




Living Fungi in an Opencast Limestone Mine: Who Are They and What Can They Do?

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Physicochemical Properties
2.2. Sample Collection
2.3. Identification of Living Fungal Community by Amplicon Sequencing of Enrichment Culture
2.3.1. Enrichment of Soil Fungi
2.3.2. DNA Extraction and Amplicon Sequencing
2.3.3. Sequencing Data Analysis
2.3.4. Functional Prediction
2.4. Statistical Analysis and Co-Occurrence Network Analysis
3. Results
3.1. General Infomation: An Overview of Sequencing Analysis
3.2. Living Fungal Community Composition: The Distribution of Living Fungi in Mining and Rehabilitaion Plots as Compared to Forest
3.3. Predictive Functional Profiles: Fungal Traits and Enzyme Activities Detected in Mining and Rehabilitaion Plots as Compared to Forest
3.4. Fungi in Harsh Condition of Limestone Quarry: Taxonomic Distribution and Co-Occorence Network of Living Fungi in the Mining and Rehabilitatoin Plots
3.4.1. Living Fungi in Mining and Rehabilitation Plots
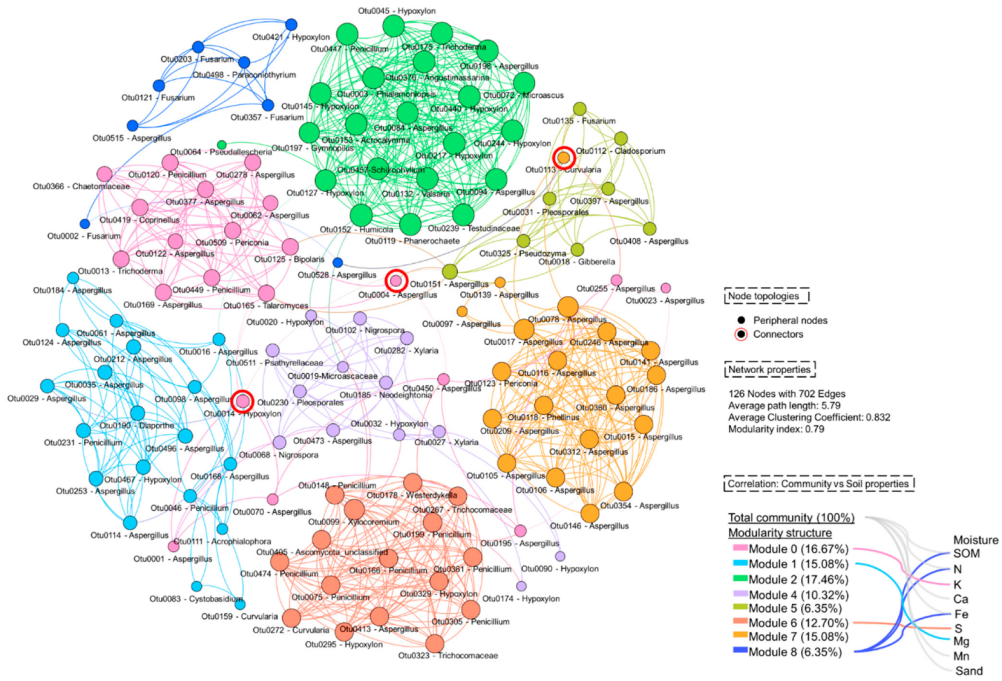
3.4.2. Co-Occurrence Network and Taxonomic Distribution of Living Fungi in Mining and Rehabilitation Plots
4. Discussion
4.1. Living Fungi and Early Mine Rehabilitation: Community Structure, Diverity and Their Associated Function on Early Rehabilitaion Plot as Compared to Mining and Forest Sites
4.2. Living Fungi in Harsh Condition of the Limestone Quarry: There Are Many Way to Survive
4.3. Co-Occorence Network of Living Fungi in Harsh Condition of The Limestone Quarry: The Modular Community and Their Correlation with Soil Physiochemical Properties
4.4. Posible Interaction of Fungi and Plant: Opportunity for Rehabilitation Procedure
4.5. Overall Living Fungal Community Assessment: Limitation and Recommendation for Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuter, N. Reclamation of Degraded Landscapes Due to Opencast Mining; IntechOpen: London, UK, 2013; ISBN 978-953-51-1167-2. [Google Scholar]
- Sonter, L.J.; Ali, S.H.; Watson, J.E.M. Mining and biodiversity: Key issues and research needs in conservation science. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181926. [Google Scholar] [CrossRef]
- Ganapathi, H.; Phukan, M. Environmental Hazards of Limestone Mining and Adaptive Practices for Environment Management Plan. In Environmental Processes and Management; Springer: Cham, Switzerland, 2020; pp. 121–134. ISBN 978-3-030-38151-6. [Google Scholar]
- Bell, R.W. Restoration of Degraded Landscapes: Principles and Lessons from Case Studies with Salt-Affected Land and Mine Revegetation. Chiang Mai Univ. J. 2002, 1, 1–21. [Google Scholar]
- Martins-Loução, M.A.; Correia, O.; Carolino, M.; Correia, A.I.; Afonso, A.; Clemente, A.; Máguas, C. Quarry Rehabilitation: A Case Study. In Advances in Ecological Sciences; WIT Press: Southampton, UK, 2001; Volume 10, pp. 331–346. [Google Scholar]
- Sansupa, C.; Purahong, W.; Wubet, T.; Tiansawat, P.; Pathom-Aree, W.; Teaumroong, N.; Chantawannakul, P.; Buscot, F.; Elliott, S.; Disayathanoowat, T. Soil bacterial communities and their associated functions for forest restoration on a limestone mine in northern Thailand. PLoS ONE 2021, 16, e0248806. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef] [PubMed]
