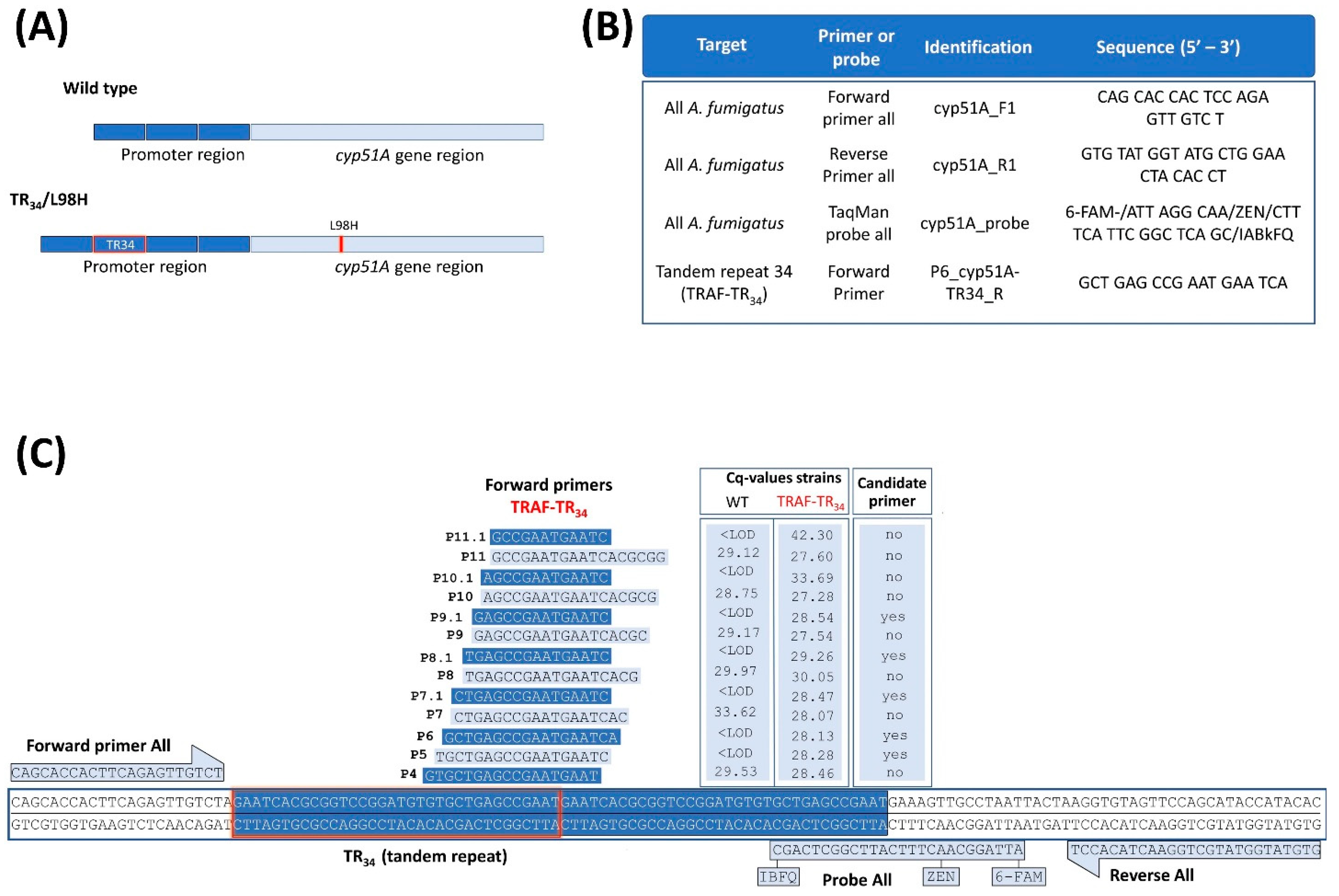{kind=link}
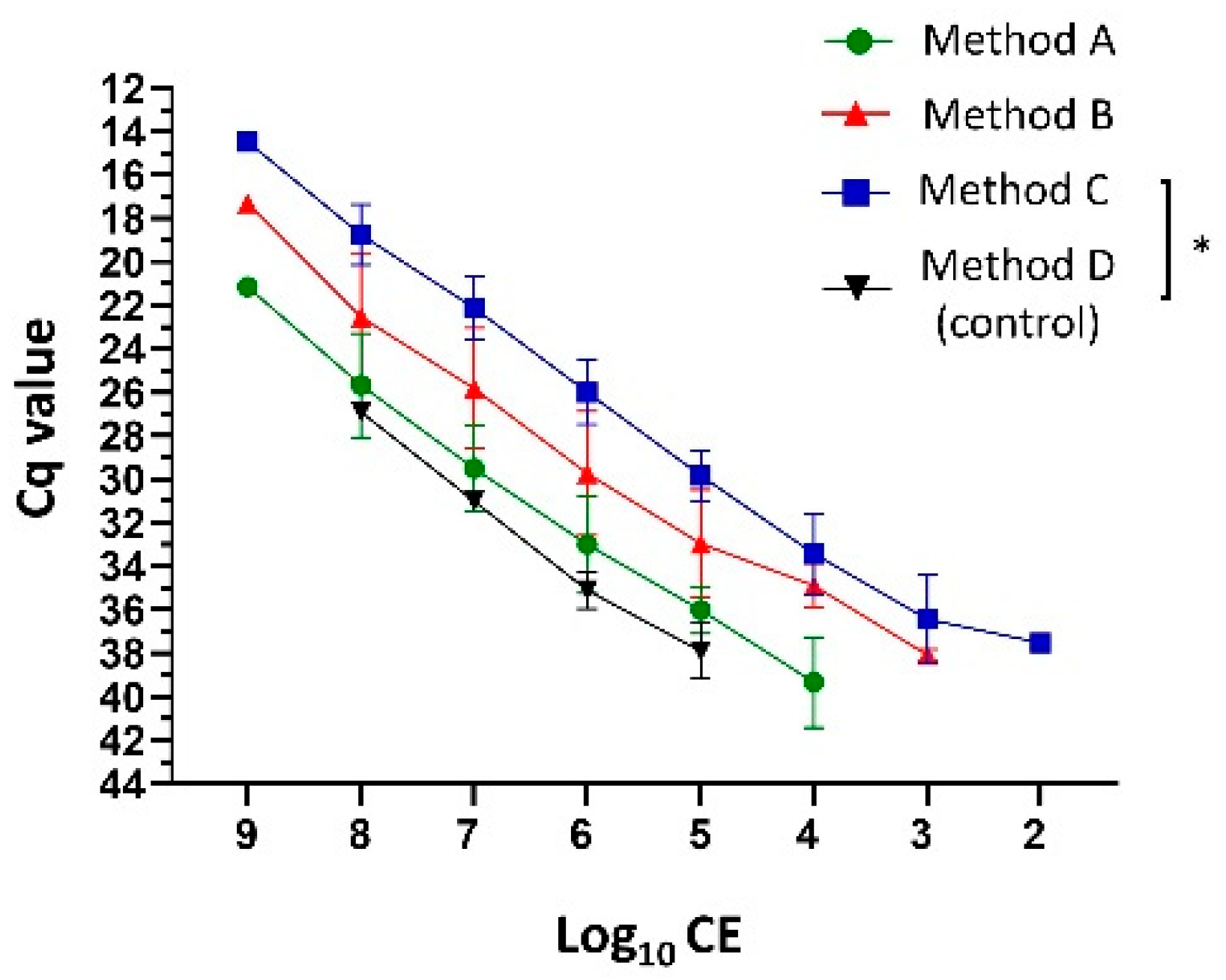{kind=link}
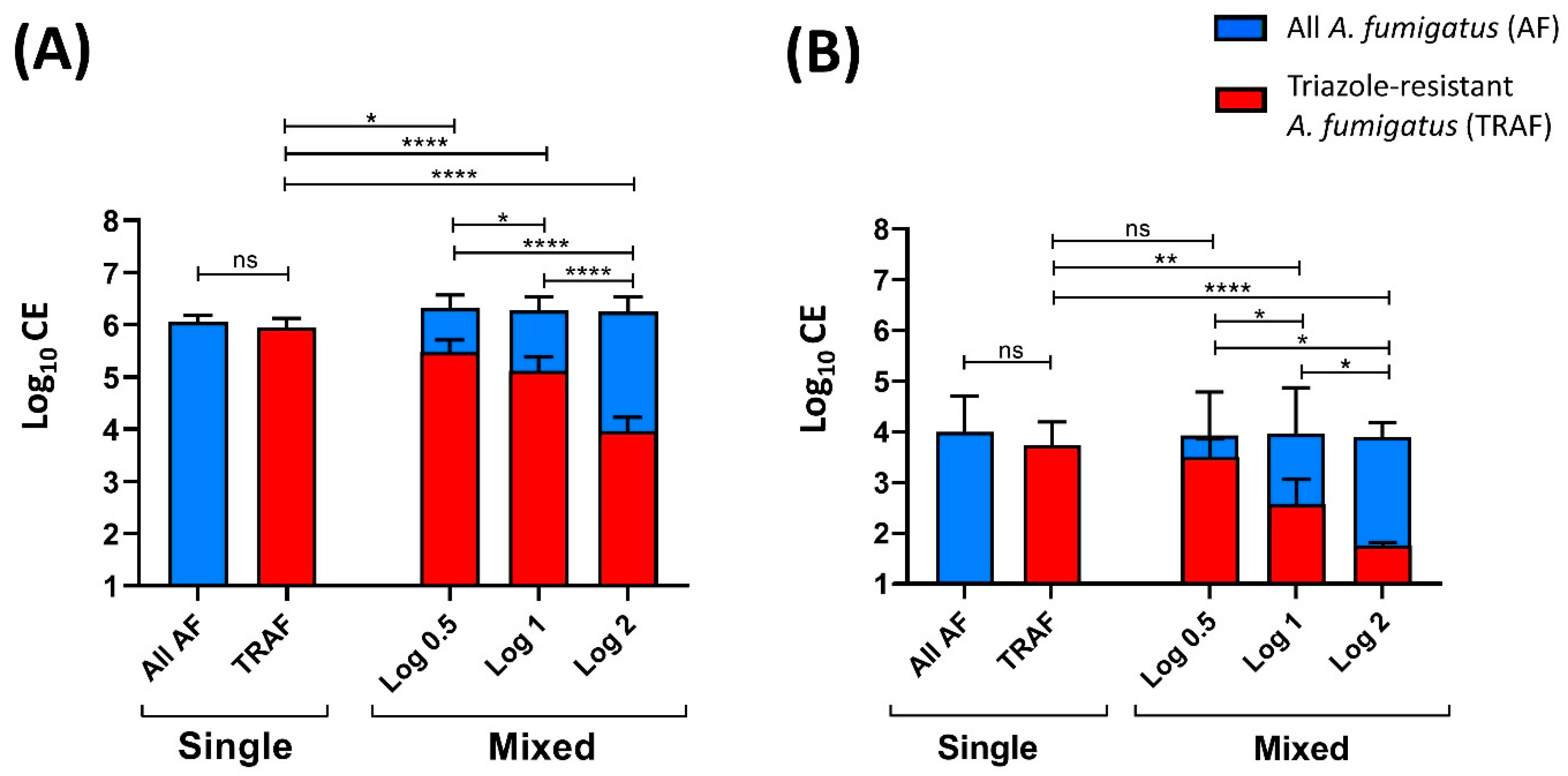{kind=link}
| All A. fumigatus Primers | TRAF-TR34 A. fumigatus Primers | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample Phenotype | Conidia in Sample | Cq Average ± SD | Cq (% CV) | Average Log10 (x/mL) ± SD | Conidia Equivalent (No.) | Cq Average ± SD | Cq (% CV) | Average Log10 (x/mL) ± SD | Conidia Equivalent (No.) |
| WT | 1 × 108 | 21.91 ± 0.15 | 0.66 | 7.78 ± 0.14 | 6.87 × 107 | ND | ND | ND | ND |
| WT | 1 × 107 | 24.35 ± 0.11 | 0.46 | 7.07 ± 0.04 | 1.20 × 107 | ND | ND | ND | ND |
| WT | 1 × 106 | 27.68 ± 0.09 | 0.32 | 6.07 ± 0.03 | 1.20 × 106 | ND | ND | ND | ND |