- Abakumov, E.; Zverev, A.; Kichko, A.; Kimeklis, A.; Andronov, E. Soil microbiome of different-aged stages of self-restoration of ecosystems on the mining heaps of limestone quarry (Elizavetino, Leningrad Region). Open Agric. 2021, 6, 57–66. [Google Scholar] [CrossRef]
- de Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Jørgensen, H.B.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbial. 2018, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Hoffland, E.; Kuyper, T.W.; Wallander, H.; Plassard, C.; Gorbushina, A.A.; Haselwandter, K.; Holmström, S.; Landeweert, R.; Lundström, U.S.; Rosling, A.; et al. The role of fungi in weathering. Front. Ecol. Environ. 2004, 2, 258–264. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F. Application and Mechanisms of Plant Growth Promoting Fungi (PGPF) for Phytostimulation; IntechOpen: London, UK, 2020; ISBN 978-1-78984-669-0. [Google Scholar]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant growth-promoting fungi (PGPF) instigate plant growth and induce disease resistance in Capsicum annuum L. upon infection with Colletotrichum capsici (Syd.) Butler & Bisby. Biomolecules 2020, 10, 41. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Grez, B.; Muñoz-Rojas, M.; Kariman, K.; Storer, P.; O’Donnell, A.G.; Kumaresan, D.; Whiteley, A.S. Reconditioning degraded mine site soils with exogenous soil microbes: Plant fitness and soil microbiome outcomes. Front. Microbiol. 2019, 10, 1617. [Google Scholar] [CrossRef] [Green Version]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Sansupa, C.; Fareed Mohamed Wahdan, S.; Disayathanoowat, T.; Purahong, W. Identifying hidden viable bacterial taxa in tropical forest soils using amplicon sequencing of enrichment cultures. Biology 2021, 10, 569. [Google Scholar] [CrossRef]
- Maxwell, J.F.; Elliott, S. Vegetation and Vascular Flora of Doi Sutep-Pui National Park, Northern Thailand; Biodiversity Research and Training Program: Bangkok, Thailand, 2001. [Google Scholar]
- Elliott, S.; David, B.; Hardwick, K. Restoring Tropical Forests: A Practical Guide; Royal Botanic Gardens, Kew: Richmond, UK, 2014. [Google Scholar]
- Shade, A.; Hogan, C.S.; Klimowicz, A.K.; Linske, M.; McManus, P.S.; Handelsman, J. Culturing captures members of the soil rare biosphere. Environ. Microbiol. 2012, 14, 2247–2252. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38-Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Schöps, R.; Goldmann, K.; Herz, K.; Lentendu, G.; Schöning, I.; Bruelheide, H.; Wubet, T.; Buscot, F. Land-use intensity rather than plant functional identity shapes bacterial and fungal rhizosphere communities. Front. Microbiol. 2018, 9, 02711. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for Illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Wang, J.Z.; Du, Z.; Payattakool, R.; Yu, P.S.; Chen, C.-F. A New method to measure the semantic similarity of GO terms. Bioinformatics 2007, 23, 1274–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, F.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-4. 2017. Available online: https://cran.r-project.org/package=vegan (accessed on 5 March 2021).
- Põlme, S.; Abarenkov, K.; Henrik Nilsson, R.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Varma, A. Role of Enzymes in Maintaining Soil Health. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 25–42. ISBN 978-3-642-14225-3. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistical software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- R Development Core Team. A language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria, 2019; pp. 1013–1453. [Google Scholar]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJ. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Newman, M.E.J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef]
- Watson, C. BrainGraph: Graph Theory Analysis of Brain MRI Data. R Package; Version 3.0.2; 2020; Available online: https://CRAN.R-project.org/package=brainGraph (accessed on 8 March 2021).
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Gaber, D.A.; Berthelot, C.; Camehl, I.; Kovács, G.M.; Blaudez, D.; Franken, P. Salt stress tolerance of dark septate endophytes is independent of melanin accumulation. Front. Microbiol. 2020, 11, 3158. [Google Scholar] [CrossRef]
- Li, X.; Han, S.; Wang, G.; Liu, X.; Amombo, E.; Fu, J. The fungus Aspergillus aculeatus enhances salt-stress tolerance, metabolite accumulation, and improves forage quality in perennial ryegrass. Front. Microbiol. 2017, 8, 1664. [Google Scholar] [CrossRef]
- Perl, T.; Kis-Papo, T.; Nevo, E. Fungal biodiversity in the hypersaline dead sea: Extinction and evolution. Biol. J. Linn. Soc. 2017, 121, 122–132. [Google Scholar] [CrossRef]
- Joshi, S.; Bajpai, A.; Johri, B.N. Chapter 1—Extremophilic Fungi at the Interface of Climate Change. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology; Sharma, V.K., Shah, M.P., Parmar, S., Kumar, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 1–22. ISBN 978-0-12-821925-6. [Google Scholar]
- Ismail; Hamayun, M.; Hussain, A.; Afzal Khan, S.; Iqbal, A.; Lee, I.-J. Aspergillus flavus promoted the growth of soybean and sunflower seedlings at elevated temperature. Biomed Res. Int. 2019, 2019, 1295457. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, A.; Thanmalagan, R.R.; Roy, A.; Arunachalam, A.; Lakshmi, P. Strategies to understand Aspergillus flavus resistance mechanism in Arachis hypogaea L. Curr. Plant Biol. 2019, 20, 100123. [Google Scholar] [CrossRef]
- Bultman, K.M.; Kowalski, C.H.; Cramer, R.A. Aspergillus fumigatus virulence through the lens of transcription factors. Med. Mycol. 2017, 55, 24–38. [Google Scholar] [CrossRef]
- Qin, W.; Liu, C.; Jiang, W.; Xue, Y.; Wang, G.; Liu, S. A coumarin analogue nfa from endophytic Aspergillus fumigatus improves drought resistance in rice as an antioxidant. BMC Microbiol. 2019, 19, 50. [Google Scholar] [CrossRef] [PubMed]
- Abrashev, R.; Stoitsova, S.; Krumova, E.; Pashova, S.; Paunova-Krasteva, T.; Vassilev, S.; Dolashka-Angelova, P.; Angelova, M. Temperature-stress tolerance of the fungal strain Aspergillus niger 26: Physiological and ultrastructural changes. World J. Microbiol. Biotechnol. 2014, 30, 1661–1668. [Google Scholar] [CrossRef]
- Ismail; Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.-J. Aspergillus niger boosted heat stress tolerance in sunflower and soybean via regulating their metabolic and antioxidant system. J. Plant Interact. 2020, 15, 223–232. [Google Scholar] [CrossRef]