| WT | 1 × 105 | 31.59 ± 0.15 | 0.47 | 4.90 ± 0.05 | 7.87 × 104 | ND | ND | ND | ND |
| WT | 1 × 104 | 34.91 ± 0.30 | 0.85 | 3.90 ± 0.09 | 7.98 × 103 | ND | ND | ND | ND |
| WT | 1 × 103 | 37.66 ± 0.35 | 0.92 | 3.07 ± 0.10 | 1.18 × 103 | ND | ND | ND | ND |
| WT | 1 × 102 | ND | ND | ND | ND | ND | ND | ND | ND |
| TRAF-TR34 | 1 × 108 | 21.72 ± 0.16 | 0.73 | 7.83 ± 0.05 | 7.14 × 107 | 22.02 ± 0.17 | 0.79 | 7.74 ± 0.07 | 5.68 × 107 |
| TRAF-TR34 | 1 × 107 | 24.02 ± 0.05 | 0.22 | 7.14 ± 0.02 | 1.42 × 107 | 24.27 ± 0.10 | 0.43 | 7.07 ± 0.02 | 1.19 × 107 |
| TRAF-TR34 | 1 × 106 | 27.92 ± 0.16 | 0.59 | 5.90 ± 0.05 | 8.82 × 105 | 27.30 ± 0.15 | 0.55 | 5.88 ± 0.08 | 8.17 × 105 |