- Yoo, S.-J.; Shin, D.J.; Won, H.Y.; Song, J.; Sang, M.K. Aspergillus terreus JF27 promotes the growth of tomato plants and induces resistance against Pseudomonas Syringae Pv. tomato. Mycobiology 2018, 46, 147–153. [Google Scholar] [CrossRef]
- Pang, K.-L.; Chiang, M.W.-L.; Guo, S.-Y.; Shih, C.-Y.; Dahms, H.U.; Hwang, J.-S.; Cha, H.-J. Growth study under combined effects of temperature, ph and salinity and transcriptome analysis revealed adaptations of Aspergillus terreus NTOU4989 to the extreme conditions at Kueishan Island Hydrothermal Vent Field, Taiwan. PLoS ONE 2020, 15, e0233621. [Google Scholar] [CrossRef]
- Morsy, M.; Cleckler, B.; Armuelles-Millican, H. Fungal endophytes promote tomato growth and enhance drought and salt tolerance. Plants 2020, 9, 877. [Google Scholar] [CrossRef]
- Kanse, O.S.; Whitelaw-Weckert, M.; Kadam, T.A.; Bhosale, H.J. Phosphate solubilization by stress-tolerant soil fungus Talaromyces funiculosus SLS8 isolated from the neem rhizosphere. Ann. Microbiol. 2015, 65, 85–93. [Google Scholar] [CrossRef]
- Zhou, W.-N.; White, J.F.; Soares, M.A.; Torres, M.S.; Zhou, Z.-P.; Li, H.-Y. Diversity of fungi associated with plants growing in geothermal ecosystems and evaluation of their capacities to enhance thermotolerance of host plants. J. Plant Interact. 2015, 10, 305–314. [Google Scholar] [CrossRef]
- Kavroulakis, N.; Doupis, G.; Papadakis, I.E.; Ehaliotis, C.; Papadopoulou, K.K. Tolerance of tomato plants to water stress is improved by the root endophyte Fusarium solani FsK. Rhizosphere 2018, 6, 77–85. [Google Scholar] [CrossRef]
- Ko, W.-H.; Yang, C.-H.; Mei-Ju, L.; Chen, C.-Y.; Tsou, J.Y. Humicola phialophoroides sp. nov. from soil with potential for biological control of plant diseases. Bot. Stud. 2011, 52, 197–202. [Google Scholar]
- Yang, C.-H.; Lin, M.-J.; Su, H.-J.; Ko, W.-H. Multiple resistance-activating substances produced by Humicola phialophoroides isolated from soil for control of Phytophthora blight of pepper. Bot. Stud. 2014, 55, 40. [Google Scholar] [CrossRef]
- Yokoya, K.; Postel, S.; Fang, R.; Sarasan, V. Endophytic fungal diversity of Fragaria vesca, a crop wild relative of strawberry, along environmental gradients within a small geographical area. PeerJ 2017, 5, 2860. [Google Scholar] [CrossRef]
- Bashyal, B.M.; Parmar, P.; Zaidi, N.W.; Aggarwal, R. Molecular programming of drought-challenged Trichoderma harzianum-bioprimed rice (Oryza sativa L.). Front. Microbiol. 2021, 12, 573. [Google Scholar] [CrossRef]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Ngugi, M.R.; Fechner, N.; Neldner, V.J.; Dennis, P.G. Successional dynamics of soil fungal diversity along a restoration chronosequence post-coal mining. Restor. Ecol. 2020, 28, 543–552. [Google Scholar] [CrossRef]
- Kane, J.L.; Morrissey, E.M.; Skousen, J.G.; Freedman, Z.B. Soil microbial succession following surface mining is governed primarily by deterministic factors. FEMS Microbiol. Ecol. 2020, 96, fiaa114. [Google Scholar] [CrossRef]
- Hart, M.; Gorzelak, M.A.; McAmmond, B.M.; Van Hamme, J.D.; Stevens, J.; Abbott, L.; Whiteley, A.S.; Nevill, P. Fungal communities resist recovery in sand mine restoration. Front. For. Glob. Chang. 2019, 2, 78. [Google Scholar] [CrossRef]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Diversity, taxonomy and phylogeny of the fungi. Biol. Rev. 2019, 94, 2101–2137. [Google Scholar] [CrossRef] [PubMed]