| TRAF-TR34 | 1 × 105 | 31.62 ± 0.14 | 0.45 | 4.75 ± 0.11 | 6.01 × 104 | 31.45 ± 0.34 | 0.97 | 4.86 ± 0.04 | 8.45 × 104 |
| TRAF-TR34 | 1 × 104 | 34.55 ± 0.34 | 0.97 | 3.84 ± 0.11 | 6.89 × 103 | 34.30 ± 0.23 | 0.67 | 4.14 ± 0.07 | 2.84 × 104 |
| TRAF-TR34 | 1 × 103 | 37.34 ± 0.37 | 0.98 | 2.97 ± 0.11 | 9.33 × 102 | 36.88 ± 0.39 | 0.77 | 3.25 ± 0.18 | 2.83 × 103 |
| TRAF-TR34 | 1 × 102 | 38.90 ± 0.36 | 0.93 | 2.48 ± 0.12 | 3.05 × 102 | 38.27 ± 0.36 | 0.93 | 2.68 ± 0.12 | 4.83 × 102 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resendiz-Sharpe, A.; Van Holm, W.; Merckx, R.; Pauwels, M.; Teughels, W.; Lagrou, K.; Vande Velde, G. Quantitative PCR Effectively Quantifies Triazole-Susceptible and Triazole-Resistant Aspergillus fumigatus in Mixed Infections. J. Fungi 2022, 8, 1120. https://doi.org/10.3390/jof8111120
Resendiz-Sharpe A, Van Holm W, Merckx R, Pauwels M, Teughels W, Lagrou K, Vande Velde G. Quantitative PCR Effectively Quantifies Triazole-Susceptible and Triazole-Resistant Aspergillus fumigatus in Mixed Infections. Journal of Fungi. 2022; 8(11):1120. https://doi.org/10.3390/jof8111120
Chicago/Turabian StyleResendiz-Sharpe, Agustin, Wannes Van Holm, Rita Merckx, Martine Pauwels, Wim Teughels, Katrien Lagrou, and Greetje Vande Velde. 2022. "Quantitative PCR Effectively Quantifies Triazole-Susceptible and Triazole-Resistant Aspergillus fumigatus in Mixed Infections" Journal of Fungi 8, no. 11: 1120. https://doi.org/10.3390/jof8111120