- Djemiel, C.; Dequiedt, S.; Karimi, B.; Cottin, A.; Horrigue, W.; Bailly, A.; Boutaleb, A.; Sadet-Bourgeteau, S.; Maron, P.-A.; Chemidlin Prévost-Bouré, N.; et al. Potential of meta-omics to provide modern microbial indicators for monitoring soil quality and securing food production. Front. Microbiol. 2022, 13, 889788. [Google Scholar] [CrossRef]
- Schoch, C.L.; Sung, G.-H.; López-Giráldez, F.; Townsend, J.P.; Miadlikowska, J.; Hofstetter, V.; Robbertse, B.; Matheny, P.B.; Kauff, F.; Wang, Z.; et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Biol. 2009, 58, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.T.; Wösten, H.A.B.; Dijksterhuis, J. Chapter Two–Fungal Spores for Dispersion in Space and Time. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 85, pp. 43–91. [Google Scholar]
- Hagiwara, D.; Sakai, K.; Suzuki, S.; Umemura, M.; Nogawa, T.; Kato, N.; Osada, H.; Watanabe, A.; Kawamoto, S.; Gonoi, T.; et al. Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 2017, 12, e0177050. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; de Vries, R.P. Compatible solutes and fungal development. Biochem. J. 2006, 399, 3. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, J.; Burns, B.P.; Charlesworth, J.; Burns, B.P. Extremophilic adaptations and biotechnological applications in diverse environments. AIMS Microbiol. 2016, 2, 251–261. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, R.; Sun, T.; Liang, Y.; Jiang, Y.; Sun, B. Organic amendments shift the phosphorus-correlated microbial co-occurrence pattern in the peanut rhizosphere network during long-term fertilization regimes. Appl. Soil Ecol. 2017, 124, 229–239. [Google Scholar] [CrossRef]
- de Menezes, A.B.; Prendergast-Miller, M.T.; Richardson, A.E.; Toscas, P.; Farrell, M.; Macdonald, L.M.; Baker, G.; Wark, T.; Thrall, P.H. Network analysis reveals that bacteria and fungi form modules that correlate independently with soil parameters. Environ. Microbiol. 2015, 17, 2677–2689. [Google Scholar] [CrossRef]
- Jiang, Y.; Sun, B.; Li, H.; Liu, M.; Chen, L.; Zhou, S. Aggregate-related s in network patterns of nematodes and ammonia oxidizers in an acidic soil. Soil Biol. Biochem. 2015, 88, 101–109. [Google Scholar] [CrossRef]
- Ma, B.; Wang, Y.; Ye, S.; Liu, S.; Stirling, E.; Gilbert, J.A.; Faust, K.; Knight, R.; Jansson, J.K.; Cardona, C.; et al. Earth microbial co-occurrence network reveals interconnection pattern across microbiomes. Microbiome 2020, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Claassens, S.; Van Rensburg, P.J.J.; Van Rensburg, L. Soil microbial community structure of coal mine discard under rehabilitation. Water Air Soil Pollut. 2006, 174, 355–366. [Google Scholar] [CrossRef]
- Gastauer, M.; Vera, M.P.O.; de Souza, K.P.; Pires, E.S.; Alves, R.; Caldeira, C.F.; Ramos, S.J.; Oliveira, G. A Metagenomic survey of soil microbial communities along a rehabilitation chronosequence after iron ore mining. Sci. Data 2019, 6, 190008. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.; Yan, J. Biodiversity of fungi on Vitis Vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Purahong, W.; Mapook, A.; Wu, Y.-T.; Chen, C.-T. Characterization of the Castanopsis Carlesii Deadwood Mycobiome by Pacbio Sequencing of the Full-Length Fungal Nuclear Ribosomal Internal Transcribed Spacer (ITS). Front. Microbial. 2019, 10, 983. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.T.V.; Irinyi, L.; Meyer, W. Long-read sequencing in fungal identification. Microbiol. Aust. 2022, 43, 14–18. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis Vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Genus | Detected Species | Stress Factors | Possible Adaptive Strategies to Extreme Environments | Application on Plant | References |
|---|---|---|---|---|---|---|
| Dothideomycetes | Periconia | unidentified | Salinity | -* | - | [41] |
| Others | Angustimassarina acerina, Curvularia clavata, Paraconiothyrium brasiliense and Valsaria neotropica | - | - | - | - | |
| Eurotiomycetes | Aspergillus | Aspergillus aculeatus | Drought | Conidia | Promote plant growth in Drought and salt stress | [42] |
| Aspergillus amstelodami | Salinity | - | - | [43,44] | ||
| Aspergillus flavus | Drought | Conidia or sclerotia | Promote plant growth under heat stress | [45,46] | ||
| Aspergillus fumigatus | Drought and oxidative stress | - | Improves drought resistance | [47,48] | ||
| Aspergillus niger | High temperature | Conidia | Heat-stress ameliorative tool | [49,50] | ||
| Aspergillus terreus | High temperature, pH and salinity | Stress response gene | Promote plant growth and control disease | [51,52] | ||
| Penicillium | Unidentified | Drought and salinity | - | Enhance drought and salt tolerance | [53] | |
| Talaromyces | Unidentified | Sanity and oxidative stress | - | - | [54] | |
| Pezizomycotina Incertae sedis | Acrophialophora | Acrophialophora fusispora | High temperature | - | - | [55] |
| Sordariomycetes | Fusarium | Fusarium solani | Drought | - | Promote plant growth under drought | [56] |
| Gibberella | Gibberella intricans | - | - | - | - | |
| Humicola | Humicola phialophoroides | Salinity | - | Control of plant diseases/enhance growth in salt stress | [57,58,59] | |
| Trichoderma | unidentified | Drought | - | Enhance drought tolerance | [60,61] | |
| Others | Hypoxylon anthochroum, Hypoxylon monticulosum, Nigrospora oryzae and Xylaria apiculata | - | - | - | - | |
| Ustilaginomycetes/ Agaricomycetes/ Cystobasidiomycetes | Pseudozyma | Pseudozyma hubeiensis | - | - | - | - |
| Coprinellus | Coprinellus aureogranulatus | - | - | - | - | |
| Gymnopilus | Gymnopilus dilepis | - | - | - | - | |
| Phellinus | Phellinus noxius | - | - | - | - | |
| Cystobasidium | Unidentified | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sansupa, C.; Purahong, W.; Nawaz, A.; Wubet, T.; Suwannarach, N.; Chantawannakul, P.; Chairuangsri, S.; Disayathanoowat, T. Living Fungi in an Opencast Limestone Mine: Who Are They and What Can They Do? J. Fungi 2022, 8, 987. https://doi.org/10.3390/jof8100987
Sansupa C, Purahong W, Nawaz A, Wubet T, Suwannarach N, Chantawannakul P, Chairuangsri S, Disayathanoowat T. Living Fungi in an Opencast Limestone Mine: Who Are They and What Can They Do? Journal of Fungi. 2022; 8(10):987. https://doi.org/10.3390/jof8100987
Chicago/Turabian StyleSansupa, Chakriya, Witoon Purahong, Ali Nawaz, Tesfaye Wubet, Nakarin Suwannarach, Panuwan Chantawannakul, Sutthathorn Chairuangsri, and Terd Disayathanoowat. 2022. "Living Fungi in an Opencast Limestone Mine: Who Are They and What Can They Do?" Journal of Fungi 8, no. 10: 987. https://doi.org/10.3390/jof8100987